基于地统计学的云斑天牛幼虫种群时空动态1)
2015-09-18胡瑜珩
胡瑜珩
(北京林业大学,北京,100083)
武海卫
(山东省林业科学研究院)
许志春
(北京林业大学)
云斑天牛(Batocera horsfieldi(Hope))隶属于鞘翅目(Coleoptera)天牛科(Cerambycidae)是我国重要的蛀干害虫[1]。在我国云斑天牛为2~3年1代[2],由于整个世代中绝大部分(>90%)的时间以幼虫隐蔽于树干中生活,本身并不具备扩散的能力[3],其垂直分布规律与时空动态都是由于成虫的生活习性,如:产卵、扩散以及补充营养的习性等决定的[4]。近年来,由于云斑天牛防治策略的需要,很多学者都以其空间分布格局为背景来试图研究出一套合理的防治虫害发生的有效措施,即基于环境友好型的特定样地的综合防治(IPM)措施[5]。但这些研究主要集中在对杨树云斑天牛空间格局的阐释上,且运用的方法是传统的指标调查法[3-4]。虽然作为调查昆虫空间动态更为优越的地统计学方法在国内外已经运用于众多昆虫的空间格局分析上,但在云斑天牛的调查上依然少见。
明确气象因子对昆虫产卵及幼虫孵化的影响有助于对种群密度与结构演变的分析研究[6-7],而温度和降水是决定病虫害空间格局的主要因素[8]。本研究利用2013—2014年6—8月份的野外观测数据,结合云斑天牛生活史特性,分析云斑天牛幼虫在白蜡树(Fraxinus chinensis)上的分布规律,揭示其年际变异动态,并基于地统计学分析方法对比研究空间格局动态变化规律,试图找出云斑天牛空间上的扩散方向,旨在了解云斑天牛在白蜡树上的危害特性,为科学地防治该虫害提供依据。
1 试验地概况
试验地设在山东省林业科学研究院盐碱地造林试验站,位于渤海湾西南岸、寿光市北部,地理位置为东经118°40',北纬37°11',平均海拔35 m,属暖温带季风区大陆性气候,年平均气温13℃,年均降水量600 mm,年均蒸发量2200 mm,地下水位2~3 m,全年无霜期 195 d[9]。植被类以白蜡、杨(Populus spp.)、旱柳(Salix matsudana)为主,还有少量桑树(Morus alba)、榆(Ulmus pumila)、刺槐(Robinia pseudoacacia)、柽柳(Tamarix chinensis)。
2 材料与方法
2.1 调查方法
