茶树芽叶紫化的生理生化分析及其关键酶基因的表达
2015-10-25周琼琼孙威江
周琼琼孙威江,2
(1. 福建农林大学园艺学院,福州 350002;2. 福建农林大学安溪茶学院,福州 350002)
茶树芽叶紫化的生理生化分析及其关键酶基因的表达
周琼琼1孙威江1,2
(1. 福建农林大学园艺学院,福州 350002;2. 福建农林大学安溪茶学院,福州 350002)
从生理生化和分子水平方面比较了茶树紫化芽叶与成熟绿色叶片的差异。结果表明,紫色幼嫩新梢中茶多酚、儿茶素总量、咖啡碱含量高于成熟绿色叶片,差异达到极显著。光合色素中,成熟绿叶样品中叶绿素及叶绿素a、b的含量、类胡萝卜素的含量极显著高于幼嫩紫叶样品,花青素含量极显著低于幼嫩紫叶;在研究的9个花青素合成途径关键酶基因中,实时荧光定量PCR分析表明,PAL、C4H、CHS、CHI、F3H、F3'H、F3'5'H、DFR和ANS在幼嫩紫叶中均呈上调表达,从而促进花青素的合成,使芽叶呈现紫色。
茶树;紫色叶片;理化分析;基因表达
DIO: 10.13560/j.cnki.biotech.bull.1985.2015.01.013
植物色素包括类胡萝卜素、叶绿素、花青素等,是影响植物色泽的重要生化成分,在植物的次生代谢和视觉吸引力方面发挥着重要作用[1]。花青素是一类广泛存在于植物中的水溶性色素,属类黄酮化合物,在植物中常以糖苷形式存在于植物细胞的液泡中。因植物细胞液泡不同的pH值条件,使花瓣、叶片或果实呈现出不同的颜色,pH<7呈红色,pH=7-8时呈紫色,pH>10时呈蓝色[2]。对于植物而言,花青素具有多种生理功能,如吸引昆虫传粉、避免紫外线伤害、抵御低温和防治植物病害等方面起着重要作用;对于人类而言,花青素作为一种天然色素,具有安全、无毒、资源丰富的特性,已广泛应用于食品、化妆品领域。
茶树作为一种多年生常绿叶用经济作物,芽叶的色泽与鲜叶的内含成分、鲜叶的适制性和成茶品质都有着密切的关系。茶树鲜叶的细胞液呈弱酸性,故当芽叶花青素含量较高时会呈紫红色[3]。紫色芽叶茶树是一种特色的茶树种质资源,在地方有性群体品种中占有很大比例。武夷山是福建省茶树种质资源的宝库,具有成百上千份珍贵优异的茶树种质资源,其中武夷奇种C18-1是典型的代表,幼嫩时新梢为紫色,但随着生长发育过程,茶树叶片颜色从幼嫩的紫色转为成熟时的绿色。茶树幼嫩芽叶的紫化现象以及特异性的紫色茶树品种被认为与花青素在叶片中的累积有关[4]。紫芽茶中花青素含量较正常芽叶高很多[5],其形成和积累与茶树生长发育状态和环境条件密切相关,较强的光照和高温能增加其含量,使茶树芽叶呈紫红色,同时也是由植物体内相关基因调控所决定的。在花青素合成的途径中,直接编码花青素苷生物合成途径中的关键酶类有查尔酮合成酶(Chalcone synthase,CHS)、查尔酮异构酶(Chalcone isomerase,CHI)、黄烷酮3-羟化酶(Flavanone 3-hydroxylase,F3H)、二羟基黄酮醇还原酶(Dihydroflavonol 4-reductase,DFR)、花色素苷合成酶(Anthocyanin,ANS)以及类黄酮3-O-糖基转移酶(Flavonoid 3-O-glucosyltransferase,UFGT)均属于花青素生物合成酶。
