春季生态因子对武夷四照花光合作用的影响研究
2015-09-30郭振黄成林
郭振+黄成林


摘 要:采用Li-6400便携式光合测定仪,在春季对武夷四照花的光合日变化进行测定,通过相关分析和通径分析,得出净光合速率与影响因子之间的关系,并拟合光响应曲线,利用多元回归分析建立相应的多元方程。结果表明:武夷四照花叶片Pn的日变化呈双峰曲线型,具有典型的光合“午睡”现象;气孔导度和相对湿度是直接影响武夷四照花叶片Pn的主要因子;武夷四照花的光补偿点为26.02μmol photons·m-2·s-1,光饱和点为670.01μmol photons·m-2·s-1,补偿点、饱和点均较低,属于耐阴植物。
关键词:武夷四照花;光合作用;生态因子
中图分类号 S685.99 文献标识码 A 文章编号 1007-7731(2015)17-39-04
Effects of Ecological Factors on Photosynthesis of Dendrobenthamia angustata var.wuyishanensis in Spring
Guo Zhen1 et al.
(1Urban Construction Institute,Luan Vocational Technical College,Lu'an 237000,China)
Abstract:Using Li-6400 portable photosynthetic apparatus,photosynthesis diurnal variation of Dendrobenthamia angustata var.wuyishanensis in spring was determined. Through correlation analysis and path analysis,this paper assessed the relationship between net photosynthetic rate(Pn)and impact factors and light response curve fitting. Multiple regression analysis is used to establish the corresponding equation. The results showed that:the diurnal variation of Pn had the double-peak curve,the typical photosynthetic“nap”phenomenon;relative humidity and stomatal conductance were the most important directfactors to influence the changes of Pn. The light saturation point was 26.02μmol photons·m-2·s-1and the light saturation point was 670.01μmol photons·m-2·s-1,so D.angustata var.wuyishanensis is shade plants.
Key words:Dendrobenthamia angustata var.wuyishanensis;Photosynthesis;Ecological factors
