猪源沙门氏菌携带I类整合子向人源细菌体外转移的研究
2015-10-20王学君卢春洪王源谭艾娟吕世明蒋
王学君+卢春洪+王源+谭艾娟+吕世明+蒋庆庆+符东顺

摘 要:为考察猪源沙门氏菌携带的I类整合子向人源细菌在体外接合试验中的转移频率,在体外接合试验中应用PCR方法检测细菌间I类整合子的转移。结果表明:人源沙门氏菌I类整合子转移效率在1.6×10-3 ~2.5×10-4 cfu·mL-1(供体菌)之间,人源大肠杆菌I类整合子转移效率在1.5×10-5 ~8.3×10-6 cfu·mL-1 (供体菌)之间,猪源沙门氏菌I类整合子未转移给人源金黄色葡萄球菌。因此,猪源沙门氏菌I类整合子携带耐药基因盒在体外可向人源革兰氏阴性菌发生转移,且转移频率较高。
关键词:沙门氏菌;体外接合试验; 整合子;转移
中图分类号:R446.5 文献标识码:A DOI 编码:10.3969/j.issn.1006-6500.2015.10.004
Abstract: To investigate the transfer frequency of class I integrons from swine Salmonella to human bacteria on the conjugation test in vitro, the transfertation of class I integrons between the strains was determined with PCR method. Results showed that the transfertation rate of class I integrons from swine Salmonella to human Escherichia coli were in 1.6×10-3 ~2.5×10-4 cfu·mL-1 (donor strain), and to human Salmonella were 1.5×10-5 ~8.3×10-6 cfu·mL-1 (donor strain). Class I integrons were no found in human in human Staphylococcus aureus. So, class I integrons of swine Salmonellae could transfer to gram-negative bacteria of human origin in vitro and the transfer frequency was quite high.
Key words:Salmonella; integron; transfer; conjugation test in vitro