本研究拟通过研究幼嫩叶片与成熟叶片之间的理化成分差异以及花青素合成过程中关键酶的基因表达情况,旨在为深入研究茶树叶片颜色转变提供理论依据。
1 材料与方法
1.1 材料
1.1.1 材料 样品于2013年春采于福建省武夷山市茶叶研究所资源圃,分别采摘武夷奇种C18-1同一时期不同成熟度的茶树叶片,即幼嫩紫色叶片与成熟绿色叶片,按照五点采样法混合均匀后,迅速用液氮处理,置-80℃冰箱保存备用。
1.1.2 试剂与设备 表没食子儿茶素没食子酸酯(EGCG,纯度>99%)、表没食子酸儿茶素(EGC,纯度>99%)、表儿茶素(EC,纯度>99%)、儿茶素(C,纯度>99%)、表儿茶素没食子酸酯(ECG,纯度>99%)、咖啡碱(Caffeine,纯度>99%)、没食子酸(GA)标样购自美国Sigma公司;Waters2890高效液相色谱仪(美国Waters公司)、台式高速冷冻离心机(AllegraTM21RC Centrifuge Beckman)、电泳仪(北京六一仪器厂DYY-10C型)、电泳槽(北京六一仪器厂DYC-20C)、凝胶成像系统(美国UVP Gel Doc-It)、PCR仪(德国Biometra公司)。
1.2 方法
1.2.1 茶多酚含量测定 采用福林酚(Folin-Ciocalten)比色法。参照GB/T 8313-2008。
1.2.2 儿茶素总量及组分、咖啡碱的测定 参照周琼琼等[6]测定样品儿茶素组分及其总量以及咖啡碱的含量。
1.2.3 光合色素含量的测定 叶片分别用80%(V/ V)丙酮提取色素后,按Lichtenthaler[7]的方法测定叶片叶绿素(Chl)、叶绿素a(Chla)、叶绿素b(C-hlb)与类胡萝卜素(Car)的含量。
1.2.4 花青素含量的测定 参照Pang[8]的方法提取花青素测定液。用紫外可见分光光度计测量530 nm和657 nm的吸光度(OD),根据公式计算总花青素苷含量(OD/g·FW),OD=OD530-0.25×OD657。
1.2.5 茶树叶片总RNA的提取和cDNA的合成 本试验参照多糖多酚植物总RNA提取试剂盒(天根生化科技(北京)有限公司)提取茶树总RNA。cDNA第一链的合成参照RevertAidTMFirst-Strand cDNA Synthesis Kit(Thermoscientific,Rockford,IL 61101,United States)试剂盒说明书进行。
1.2.6 花青素苷合成关键基因的表达分析 根据GenBank已公布的茶树花青苷合成关键酶基因的序列或同源基因的氨基酸保守序列和核苷酸序列,用Primer Premier 5.0设计相应的荧光定量PCR引物,以茶树β-tubulin为内参基因(表1),按照SYBR®Premix Ex TaqTM(TaKaRa,Japan)试剂盒说明进行荧光定量PCR。采用2-ΔΔCt方法分析各关键酶基因的相对表达量及其标准差。每个样品检测设置3个重复,整个试验重复3次。
2 结果
2.1 茶多酚含量分析
茶鲜叶中多酚类物质与茶树的生长发育、新陈代谢和茶叶品质关系密切。多酚类物质主要集中于茶树新梢的生长旺盛部分,不同伸育程度的茶叶中茶多酚含量存在明显差异;不同色泽的茶鲜叶其化学成分的含量也是有差异的。图1显示,紫色幼嫩新梢中茶多酚含量高于成熟绿色叶片,差异达到极显著。

表1 花青素合成关键酶基因及内参基因的引物序列