植物总是生活在某环境之中,并且不断地与环境进行能量、物质交换[1]。在自然条件下,没有一种环境条件是恒定不变的[2],外界环境因子是影响光合作用的重要因素之一[3]。武夷四照花(Dendrobenthamia angustata var.wuyishanensis)为山茱萸科山茱萸属尖叶四照花的变种,半常绿灌木,主要分布在中国大陆的江西、广东等地的山区海拔300~1 000m的地区,多数生长于山谷、常绿阔叶林及杂林中[4]。
目前对于四照花属植物的研究多为繁殖体系方面的研究[5],而关于其生理生态特性的研究比较少见。在生理特性的研究中,主要是关于四照花属植物逆境生理研究[6],关于四照花植物净光合速率与主要生态因子之间的关系尚未见报道。为此,笔者对四照花的光合特性进行了研究,探讨影响光合作用的主要生态因子对其净光合速率的影响,以期完善四照花属植物的生理研究,为该植物更好地应用于园林配植造景提供理论依据。
1 材料与方法
1.1 材料 试验材料选取露地条件下栽植的武夷四照花5年生植株。原产地福建武夷山,引种并栽植于合肥市半岛苗圃。
1.2 方法
1.2.1 光合日变化的测定 4月下旬,在晴朗无风的天气状况下,采用Li-6400便携式光合测定仪在一天中的7:00~18:00每隔1h测定一次净光合速率(Pn,μmolCO2·m-2·s-1),选择生长健康的植株,选取该植物位于植株中部向阳面的叶片进行测定,重复3次测定。待仪器稳定后,测定叶片瞬时净光合速率值,同时对影响净光合速率值的气孔导度(Gs,mmolH2O·m-2·s-1)、胞间CO2浓度(Ci,μmolCO2·mol-1air)、
蒸腾速率(Tr,mmol H2O·m-2·s-1)、饱和蒸气压亏缺(VpdL,Kpa)、气温(Ta,℃)、叶温(Tl,℃)、大气CO2浓度(Ca,μmol CO2·mol-1 air)、空气相对湿度(RH,%)、光合有效辐射(PAR,μmol photons·m-2·s-1)等因子进行测定。为了避免因为叶片不同导致的对光照强度反应的差异性,每次取同一叶片的相同部位进行观测,取测量平均值进行各项数据分析。
1.2.2 光响应曲线的测定 与日变化同一时期进行测定,选择健康叶片,测定时每组重复3次,利用Li-6400便携式光合测定仪,光源利用仪器自带的LED红蓝光源进行设置,设定光合有效辐射(PAR)从最大值(2 000μmol photons·m-2·s-1)逐渐降低至最小值(0),测定在不同光强下的净光合速率,通过公式拟合并绘制武夷四照花的光响应曲线。在估计的光补偿点和光饱和点位置附近将光强梯度划分密些,以便更加精确的求出光补偿点和光饱和点的数值。光强的梯度划分为:2 000、1 500、1 300、1 000、800、500、300、150、100、80、50、30、0μmol photons·m-2·s-1[7]。以光照强度(PAR)为横轴,净光合速率(Pn)为纵轴绘制武夷四照花的光响应曲线,计算武夷四照花的光补偿点、光饱和点、最大表观量子效率和最大净光合速率,并分析其与各生态影响因子之间的关系。
1.3 数据处理 实验所得数据利用Microsoft Excel读取并进行初步处理,用SAS软件进行影响因子相关分析、多元逐步回归分析和通径分析,利用叶子飘的光响应新模型对光响应曲线拟合[8]。
2 结果与分析
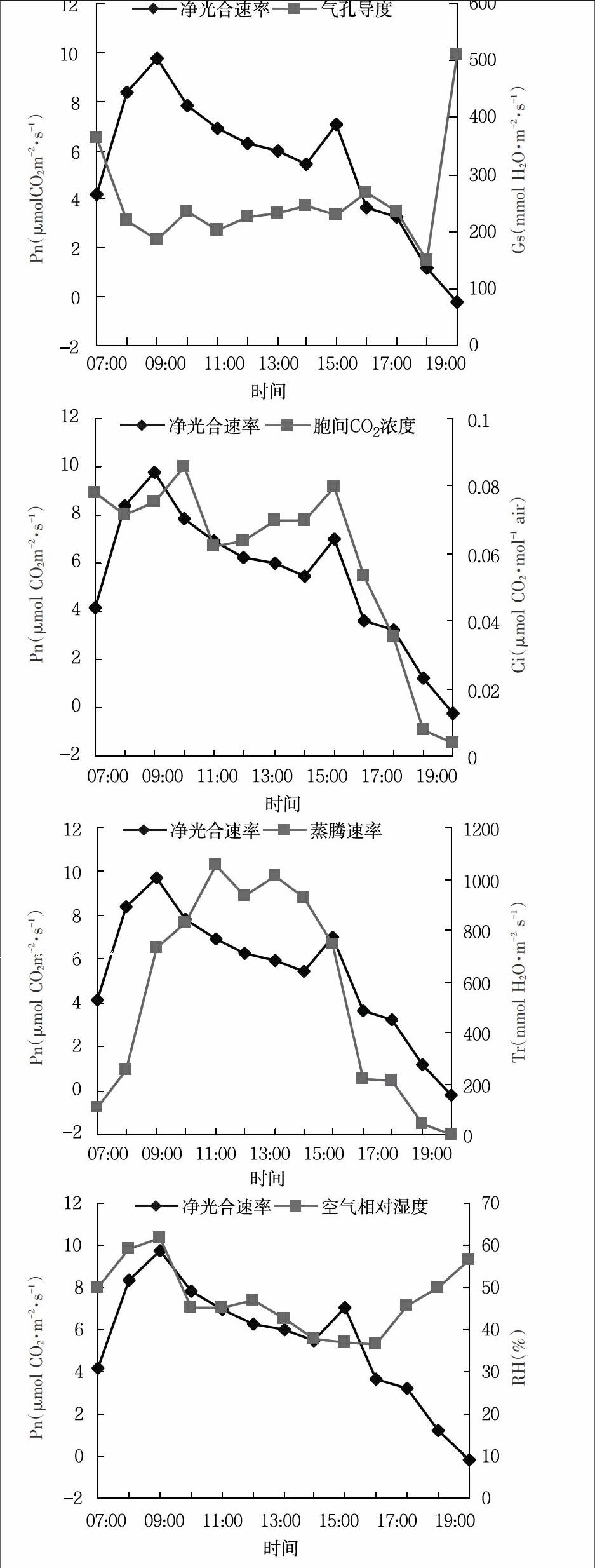