根据白蜡林中云斑天牛危害情况将其分为轻度受害、中度受害和重度受害林,3个调查样地受害白蜡的胸径区间在 7.10~9.84 cm,树高区间在 7.32~10.08 m。根据其有虫株率分为轻度(8.8%)、中度(22.7%)和重度(32.7%) 危害林,各林分抽样调查的数量分别为179、137、98株,总计样本量为414株。
2013年6—8月份,根据试验地的大小及白蜡树的分布情况,采取逐株取样调查的方法,从白蜡树的基部向上至树梢,每株分别按1 m以下、>1~2 m、>2~3 m和>3 m这4个高度区域划分,统计每个高度区域内的排粪孔数量,并以此作为密度指标,同时在坐标纸上记录样地中每株白蜡树的相对空间位置。
2014年同期基于上一年的调查数据,在选定的相同林分样地内对云斑天牛幼虫在白蜡树上的各个高度区域内的虫口密度进行统计。
2.2 数据处理
2.2.1 云斑天牛幼虫垂直分布、时间动态
调查发现,云斑天牛在试验地每年6—7月份为产卵盛期,第2年8月份老熟幼虫化蛹,1月份以后开始破蛹,之后在蛀道内越冬,第3年5月份羽化进入下一世代生活史,并在树干上形成一个圆形或椭圆形的孔状结构[2]。当年产生的云斑天牛排粪孔和羽化孔均显亮黄色,与往年的相比较易识别,所以可根据2013与2014年的羽化孔数分别对2011和2012年的排粪孔数做近似估计(图1)。利用 SPSS Statistics 22.0分别对不同样地内云斑天牛幼虫4个高度区域的垂直分布和在不同受害林分1 m以下各年的虫口数量进行单因素方差分析(ANOVA)与多重比较(LSD),从而分别判别幼虫垂直分布与年际变异的差异显著性(α=0.05)。
获取当地2011—2014年6—8月份的气候资料,包括温度与降水,分析云斑天牛虫害发生与气候因子之间的相关性。所有统计分析都基于SPSS Statistics 22.0 软件。
2.2.2 云斑天牛幼虫空间动态
(1)半变异函数与变异函数图
半变异函数是地统计学中研究空间变异性的关键函数,是半变异函数正则化变量的拟合与计算、离散方差的分析以及方差估计等地统计方法的前提,同时也可以描述区域化变量的重要性质。其公式为:
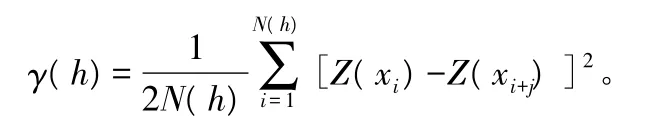
式中:γ(h)为相对于距离h的估计半方差函数值;N(h)为被 h分割的数据对(xi,xi+h)的对数,Z(xi)和Z(xi+j)分别为xi和xi+h处样本的测量值,h为分割2 样点的距离[10-11]。
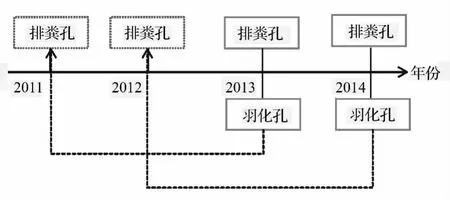
图1 云斑天牛幼虫数量年际变异模拟示意图