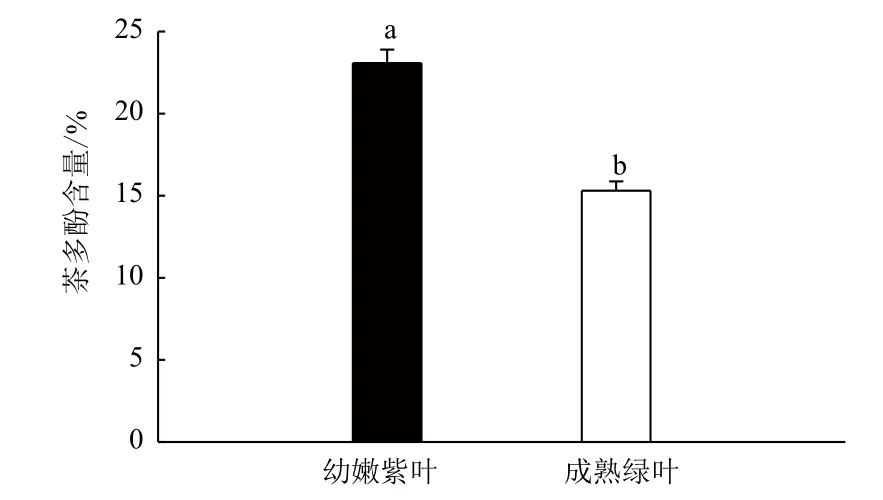
图1 不同色泽的叶片中茶多酚含量
2.2 儿茶素总量及组分、咖啡碱含量分析
研究结果(表2)表明,两个样品的儿茶素组成中酯型儿茶素的含量均高于简单儿茶素含量,含量最高的是EGCG,其次是EGC和ECG,C含量最低。儿茶素总量及组分在紫色幼嫩新梢和绿色成熟叶片中差异均达到了极显著水平。咖啡碱在两个样品中差异也达到了极显著水平。

表2 不同颜色的叶片中儿茶素总量及组分、咖啡碱含量
2.3 光合色素含量的分析
叶片是光合作用的主要器官,叶绿体是进行光合作用的主要细胞器,叶绿素是参与光合作用光能吸收、传递和转化的重要色素。图2显示,成熟绿叶样品中叶绿素及叶绿素a、b的含量(图2-D、2-A、2-B)、类胡萝卜素的含量(图2-C)均高于幼嫩紫叶样品,且差异达到了极显著水平;而成熟绿叶样品的叶绿素a/b(图2-E)、类胡萝卜素/叶绿素的比值(图2-F)均低于幼嫩紫叶样品,差异也达到了极显著水平。
2.4 花青素含量的分析
图3表明,不同色泽的茶叶样品中花青素含量差异明显。紫色幼嫩新梢中花青素含量高于成熟绿色叶片,差异达到极显著。茶树芽叶呈紫色是由其高含量的花青素组分引起的。
2.5 花青素合成关键酶基因的荧光定量分析
选取茶叶中已获得的与花青素类物质代谢相关的9个结构因子PAL、C4H、CHS、CHI、F3H、F3'H、F3'5'H、DFR和ANS,对其在不同部位茶叶中的表达情况进行实时荧光定量分析,各目标基因在不同叶片中的相对表达量采用比较阈值法,即通过计算2-ΔΔCT来确定。结果(图4)表明,在鲜叶不同发育时期,PAL、C4H、CHS、CHI、F3H、F3'H、F3'5'H、DFR和ANS基因的表达水平与儿茶素积累规律一致。在研究的9个花青素合成途径关键酶基因中,上游基因PAL、C4H、CHI、F3'H、F3'5'H在幼嫩紫叶和成熟绿叶中表达水平呈极显著差异,即在幼嫩紫叶中均呈上调表达,说明花青素生物合成的上游关键酶基因对幼嫩叶片中花青素合成过程中有重要作用。CHS、F3H、DFR基因的表达水平在幼嫩紫叶和成熟绿叶中表达水平差异不显著,其原因有待进一步验证。

图2 不同色泽的叶片中光和色素的含量
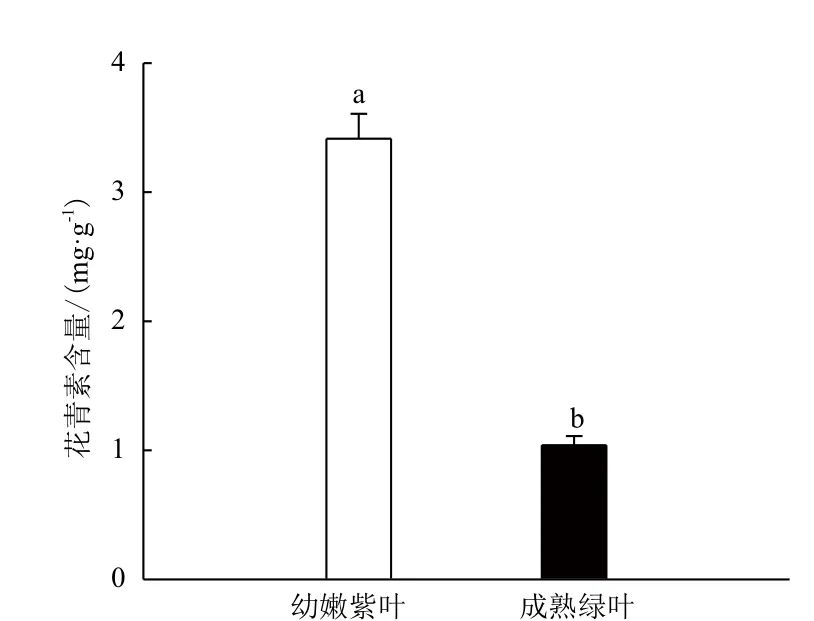
图3 不同色泽的叶片中花青素含量
3 讨论
3.1 生化成分变化
茶树新梢所含的生化成分是形成成茶品质的重要物质基础,多酚类物质不仅是形成茶叶色香味的主要成分之一,也是茶叶发挥保健作用的重要功能性成分。本研究的结果是紫色幼嫩新梢中茶多酚含量高于成熟绿色叶片,这与刘富知[9]、周志高等[10]的研究结果一致,红紫色芽叶较绿色芽叶含有较多的茶多酚。萧力争等[11]的研究结果也表明,同一品种,紫色芽叶中的儿茶素含量比正常芽叶高(除简单儿茶素外)。茶树芽叶呈紫色与芽叶中的花青素含量关系密切,研究表明红紫芽茶树品系芽叶的红紫色是由其高含量的花青素组分引起的,且成正相关,随着花青素含量的增加其芽叶的红紫色加深[12]。
3.2 色素含量变化