沙门氏菌(Salmonella)是全球公认的主要动物源细菌传染病,危害人和动物健康的重要致病菌,严重威胁养殖业和食品安全[1]。整合子是介导细菌耐药性产生的主要机制之一[2],同时整合子可定位于质粒上,也可以作为转座子的一部分参与转移,使耐药基因在细菌种间和种内发生转移[3]。整合子携带耐药基因体外转移多集中在临床菌株的研究中[4],但对动物源细菌的报道较少,且整合子携带耐药基因盒在细菌间水平转移,加快了临床耐致病菌获得耐药性。本研究是将5株猪源沙门氏菌I类整合子阳性菌株与人源性不携带I类整合子的沙门氏菌、大肠杆菌和金黄色葡萄球菌各1株进行耐药基因的体外转移试验,考察I类整合子携带耐药基因能否在人源、动物源细菌间传播,为细菌耐药性的传播监测提供理论依据。
1 材料和方法
1.1 试验材料
1.1.1 试验菌株 供体菌:猪源沙门氏菌5株,含有I类整合子(贵州大学药理实验室保存)。
受体菌:人源大肠杆菌、沙门氏菌、金黄色葡萄球菌各1株(由贵阳市疾病预防控制中心惠赠),且不携带I类整合子及耐药基因盒。
1.1.2 培养基和试剂 LB肉汤、缓冲蛋白胨水(BPW)、GN增菌液、木糖赖氨酸脱氧胆酸钠琼脂培养基(XLD)、伊红美兰琼脂培养基(EMB)、贝尔德-帕克氏琼脂培养基(BP)均购自杭州博微生物技术有限公司;四环素和链霉素购自上海博微科技有限公司。Ex Taq酶试剂盒购自TaKaRa公司。
1.2 试验方法
1.2.1 体外接合试验 [5-7] 供、受体菌分别于LB肉汤中37 ℃过夜;调整菌液为2个麦氏浓度,各吸0.1 mL加入0.8 mL LB肉汤,37 ℃孵育4 h;菌液3 800 r·min-1离心2 min,弃上清液,1 mL生理盐水将沉淀混匀;吸取0.1 mL涂布于MHA平板,37 ℃过夜;经102~103倍稀释后吸取0.1 mL菌液涂布于含双抗菌药物(0.02 mg·mL-1四环素+0.05 mg·mL-1链霉素)的EMB、XLD、BP选择性培养基上,37 ℃培养18~24 h;供体菌涂布于含单抗的XLD培养基上,受体菌涂布于单抗的EMB、XLD、BP平板上,37 ℃培养18~24 h。
1.2.2 接合频率统计 供/受体菌进行选择性平板计数,当供体菌和受体菌在双抗选择性培养基中不生长的情况下,计数接合菌、供受体菌数。接合频率Fc (conjugation frequency):Fc=T/R,T为耐药性转移接合子的平均菌落数,R为受体菌平均菌落数。
1.2.3 接合子中整合子阳性菌株检测 在不同抗性平板上随机挑选 10 个单菌落,进行 PCR 验证,阳性菌株数为 I1,参照文献[8],以整合酶intI-F/intI-R为引物进行PCR扩增。上游:5′-CGATGCGTGGAGACCGAAACCTT-3′,下游:5′-GTAACGCGCTTGCTGCTTGGATGC-3′,PCR扩增条件:94 ℃预变性300 s,94 ℃变性30 s,58 ℃退火30 S,72 ℃延伸30 s,共30个循环,72 ℃终延伸10 min。产物采用2.0% 琼脂糖凝胶进行电泳分析。