地统计学中变异函数理论模拟昆虫种群空间格局特征常用的模型有表征所研究种群为聚集分布的球型、指数型、高斯型,以及表征种群呈随机分布或均匀分布的直线型。
模型的基台值、偏基台值、空间依赖范围即变程和块金值用来描述空间相关性。采用如下公式计算空间依赖性(DD):

式中:C为偏基台值,C0为块金值。
(2)空间分布图的生成
利用Gs+软件普通Kriging插值法对云斑天牛幼虫数量进行插值分析,通过Surfer 8.0软件得到空间分布的等值线与矢量叠加图。
3 结果与分析
3.1 云斑天牛幼虫的垂直分布与年际差异
对不同受害林分(轻度、中度、重度)和综合样地幼虫垂直分布的差异显著性检验结果显示(表1):云斑天牛幼虫在白蜡树上的分布具有明显的垂直分布特征(P<0.05),主要在基部往上1 m以内的高度集中分布,其他高度的天牛分布量均较低。故只对树干0~1 m内云斑天牛幼虫数量的年际变异进行差异显著性分析。结果显示,2013年各受害林分的虫口数量都显著高于其他年份(P<0.05),在重度危害林中虫口密度几乎可达1头·株-1,其他年份之间除重度危害林中2012年虫口数量(0.5头·株-1)显著高于其余2年外,其他林分各年际间虫口数量的差异均未达到显著水平(P>0.05),虫口密度一般小于0.2头·株-1(表1)。
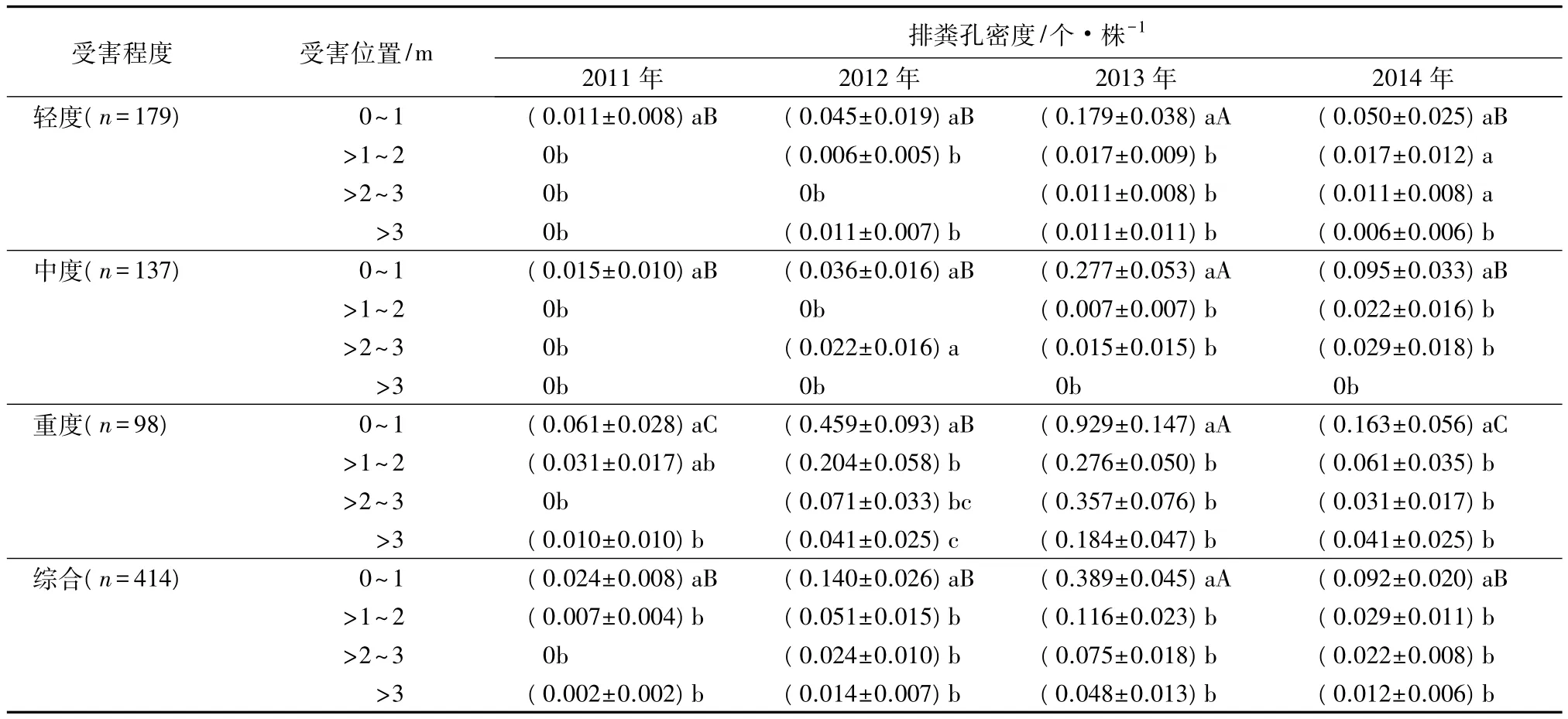
表1 2011—2014年不同受害林分云斑天牛排粪孔数量
3.2 云斑天牛幼虫数量与气象因子的关系
由2010—2014年6—8月份逐月气象数据资料(表2)可知:5年间6、7月份的月均温变化不大,分别为(25.2±0.3)、(27.4±0.1)℃;而 8 月份温度数据变化较大,其值为(26.5±0.6)℃,尤其是 2013 年 8月份的温度显著高于其余4年的同期温度((25.9±0.4)℃),在28℃以上。同期降雨量在5年间的变化要较气温大,相比而言,6月份降雨量最稳定,维持在(56.5±8.7)mm 水平上;8月份降雨呈现比较规律的“大小年”波动变化趋势,因而降雨量也相对稳定,为(156.2±48.4)mm;但是 2013 年 7 月份降雨量(454.4 mm)显著高于其余4年的同期降雨量((211.2±50.1)mm)。
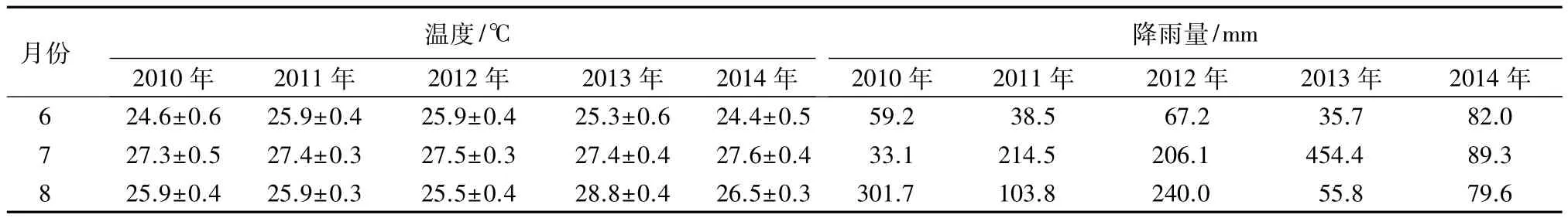
表2 2010—2014年山东寿光盐碱地实验林场6—8月份逐月气象数据