2.1 武夷四照花光合作用的日变化 在春季,武夷四照花叶片Pn随着各种生态因子的变化而表现出不同的日变化规律(图1)。由图1可知,上午7:00~9:00时叶片净光合速率随气孔导度的增加呈直线上升趋势,到9:00时,当气孔导度达到0.075mmol H2O·m-2·s-1时,净光合速率出现全天第一个高峰值,为9.73μmol CO2·m-2·s-1。之后气孔导度继续升高,在10:00时达到最大值,此时的气孔导度值是0.086mmol H2O·m-2·s-1,下午的15:00时气孔导度和净光合速率值同时达到最大值,此时的气孔导度值和净光合速率值分别是0.079mmol H2O·m-2·s-1和7.03μmol CO2·m-2·s-1,此时为一天中的第二个高峰。2次高峰之间出现很明显的光合“午睡”现象,气孔导度值和净光合速率的变化趋势体现出高度一致性且为正相关。与气孔导度同样为正相关的影响因子还有蒸腾速率和光合有效辐射。其中净光合速率和光合有效辐射的关系中,分别在9:00和15:00时出现峰值,此时的净光合速率分别是9.73μmol CO2·m-2·s-1和7.03μmol CO2·m-2·s-1,此时的光合有效辐射分别是728.82μmol photons·m-2·s-1和745.64μmol photons·m-2·s-1。和净光合速率负相关的因子是胞间CO2浓度、饱和蒸汽压亏缺和大气CO2浓度,叶面温度对净光合速率的变化影响不明显。
植物光合作用的“午睡”现象比较普遍,一般认为出现“午睡”现象的原因主要有气孔限制原因和非气孔限制2方面[9]。本实验数据表明,武夷四照花的“午睡”现象和气孔限制正相关且影响显著。根据Farquhar和Sharkey的观点[10],本实验气孔导度和胞间CO2浓度变化正相关,表明由于气孔的关闭导致CO2的供应减少,从而降低了净光合速率。
[Pn(μmol CO2·m-2·s-1)][PAR(μmol photons·m-2 s-1)]
图1 武夷四照花光合速率与不同生态因子的日变化
2.2 武夷四照花的光响应曲线 在春季测定武夷四照花的光响应曲线,当日气温26℃,CO2浓度为400.08μmol·mol-1,H2OR和H2OS平均值为22.84μmol·mol-1,测定结果用叶子飘新模型拟合。结果如图2所示,当光合有效辐射从0μmol photons·m-2·s-1逐步增加到l00μmol photons·m-2·s-1时,光照强度(X)的与净光合速率(Y)之间的变化线性相关,得出线性方程:Y=0.5432X-1.3002(R2=0.993),通过方程可计算出武夷四照花在春季的光补偿点是26.02μmol photons·m-2·s-1,光饱和点是670.01μmol photons·m-2·s-1。其光饱和点低于中性植物杉木(751.5μmol photons·m-2·s-1),属于阴性植物。光补偿点低于耐阴性较强的洒金珊瑚(29.71μmol photons·m-2·s-1),说明其对弱光的利用能力较强。
[PAR(μmol·m-2 s-1)][Pn(μmol CO2 m-2·s-1)]
图2 武夷四照花春季光响应曲线
2.3 生态因子对武夷四照花光合作用的影响
2.3.1 多元回归分析 现以Pn为因变量(Y),分别以各生态因子为自变量(Xi),即气孔导度(X1)、胞间CO2浓度(X2)、蒸腾速率(X3)、饱和蒸汽压亏缺(X4)、叶面温度温(X5)、大气CO2浓度(X6)、空气相对湿度(X7)、光合有效辐射(X8),作多元逐步回归方程:
Pn=12.389+90.2027Cond-0.023Ci+0.3516Tr-0.3797VpdL
+0.0227Ta-0.0501Ca+0.1762Rh+0.0007PAR,R2=0.9715
由多元回归分析可看出,影响净光合速率(Pn)最大的是气孔导度(Cond),其与净光合速率正相关,同样正相关的还有蒸腾速率、光合有效辐射和相对湿度。与净光合速率值负相关的因子有胞间CO2浓度(Ci)、饱和蒸气压亏缺(VpdL)和大气CO2浓度(Ca)。这些生理因子不仅直接影响净光合速率值,而且会通过间接效应影响到净光合速率,此结果与相关性分析的结果基本一致。
