21份澳洲坚果种质开花结果物候期的变异分析
2015-10-21张汉周王维杨为海曾辉邹明宏陆超忠万继锋
张汉周 王维 杨为海 曾辉 邹明宏 陆超忠 万继锋
摘 要 系统观测21份澳洲坚果种质资源的开花结果物候期,分析其开花结果物候期的变异规律,并根据物候差异划分物候类型。结果表明:不同种质的开花结果物候期存在不同程度的差异,变异系数为2.91%~37.23%,进入花芽萌动期、花序抽生期、开花期、谢花期、果实膨大期、油份积累期及果实成熟期的时间分别相差约45、43、41、38、30、28、30 d;聚类分析将澳洲坚果种质资源分为物候期特点各异的3类;主成分分析第1、2主成分解释的总变异为 91.985%,更为直观展现了澳洲坚果种质开花结果物候期特点,其结果与聚类分析基本一致。
关键词 澳洲坚果;种质资源;开花;结果;物候期;变异
中图分类号 S664.9 文献标识码 A
Abstract The blooming and fruiting phenophases of 21 Macadamia spp. germplasm resources were observed systematically to analyze the phenological variation and to divide the phenological types according to phenological differences. The results showed that the blooming and fruiting phenophases had a large variation among all tested germplasm resources with variation coefficient of 2.91%-37.23%.The time difference of flower bud sprouting stage, inflorescence elongating stage, blooming stage, flower wilting stage, fruit expanding stage, fruit oil accumulation stage and fruit maturity stage was 45, 43, 41, 38, 30, 28 and 30 d, respectively. Three phenological types of Macadamia spp. germplasm resources were classified by cluster analysis. The results of principal component analysis indicated that the first and second principal components could explain 91.985% of total variance, directly demonstrating the characteristics of blooming and fruiting phenophases. The results were basically consistent with that of the cluster analysis.
Key words Macadamia spp.; Germplasm resources; Blossom; Fruit; Phenological phase; Variation
doi 10.3969/j.issn.1000-2561.2015.11.019
澳洲堅果(Macadamia spp.)又名夏威夷果、澳洲胡桃、昆士兰栗等,属山龙眼科(Proteaceae)澳洲坚果属(Macadamia F. Mull)常绿乔木果树。澳洲坚果原产于澳大利亚昆士兰州南部和新南威尔士州北部的沿海亚热带雨林地区[1-2],其人工栽培历史仅150多年,是一种新兴的高档坚果类果树,正在为越来越多的国家和地区所重视。我国于20世纪70年代末开始对澳洲坚果进行引种试种,已从国外引进品种中筛选出适应我国气候和土壤特点的H2、O.C.、HAES 344、HAES 788、HAES 695、HAES 922等优良品种,选育出南亚1号、南亚2号、桂热1号等新品种(系),目前主要栽植区域是云南和广西,广东、海南、四川、福建、贵州等省也均有种植[3-4]。对澳洲坚果的物候期研究是对其种质资源研究与利用的基础性工作。