1.2.4 体外I类整合子转移效率的测定 由整合子转移效率公式计算,整合子转移率 =I1/N*Fc,其中,I1/N为 PCR 阳性率,M/D为接合效率。
2 结果与分析
2.1 体外接合子筛选结果
从表1数据可知,5株猪源沙门氏菌与人源沙门氏菌的接合频率在2.1×10-3~3.5×10-4之间;与人源大肠杆菌接合频率在2.9×10-4~4.1×10-5之间;人源金黄色葡萄球菌接合频率在2.3×10-7~5×10-8之间,且有两株未检测到接合子。
2.2 接合子中I类整合子阳性菌株检测结果
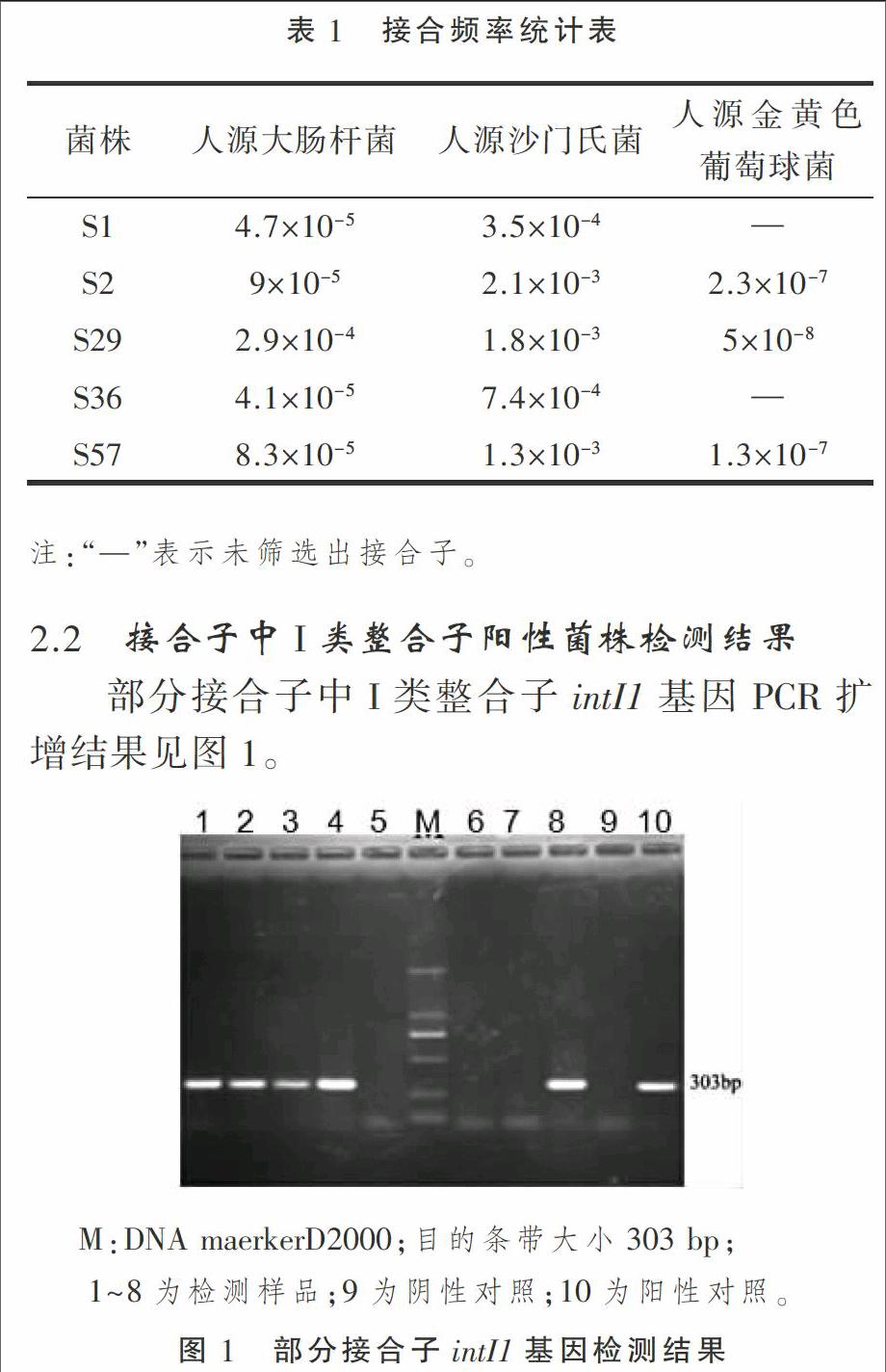
部分接合子中I类整合子intI1基因PCR扩增结果见图1。
由表2可知,5株人源沙门氏菌接合子中均检测到I类整合子阳性菌株,4株人源大肠杆菌接合子中检测得到I类整合子阳性菌株,人源金黄色葡萄球菌未检测到I类整合子。人源沙门氏菌接合子中检测得到阳性整合菌株明显高于人源其他菌株,人源金黄色葡萄球菌接合子中未检测到阳性整合菌株。
2.3 体外I类整合子转移频率结果
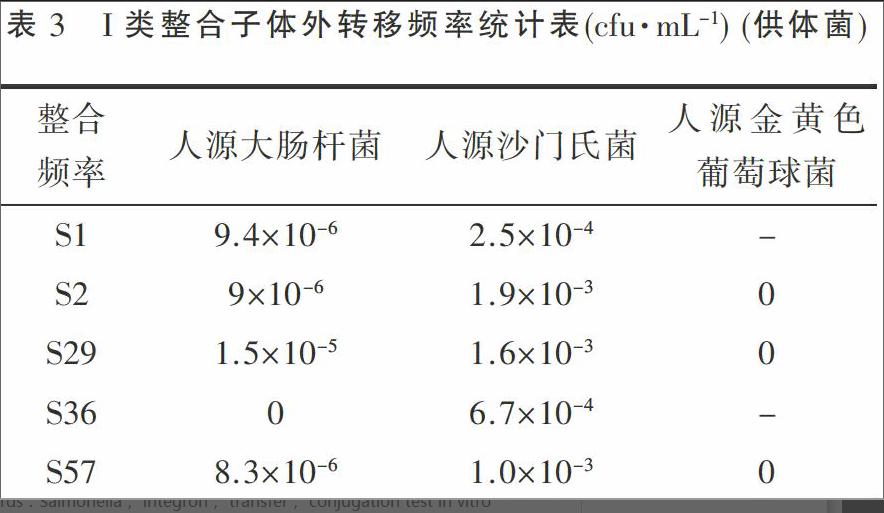
由表3可知,人源沙门氏菌I类整合子转移频率在1.6×10-3 ~2.5×10-4 cfu·mL-1 (供体菌)之间,人源大肠杆菌I类整合子转移频率在1.5×10-5 ~8.3×10-6 cfu·mL-1 (供体菌)之间,猪源沙门氏菌I类整合子未转移给人源金黄色葡萄球菌。说明沙门氏菌同种属之间转移效率高于其他人源细菌,革兰氏阴性菌之间整合子携带耐药基因盒存在转移风险,革兰氏阴性菌向阳性菌之间水平转移可能性极小。
3 结论与讨论
研究动物源性细菌整合子-基因盒系统介导的耐药机制是控制细菌耐药性传播的根源途径。本研究探讨猪源沙门氏菌向人源不同菌种之间的水平转移,从分子水平分析细菌耐药性获得及传播方式,为我国动物源性细菌耐药性传递研究积累基础资料,为指导规模猪场合理选择用药、制定干预细菌多重耐药的产生及传播措施提供了理论依据。
通过体外转移试验,发现猪源沙门氏菌与人源细菌体外接合频率在10-3~10-8之间,低于刘渠等[6]报道的接合频率10-1~10-5,而本试验中沙门氏菌同菌属细菌之间的体外接合频率高于不同菌属细菌之间的接合频率。
由猪源沙门氏菌I类整合子转移频率结果表明,整合子在人源沙门氏菌与人源大肠杆菌中存在转移,且转移频率1.6×10-3 ~8.3×10-6 cfu·mL-1 (供体菌)之间, 与文献报道一致[9]。但是人源金黄色葡萄球菌接合子中未检测到I类整合子,说明猪源沙门氏菌中I类整合子向人源革兰氏阳性菌之间体外转移可能性极小。