不同受害白蜡林内云斑天牛幼虫垂直分布的年际变异调查数据显示(表1):2013年各受害林分的虫口数量都显著高于其他年份(P<0.05)。这很可能与当地的气象因子有直接的关系。首先,就温度而言,2013年6—8月份的月平均气温(27.2℃)要比其余4年高1℃左右,尤其是8月份的温度差异更为显著(表2)。因此,2013年幼虫孵化所需的有效积温较其余3年就更容易达到从而成功孵化形成1龄幼虫,之后幼虫再向木质部取食形成排粪孔;另一方面,8月份的高温也可能会使少数1龄幼虫未达到其固定生活史而早先羽化飞出,从而减小了2014年检测的羽化孔数量,亦即减少了2012年排粪孔的观测数;其次,从降雨量上看,6、8月份的降雨量均较稳定,但7月份的降雨量从2010到2014呈现显著的单峰趋势,并在2013年达到最大值454.4 mm(表2),而云斑天牛产卵盛期一般在6—7月份,孵化期在7—8月份,雨水冲刷卵块或者幼虫均不利于云斑天牛的存活。而前3年7月份较小的降雨量保证了云斑天牛成虫在白蜡树上产卵以及1龄幼虫向木质部蛀食的成功率,相反2013年同期较大的降雨就不利于成虫产卵以及1龄幼虫的存活。2010—2014年间的降雨次数与降雨量数据表明,2013年7月份出现的单日降雨量超过50 mm的暴雨天气就有4次,其余4年均偶见一次甚至未见,尤其是2013年7月1日发生了日降雨量达到168 mm的大暴雨,这就使得成虫很难在白蜡树上刻槽产卵。因此,2014年云斑天牛的虫口密度就有明显的回落。
3.3 云斑天牛幼虫空间扩散动态
对2013—2014年不同受害林分云斑天牛幼虫空间格局分析可知(表3):2年间各林分内昆虫空间分布的变异函数理论模型并不相同,但是均表现为较强的空间聚集性(C/sill>0.64);2014年模型的拟合度(R2)要比2013年普遍偏低,但是其空间依赖性变化不大甚至还高于2013年的轻度危害林,多达99.9%的空间变异性都来自于空间自相关现象,证明2014年云斑天牛在轻度危害样地中的空间聚集性很强(>76%);相比而言,2014年昆虫分布的空间依赖范围明显变小,从2013年的6.4~7.8 m变为2014年的不足2 m,说明云斑天牛种群数量在2014有所降低。

表3 2013—2014年不同受害林分云斑天牛幼虫空间格局年际变异