花青素含量与茶树生长发育阶段及环境条件关系密切,光照强、气温高则有利于花青素的累积,红紫色芽叶的形成便是花青素积累较多的标志。叶片中花青素的合成与其它光合色素有显著的联系。随着叶片的成熟,叶片中的各种色素呈现出不同的消长变化。这与蔡丽等[13]的研究结果是一致的。叶绿素在弱光下合成的多,在强光下一部分叶绿素被破坏而由胡萝卜素取而代之,因此这些叶片在遮荫、光照不足的情况下是绿色的,在强光下变成紫色。幼嫩新梢中含有较高的花青素,可能是通过花青素吸收光子从而达到保护叶绿体的作用。有研究表明[14,15],含有花青素成分的紫叶通常比绿叶吸收的绿光多,花青素通过拦截能量量子,保护尚不成熟的叶绿体使其发育成熟,幼嫩叶片中花青素的积累和植物耐光抑制呈正相关,从而使叶绿体发育成熟避免过量的光辐射造成干扰,也是植物本身对逆境环境的一种适应机制。

图4 不同色泽的叶片中花青素合成途径关键酶基因的实时定量表达分析
3.3 基因表达量变化
花青素代谢合成途径涉及诸多因素,包括环境因素、信号转导、关键酶基因的表达及转录因子的调控等。这些环境因子的作用及色素的合成都是通过基因表达来起作用的[16]。目前,已有一些结构基因和调节基因从多种植物的花、果皮等组织中被分离和克隆出来,其结构基因的表达和调控也有较深入的研究,主要集中于模式植物,如拟南芥[17]、矮牵牛花[18]、金鱼草[19]等,对茶叶的研究较少。本研究对不同部位茶叶花青素合成相关结构基因的表达量进行了分析,结果表明同一株茶树不同部位茶叶色泽的差异可能是由于基因表达丰度的强弱从而影响花青素含量,成熟绿叶中相关基因表达量虽减少,但仍有一定的表达量,原因可能是由于上游基因除了合成与花青素相关的酶以外,还参与了类黄酮生化途径其它分支上有关物质,如黄酮类物质、儿茶素的合成等。夏涛[20]课题组利用Q-PCR分析,发现具有不同儿茶素含量的细胞系之间F3H、ANS、ANR、DFR基因表达水平差异较大,DFR是类黄酮途径中花青素合成的关键酶,该酶可催化二氢黄酮醇转变为相应的黄烷-3,4-二醇,即变为无色花色素,而后者又是花色素苷、儿茶素和原花色素形成的共同前体。在茶树芽叶发育期间,DFR的表达与儿茶素和多酚类的积累密切相关,而成熟绿叶中茶多酚、儿茶素总量低于幼嫩紫叶,其相关基因是如何调控则需进一步分析。
4 结论
本研究从生理生化和分子水平方面比较了茶树同一时期不同发育程度紫化芽叶与成熟绿色叶片的差异,结果表明紫色幼嫩新梢中茶多酚、儿茶素总量、咖啡碱含量高于成熟绿色叶片,差异达到极显著;光合色素中,成熟绿叶样品中叶绿素及叶绿素a、b的含量、类胡萝卜素的含量极显著高于幼嫩紫叶样品,花青素含量极显著低于幼嫩紫叶;在研究的9个花青素合成途径关键酶基因中,实时荧光定量PCR分析表明,PAL、C4H、CHS、CHI、F3H、F3'H、F3'5'H、DFR和ANS在幼嫩紫叶中均呈上调表达,从而促进花青素的合成,使芽叶呈现紫色。
[1]Bamberg J, Bethke P, Brunet J, et al. Plant pigments for color and nutrition[J]. HortScience, 1997, 32(1):12-13.
[2]张宏宝. 茶树红紫色芽叶中花青素组分的分离及鉴定[D]. 济南:山东农业大学, 2009.
[3]杨贤强, 王岳飞, 陈留记. 茶多酚化学[M]. 上海:上海科学技术出版社, 2003.
[4]马春雷, 姚明哲, 王新超, 等. 利用基因芯片筛选茶树芽叶紫化相关基因[J]. 茶叶科学, 2011, 31(1):59-65.