参考文献:
[1] TAIWO S S, FADIORA S O, FAYEMIWO S A . High antimicrobial resistance among bacterial isolates of blood stream infections (BSI) in a Nigerian University Teaching Hospital [J]. World Journal of Microbiology and Biotechnology, 2008,24(2):231-236.
[2] 魏述永,吴邓红,刘世东.细菌基因盒-整合子系统研究进展[J].动物医学进展, 2008,29(1):53-56.
[3] BARBEI D A, BAHNSON P B, ISAACSON R, et al .Distribution of Salmonella in swine production ecosystems[J]. Food Prot , 2002, 65(12): 1861-1868.
[4] LABBATE M,CSES R J,STOKES H W.The integron gene cassette sys-tem:An active player in bacterial adaptation[J].Methods Molbiol,2009,532:103-125.
[5] WASTESON Y,ROE D E, FALK K. Characterization of tetracycline and erythromycin resistance in Actinobacillus pleuropneumoniae[J]. Veterinary Microbiology,1996,48:1-2.
[6] 刘渠,刘衡川,白松涛,等. 食品中大肠埃希氏菌、沙门氏菌整合子的耐药性水平传递研究[J]. 现代预防医学,2004,31(5) :681-684.
[7] 魏峰,李杰,金山,等.鸡源大肠杆菌对氨基糖苷类抗生素耐药性检测[J].广东农业科学,2012,39(4):90-91.
[8] KRAULAND M G, MARSH J W, PATERSON D L, et al. Integron-mediated multidrug resistance in a global collection of nontyphoidal Salmonella enterica isolates[J]. Emerging Infectious Diseases,2009,15(3): 388-396.
[9] 练维,石翠,冒群,等. 整合子介导的沙门菌耐药性水平播散机制的初步研究[J]. 中国食品卫生杂志,3013,25(1):16-19.