目前,国内外学者陈国云等[5]、郑树芳等[6]、陈显国等[7]、Boyton和Hardner[8]、Sobierajski等[9]、Rojas等[10]已对澳洲坚果开花结果物候期进行了报道,但由于气候、土壤条件差异,国外学者的研究成果不适用于我国,而国内学者仅对其开花结果物候期进行简单的观测,尚未见系统观测的研究报道。因此,有必要对国外引进的适应我国气候和土壤特点的澳洲坚果优良品种和我国自选品种及优株的开花结果物候期进行研究。本研究以21份澳洲坚果主要种质资源(包括国外引进的适应我国气候和土壤特点的优良品种和我国自选品种及优株)为研究对象,对其开花结果物候期进行了系统观测,分析其开花结果物候期的变异规律,并基于此通过聚类分析和主成分分析方法对其开花结果物候期进行划分,以期为今后澳洲坚果的种质资源鉴定评价及利用提供依据。
1 材料与方法
1.1 材料
以中国热带农业科学院南亚热带作物研究所澳洲坚果种质资源圃内的21份种质资源为研究对象,其中A4、H2、O.C.、Own venture、Yonik、HAES333、HAES344、HAES695、HAES783、HAES788、HAES814、HAES922为从澳大利亚、美国夏威夷引进的适应我国气候和土壤特点的优良品种,南亚1号、南亚2号、南亚3号为我国自选品种以及A、B、D、24、114、116为我国自选优株,所有种质均为嫁接成年树,树龄为10~12 a,常规管理。
1.2 观测方法
采用完全随机区组设计,3次重复,每个小区各选择3株生长正常、长势良好、无病虫害的成年植株为观测对象。于2010年定株观测各种质资源的花期和结果期。开花结果物候期观测方法参照《澳洲坚果种质资源描述规范和数据标准》[11]和农业行业标准《澳洲坚果种质资源鉴定技术规范》[12]。
1.3 数据统计
以1月4日的物候期观察数据为基准(0)对其他物候期数据进行换算后,采用聚类分析方法对所得的原始数据矩阵进行聚类分析,为了消除不同量纲对数据分析产生的影响,首先对原始数值矩阵进行标准差标准化(STD)处理,利用标准化数据计算各个分类运算单位(Operational Taxonomic Unit,OTU)之间的平均欧氏距离,然后采用系统聚类中的离差平方和法作出21个OTU分类结果的树系图。所有数据均在DPS v7.05软件上完成。
2 结果与分析
2.1 物候期的变异分析
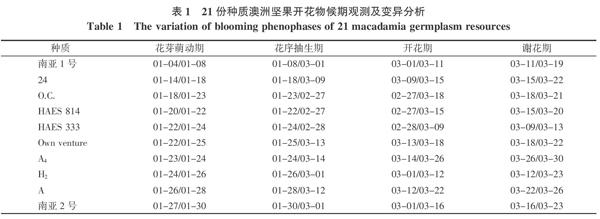
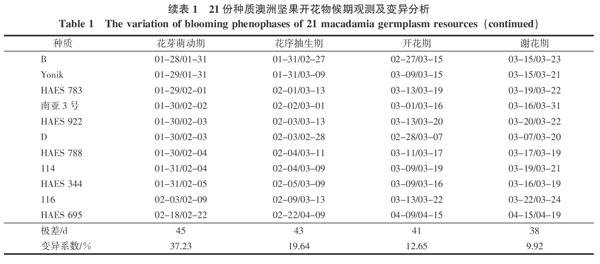
2.1.1 开花物候期的变异分析 由表1可看出,澳洲坚果不同种质间开花物候期存在明显差异。花芽萌动期在1月初至2月初,持续3~7 d;花芽萌动最早的是南亚1号,其次是自选优株24;花芽萌动期最晚的是HAES695,其次是自选优株116;其他种质的花芽均在1月下旬开始萌动;最早和最晚种质相差45 d。花序抽生期多数种质在1月末至3月初开始,到2月末至3月中旬结束,多数持续35~50 d;南亚1号、24、Own venture、A4、B、D和HAES695等7个种质的花序抽生期持续约45 d以上,而其他种质的花序抽生期持续约35~40 d左右;最早和最晚种质相差43 d。除HAES695在4月上中旬完成开花与谢花外,H2、O.C.、HAES333、HAES814、南亚1号、自选优株B、D等种质开花期较早,其他种质的开花期集中在3月上中旬,而谢花期出现在3月中下旬,持续天数在种质之间相差较大;最早和最晚种质分别相差41 d和38 d。
各种质间开花物候期差异,从变异系数来看,花芽萌动期的差异最大,变异系数为37.23%;其次为花序抽生期、开花期和谢花期,变异系数分别为19.64%、12.65%和9.92%。
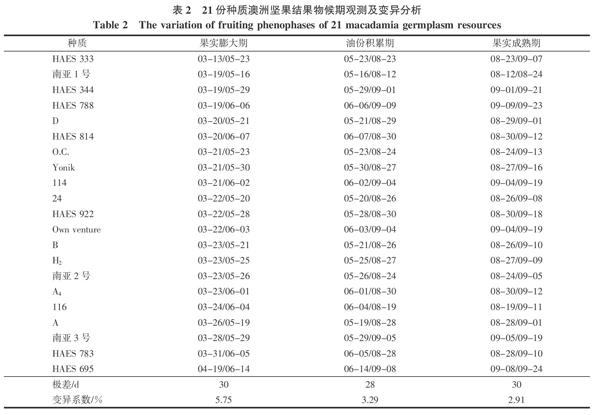
2.1.2 结果物候期的变异分析 如表2所示,澳洲坚果在3月开始陆续有小果出现,HAES333小果出现较早,HAES695小果出现较晚;多数种质的果实膨大期在3月中下旬至6月上中旬,持续天数约50~70 d,其中HAES333、HAES 344、HAES788、南亚1號等种质果实膨大生长较早,HAES695果实膨大生长较晚;最早和最晚种质相差30 d。多数种质果实的油份积累期在5月下旬至8月下旬,持续天数约80~100 d;最早和最晚种质相差28 d;果实最早成熟的种质是南亚1号,果实成熟最晚的是HAES695,多数澳洲坚果种质的果实成熟期在8月下旬至9月中旬;最早和最晚种质相差30 d。从变异系数来看,各种质间结果物候期差异不大,变异系数为2.91%~5.75%。
澳洲坚果的落果期在花期结束即伴随着果实膨大期与油份积累期同时进行,直至果实接近成熟期落果期才基本结束。