2.3.2 相关性 分析武夷四照花净光合速率(Pn)与各生理因子的相关分析(表1)表明:净光合速率(Pn)与气孔导度呈极显著正相关,与相对湿度呈显著正相关,与胞间CO2浓度、饱和蒸汽压亏缺、大气CO2浓度呈负相关。
表1 武夷四照花净光合速率与各影响因子间的相关系数
[\&Gs\&Ci\&Tr\&VpdL\&Tl\&Ca\&Rh\&Par\&相关
系数\&0.8362**\&-0.0732\&0.11266\&-0.1288\&0.0382\&-0.3499\&0.5025*\&0.0985\&]
注:**表示影响极显著(显著水平为0.01),*表示影响显著(显著水平为0.05)。
2.3.3 通径分析 为了进一步分析各生态因子对净光合速率(Pn)的影响,对测定结果进行通径分析,通过通径分析不仅可以反映各影响因子对于净光合速率(Pn)的直接影响,还可以通过间接通径系数揭示各因子之间的相互作用,从而了解一个因子通过另一个因子影响净光合速率(Pn)的效应[10]。通径分析表明,气孔导度对净光合速率的影响最大,总效应为0.862 5,其影响主要来自于自身的直接作用,这与相关性分析的结论一致,相关性分析中显著正相关的相对湿度在通径分析中表现相关性不大,这是因为气孔导度、胞间CO2浓度、蒸腾速率和叶面温度等因素对其施加了负效应,抵消了其对净光合速率的正影响;蒸腾速率和光合有效辐射在通径分析中表明其对净光合速率影响较大,其总效应分别是0.531 3和0.672 3,主要是因为气孔导度通过这2个因子对净光合速率施加了较大的间接影响(表2)。
表2 武夷四照花光合速率与生态因子的通径分析
[生态因子\&直接通径
系数\&间接通径系数\&Gs\&Ci\&Tr\&VpdL\&Tl\&Ca\&Rh\&Par\&Gs\&0.8362\&\&0.0236\&0.0791\&-0.042\&0.009\&0.0018\&-0.1076\&0.0619\&Ci\&-0.0732\&-0.2702\&\&-0.0358\&0.0407\&-0.0135\&-0.2341\&0.08467\&-0.0433\&Tr\&0.11266\&0.5876\&0.0232\&\&-0.1142\&0.0315\&0.17071\&-0.3635\&0.0832\&VpdL\&-0.1288\&0.27276\&0.0231\&0.0999\&\&0.0371\&0.24454\&-0.4359\&0.0719\&Tl\&0.0382\&0.2063\&0.0260\&0.0929\&-0.1253\&\&0.2788\&-0.4312\&0.0659\&Ca\&-0.3499\&-0.0043\&-0.0489\&-0.0549\&0.0900\&-0.0304\&\&0.2733\&-0.0472\&Rh\&0.5025\&-0.1791\&-0.0123\&-0.0814\&0.1117\&-0.0327\&-0.1903\&\&-0.035\&Par\&0.0985\&0.5258\&0.0322\&0.0951\&-0.0941\&0.0255\&0.1677\&-0.1786\& \&]
3 讨论与结论
近些年来,随着光合测定仪器的不断更新[11],实验方法的不断成熟[12],以及数据分析的不断深入[13],对植物光合生理的研究更加深入和全面[14]。植物光合作用的特征和各因子对光合作用效率的影响逐渐成为植物光合研究的重点[15]。