[5]萧力争, 苏晓倩, 李勤, 等. 紫芽品种茶树春梢芽叶生化成分分析[J]. 福建农林大学学报:自然科学版, 2009, 38(1):30-33.
[6] 周琼琼, 孙威江, 叶艳, 等. 不同年份白茶的主要生化成分分析[J]. 食品工业科技, 2014, 35(9):351-354.
[7] Lichtenthaler HK. Chlorophylls and carotenoids:pigments of photosynthetic biomembranes[J]. Methods Enzymol, 1987, 148:350-382.
[8] Pang Y, Peel GJ, Sharma SB, et al. A transcript profiling approach reveals an epicatechin-specific glucosyltransferase expressed in the seed coat of Medicago truncatula[J]. Proceedings of the National Academy of Sciences, 2008, 105(37):14210-14215.
[9] 刘富知, 黄建安, 付冬和, 等. 茶树上红紫色芽叶部分生化特性的研究[J]. 湖南农业大学学报, 2000, 26(1):41-42.
[10]周志高, 车玉萍, 罗质超, 等. 历史名茶——阳羡紫笋的品质化学初探[J]. 蚕桑茶叶通讯, 2004(1):23-25.
[11]萧力争, 苏晓倩, 李勤, 等. 紫芽品种茶树芽叶多酚类物质组成特征[J]. 湖南农业大学学报:自然科学版, 2008, 34(1):77-79.
[12]吴华铃, 何玉媚, 李家贤, 等. 11个红紫芽茶树新品系的芽叶特性和生化成分研究[J]. 植物遗传资源学报, 2012, 13(1):42-47.
[13]蔡丽, 梁名志, 夏丽飞, 等. “紫娟” 茶外观表象差异研究[J].西南农业学报, 2010, 23(3):700-703.
[14]Manetas Y, Drinia A, Petropoulou Y. High contents of anthocyanins in young leaves are correlated with low pools of xanthophyll cycle components and low risk of photoinhibition[J]. Photosynthetica,2002, 40(3):349-354.
[15] Zhang K, Wang X, Cui J, et al. Characteristics of gas exchange and chlorophyll fluorescence in red and green leaves of Begonia semperflorens[J]. Biologia Plantarum, 2011, 55(2):361-364.
[16]史宝胜. 紫叶李叶色生理变化及影响因素研究[D]. 哈尔滨:东北林业大学, 2006.
[17] Bartel B, Matsuda SP. Seeing red[J]. Science, 2003, 299(5605):352-353.
[18] Quattrocchio F, Wing JF, Leppen HT, et al. Regulatory genes controlling anthocyanin pigmentation are functionally conserved among plant species and have distinct sets of target genes[J]. The Plant Cell Online, 1993, 5(11):1497-1512.
[19] Holton TA, Cornish EC. Genetics and biochemistry of anthocyanin biosynthesis[J]. The Plant Cell, 1995, 7(7):1071.
[20]夏涛, 高丽萍. 类黄酮及茶儿茶素生物合成途径及其调控研究进展[J]. 中国农业科学, 2009, 42(8):2899-2908.
(责任编辑 马鑫)
Physiological and Biochemical Analysis of Young Shoot Purple-related and Gene Expression of Key Enzymes in Tea Plant(Camellia sinensis)
Zhou Qiongqiong1Sun Weijiang1,2
(1. College of Horticulture,Fujian Agriculture and Forestry University,Fuzhou 350002;2. Anxi Tea College,Fujian Agriculture and Forestry University,Fuzhou 350002)
The young purple shoots of tea plant(Camellia sinensis)were compared to mature green leaves based on the biochemical and molecular level. Results showed that the content of tea polyphenols,catechins,caffeine and anthocyanins in the tender purple leaves were significantly higher than those in mature green leaves,while the content of chlorophyll and chlorophyll a,b,carotenoids were significantly lower than those mature green leaves. Q-PCR analysis indicated that the key genes of PAL、C4H、CHS、CHI、F3H、F3'H、F3'5'H、DFR and ANS in the anthocyanin biosynthesis were up-regulated in the tender purple leaves,which may contribute to the synthesis of anthocyanins and the color of the young shoots.
tea plant;purple leaves;physical and chemical analysis;gene expression
2014-06-15
国家国际科技合作项目(2010DFB33030-4),国家“863”计划项目(2013AA10260605)
周琼琼,女,博士研究生,研究方向:茶树特异性种质资源、茶树遗传育种与分子生物学;E-mail:zqxy1223@qq.com
孙威江,男,博士,教授,研究方向:茶叶质量安全与茶叶品质化学、茶树遗传育种;E-mail:swj8103@126.com