2.2 物候期的聚类分析
聚类结果见图1。在欧氏距离12.15处,21份种质资源被明显分为3大类群,即早花早熟型、中花中熟型和晚花晚熟型。早花早熟型(类群Ⅰ)包含 H2、O.C.、HAES333、HAES814、南亚1号、自选优株24、B、D 等8份种质,占总数的38.10%,其花序抽生期、开花期、果实膨大期及果实成熟期较早;晚花晚熟型(类群Ⅲ)只含有HAES695 1份种质,占总数的4.76%,其花芽萌动期、花序抽生期、开花期、谢花期、果实膨大期及果实成熟期较晚;中花中熟型(类群Ⅱ)包含A4、Own venture、Yonik、HAES344、HAES783、HAES788、HAES922、南亚3号、南亚2号、自选优株A、114、116等12份种质,占总数的57.14%,其花序抽生期、开花期、果实膨大期及果实成熟期等开花结果物候期介于类群Ⅰ和类群Ⅲ之间。这些结果为今后对澳洲坚果种质资源的开发利用奠定了实验基础。
2.3 物候期的主成分分析
主成分分析结果(表3)表明,前2个主成分的累积贡献率达到91.985%。第 1 主成分的贡献率为80.493%,主要为花序抽生期、果实膨大期、开花期等物候期,其特征向量绝对值较大,主要反映了花、果实生长物候特点;第2主成分的贡献率为11.492%,果实成熟期等物候期的特征向量绝对值较大,主要反映了果实成熟物候特点。
以主成分分析为基础,以第1主成分值为横坐标,第2主成分值为纵坐标,构建21份澳洲坚果种质资源的二维平面散点图(图2)。Ⅰ类群种质位于原点的左侧,主要体现了第1主成分的花、果实生长物候特点,其花序抽生期、开花期及果实膨大期较早,属于早花早熟型;Ⅱ类群多数种质集中分布在原点四周,种质数量最多,物候表现并不突出,属于中花中熟型;Ⅲ类HAES695单独分布于二维图原点右边最下侧,突出表现为花序抽生期、开花期、果实膨大期及果实成熟期较晚,属于晚花晚熟型。
3 讨论与结论
种质资源是作物遗传育种的物质基础,作物新品种的成功选育大多与优异种质资源的发现和利用有关[13]。因此,开展澳洲坚果种质物候期的研究,发掘其优异资源,对澳洲坚果的育种材料选择有重大意义。澳洲坚果开花结果物候期存在不同程度的差异,进入花芽萌动期、花序抽生期、开花期及谢花期的时间分别相差1个多月,进入果实膨大期、油份积累期及果实成熟期的时间分别相差近1个月,开花、结果物候期的变异系数分别为9.92%~37.23%和2.91%~5.75%。由此可见,澳洲坚果开花结果物候期存在丰富的变异,其开花结果物候期长短的影响因素主要是开花物候期的差异,可为育种材料筛选提供保障。多数供试种质在1月下旬萌动花芽,1月下旬至3月初抽生花序;2月底至3月初陆续有小花开放,盛花期集中在3月上中旬,末花期在3月中下旬,果实成熟于8月末至9月中下旬。这与前人的研究结果不大一致[5-10],可能与品种、气候差异等有关。
澳洲坚果成花量非常大,每个花序可着生100~300朵小花[14],成年树可产生超过100 000朵小花[15]。仅约10%以下的小花才可以在花后2周内成功坐果,但其中又约有80%以上的幼果在花后3~8周内发生脱落[16]。异花授粉可提高澳洲坚果的产量[15]。在澳大利亚,HAES 660与HAES 246授粉,HAES 660的产量比其自花授粉树增产17%[17];HAES 246与O.C.授粉,HAES 246的产量比其自花授粉树增产15%[18]。因此,品种间混种的产量要比单一品种连片种植的产量高。聚类分析将21份澳洲坚果种质在欧氏距离12.15处划分为3大类群,大体可分为早花早熟型、中花中熟型和晚花晚熟型。通过对澳洲坚果开花结果物候期的聚类分析,明确了澳洲坚果种质资源类型,这为品种间的配置提供了依据,但品种间的最佳配置方式还有待进一步研究。通过对澳洲坚果开花结果物候期的观测分析,明确了澳洲坚果开花结果的物候规律,尤其是对开花物候期观察,发现供试的澳洲坚果的花期之间有重叠,这为杂交创造了条件,为杂交亲本选择和种质创新提供了依据。
鉴定评价是种质资源研究工作的重要环节,是种质资源有效利用的前提。利用主成分分析澳洲坚果种质的7个开花结果物候期,筛选出2个主成分,这2个主成分提供了原性状91.985%的信息,以花序抽生期、开花期、果实膨大期、果实成熟期等4个物候期为代表,足以反映开花结果物候期。因此,可将这4个代表物候期作为澳洲坚果種质开花结果物候期鉴定评价指标。
参考文献