云斑天牛幼虫在不同受害林分2年间的空间分布如图2所示:虽然中度危害林内2014年单株虫口为3.9头,高于2013年的3头,但在2014年各林分内空间依赖范围、危害面积要明显小于前1年,即2013年云斑天牛在白蜡林内成片状区域发生,而2014年表现为零星分布或者少有的小区域片状分布,所以2013年各样地内虫口密度要比2014年高(图2)。在轻度危害林中,2013年云斑天牛幼虫主要集中分布于样地的南部与东部地区,西部与北部分布量不多,且聚集性不强(图2)。但通过2014年的调查发现,云斑天牛幼虫有明显的由东向西,由南北边缘向中间聚集扩散的趋势,从而2014年幼虫主要集中于西部中央地带,其他地方都是零星状或者聚集形态不明显的斑块状分布;中度危害林内,2年间除受虫害程度明显变小外,扩散动态变化并不明显,但同样可以看出,南北方向边缘的幼虫有向中部,东部幼虫有向西部靠拢的趋势;与前两者不同,重度危害林2013年的幼虫主要集中分布在边缘地带,但在2014年幼虫表现出自南向北,自西向东明显的扩散趋势,最后集中分布于样地的东北角,其余地方呈现零星分布(图2)。
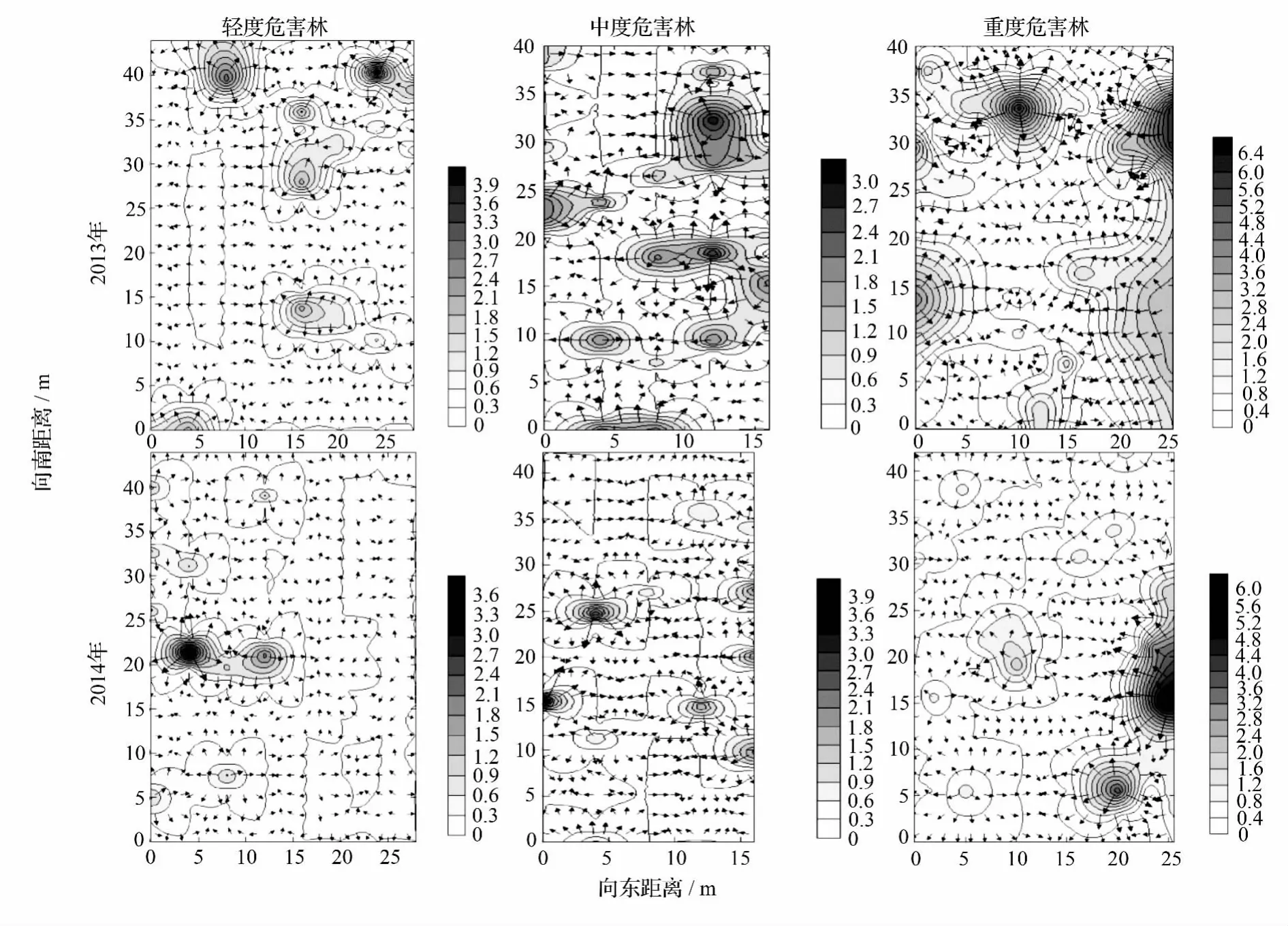
图2 不同受害林分云斑天牛幼虫空间格局年际变异(2013—2014年)
4 讨论
云斑天牛成虫在白蜡林内的产卵习性决定了云斑天牛幼虫的垂直分布特征,即一般集中分布在树干1 m以下。刁志娥等[12]在距样地30多km的山东省东营市对云斑天牛在白蜡树上的生活特性所做的调查也表明:云斑天牛成虫一般将卵产于1.5 m以下或主干分叉;其实,云斑天牛不仅在白蜡树上表现为在树干基部区域刻槽产卵[11],对许多杨树上的研究也表明1 m以下刻槽产卵量与1 m以上的刻槽产卵量具有显著差异(a=0.05)[4,13]。
气象因子,主要是温度与降水,对云斑天牛幼虫危害的年际动态具有重要的影响(表1,表2)。影响昆虫种群年际动态的因子除了林分本身的结构特征如林龄、树高、胸径、郁闭度等,还包括地理位置如坡位、坡向等,海拔,气候条件,天敌与营林措施等[14-15]。明确气象因子对昆虫产卵及幼虫孵化的影响有助于对种群密度与结构演变的分析研究[6-7],而温度和降水是决定病虫害空间格局的主要因素[8]。调查数据显示,在2013年7月份,即成虫产卵期或者幼虫的孵化期,由于降雨过多尤其是单次降雨量太大就会对卵块和幼虫形成冲刷,不利于幼虫成功向木质部蛀食,从而会降低次年幼虫的种群数量;相反2010—2012年较低的降雨使得2013年虫口密度逐渐累计,白蜡林分因此受害加剧(表 1,表 2)。Faccoli[16]的研究表明,云杉八齿小蠹(Ips typographus)所造成的损害与上年度3—7月份降雨量之间成反比关系。习妍等[17]也发现6月份合理的人工降雨会抑制松材线虫病的发生。降雨对昆虫种群空间格局的影响可以分为直接作用和间接作用[18]:一方面,在产卵期与孵化期1龄幼虫未进入木质部之前强降雨的机械冲刷作用具有直接的致死效应[7,19];另一方面,频繁降雨形成的高湿环境或者持续干旱形成的缺水环境不仅会直接影响昆虫的生长发育、繁殖及其产卵和取食行为,而且还会通过影响寄主植物而间接作用于昆虫,从而反映在昆虫种群密度以及空间分布形态上[20]。