本实验表明,春季武夷四照花叶片净光合速率表现出明显的双峰曲线,净光合速率的峰值分别出现在9:00和15:00,其数值分别为9.73μmol CO2·m-2·s-1、7.03μmol CO2·m-2·s-1;武夷四照花气孔导度和蒸腾速率的日变化也为双峰曲线,峰值大小和时间随环境变化而变化,其气孔导度和蒸腾速率的最大值分别为0.086mmol H2O·m-2·s-1、2.605mmol H2O·m-2·s-1,
相关分析和通径分析表明,净光合速率(Pn)与气孔导度呈极显著正相关,与相对湿度呈显著正相关,与胞间CO2浓度、饱和蒸汽压亏缺、大气CO2浓度呈负相关。气孔导度对净光合速率的影响最大,总效应为0.862 5,其次是蒸腾速率和光合有效辐射。
另外,据光响应曲线,春季武夷四照花的光补偿点为26.02μmol photons·m-2·s-1,光饱和点为670.01μmol photons·m-2·s-1,属于耐阴性植物,适合在适当遮光的条件下栽植。关于武夷四照花抗旱性或者温度胁迫实验,应在今后的试验中作进一步研究。
参考文献
[1]许大全,张玉忠,张荣铣.植物光合作用的光抑制[J].植物生理学通讯,1992,28(4):237-243.
[2]许大全,沈允钢.植物光合作用效率的日变化[J].植物生理学报,1997,23(4):410-416.
[3]坎内尔M G R,拉斯特F T.树木生理与遗传改良[M].熊文愈,吴贯明,译.北京:中国林业出版社,1981.
[4]韩维栋.四照花类群种质资源及其开发利用[J].中国野生植物资源,1993,11(2):37-40.
[5]余道平,彭启新,李策宏,等.峨眉四照花育苗技术及苗高生长节律研究[J].西北林学院学报,2008,23(1):111-114.
[6]彭诚,易咏梅.冷冻胁迫对狭叶四照花苗丙二醛等生理特性的研究[J].湖北民族学院学报(自然科学版),2007,25(2).
[7]郭振,黄成林.武夷四照花光合生理生态特性的研究[J].安徽农业大学学报,2013,06.
[8]叶子飘.光响应模型在超级杂交稻组合——II优明86中的应用[J].生态学杂志,2007,08:1323-1326.
[9]黄成林,吴泽民.遮荫条件下绞股蓝光合作用特点的研究[J].应用生态学报,2004,15(11).
[10]FARQUHAR G D,SHARKEY T D.Stomatal conductance and photosynthesis[J].Ann Rev Plant Physiol,1982,33:317-345.
[11]刘宇锋,萧浪涛,童建华,等.非直线双曲线模型在光合光响应曲线数据分析中的应用[J].中国农学通报,2005,121(8):76-79.
[12]黄成林,赵昌恒,傅松玲,等.安徽休宁倭竹光合生理特性的研究[J].安徽农业大学学报,2005,32(2).
[13]郑炳松,金爱武.雷竹光合特性的研究[J].福建林学院学报,2001,21(4):355-362
[14]郭志达,张宏达,李志安,等.鹅掌楸苗期光合特性的研究[J].生态学报,1999,19(2):164-169.
[15]王克勤,王斌瑞.土壤水分对金矮生苹果光合速率的影响[J].生态学报,2002,22(2):206-214.
[16]陈根云,俞冠路,陈悦,等.光合作用对光和二氧化碳响应的观测方法讨论[J].植物生理与分子生物学学报,2006,32(6):691-696.
[17]付为国,李萍萍,卞新民,等.镇江北固山湿地芦苇光合日变化的研究[J].西北植物学报,2006,26(3):496-501.
(责编:张宏民)