[1] Johnson L A S, Briggs B G. On the Proteaceae:the evolution and classification of a southern family[J]. Botanical Journal of the Linnean Society, 1975, 70: 83-182.
[2] Stephenson R. Macadamia: Domestication and commercialization[J]. Chronica Horticultura, 2005, 45(2): 11-15.
[3] 焦 云, 邹明宏, 曾 辉, 等. 澳洲坚果研究进展[J]. 安徽农业科学, 2008, 36(8): 3 366-3 368.
[4] 王文林, 陆超忠, 曾 辉, 等. 我国澳洲坚果的研究及发展[J]. 中国热带农业, 2008(3): 24-25.
[5] 陈国云, 岳 海, 倪书邦, 等. 12个澳洲坚果品种在云南西双版纳的表现[J]. 中国果树, 2009(5): 38-41.
[6] 郑树芳, 覃杰凤, 何铣扬, 等. 10个澳洲坚果品种在广西西南地区的生长结果表现[J]. 中国热带农业, 2011(1): 53-55.
[7] 陈显国, 黄 华, 林玉虹, 等. 澳洲坚果在广西桂中地区的适应性研究[J]. 热带农业科学, 2013, 33(7): 3-34, 53.
[8] Boyton S J, Hardner C M. Phenology of flowering and nut production in macadamia[J]. Revista Acta hortscience(ISHS), 2002, 575(43): 381-387.
[9] Sobierajski G D R, Barbosa W, Neto B, et al. Characterization of the phenological stages of seven macadamia nut cultivars and selection[J]. Revista Brasileira de Fruticultura, 2007, 29(3): 690-694.
[10] Rojas C M, García C V, Munoz M E L, et al. Flowering and fructification phenology in Macadamia integrifolia[J]. Acta Agronomica, 2009, 58(4): 277-284.
[11] 陆超忠, 杜丽清. 澳洲坚果种质资源描述规范和数据标准[M]. 北京: 中国农业出版社, 2008.
[12] 邹明宏, 陆超忠, 梁李宏, 等. NY/T 1687-2009澳洲坚果种质资源鉴定技术规范[M]. 北京: 中国农业出版社, 2009.
[13] 苏玉环, 王静华, 李文芹, 等. 西瓜种质资源果实性状及聚类分析[J]. 河北农业大学学报, 2008, 31(6): 21-26.
[14] Ito P J. Effect of style removal on fruit set in macadamia[J]. HortScience, 1980, 15: 520-521.
[15] Hardner C M, Peace C, Lowe A J, et al. Genetic resources and domestication of macadamia[M].// Janick J.(ed.). Horticultural Reviews, John Wiley & Sons, Hoboken, New Jersey, 2009: 1-125.
[16] Sakai W S, Nagao M A. Fruit growth and abscission in Macadamia integrifolia[J]. Physiol Plant, 1985, 64: 455-460.
[17] Trueman S J, Turnbull C G N. Effects of cross-pollination and flower removal on fruit set in macadamia[J]. Annals of Botany, 1994, 73: 23-32.
[18] Wallace H M, Vithanage V, Exley E M. The effect of supplementary pollination on nut set of macadamia(Proteaceae)[J]. Annals of Botany, 1996, 78: 765-773.