云斑天牛幼虫数量在2013年明显高于其它年份,这不仅与降雨有关,而且与2013年当年较高的温度有密切关系。Jönsson et al.[21]调查瑞典南部云杉八齿小蠹生活习性时发现,从卵到成虫发育阶段的虫口密度随温度的增加而减少,从而有可能促成其夏季的第二季分蜂活动。翁少容[22]通过统计分析气温与降雨对松墨天牛(Monochamus alternatus)发生面积的关系,发现温度能够显著影响其发生面积并呈现出正相关的关系,但降雨的影响未达到显著水平,这有可能是因为福建当地降雨量较稳定并非限制因素所致。温度在昆虫的越冬、发生世代、迁移扩散、空间分布以及发生物候方面都有至关重要的影响[23-24],例如,甘蓝角果象甲(Ceutorhynchus obstrictus)原本在冬季低温条件下是不能越冬存活的,由于近年来气候变暖[23]的趋势愈加明显,该虫在加拿大北部建立种群并为害油料作物[25];安志芳等[26]对灰飞虱(Laodelphax striatellus)做低温胁迫试验发现,虽然低温的短期胁迫不会对其种群的发生发展产生抑制,但低温会显著延长其若虫发育所需的时间。
2013到2014年,不同林分云斑天牛幼虫的空间分布型均为聚集性,除中度危害林的扩散趋势不太明显外,其他样地扩散趋势很明显,南北方向上有一致的自南向北扩散趋势,但东西方向扩散方向相反。从地理位置上看,由于重度危害林在最西侧,中度次之,轻度在最东侧,所以3块样地合起来看,在地理上云斑天牛的扩散方式是由南向北,由两侧向中间中度危害林聚集(图2)。造成昆虫空间格局扩散的原因有3个方面,即:外界力,如风向、水力、人力等;中心虫源扩散;生活习性,如产卵习性等[27]。我国山东地区夏季盛行偏南风,所以可能会影响昆虫在产卵时的习性,从而由南向北扩散。但是由于调查样地数以及调查年份有限,很难解释清楚为何导致这样的扩散格局,因此进一步研究有关扩散动态很有必要。
由于2014年6—8月份降雨量及降雨次数均较其他年份低(250 mm),而同期温度(26.2℃)与2010—2012年的温度(26.3℃)基本一致(表 2),当地管理措施与往年并无差异,所以这给云斑天牛幼虫孵化与存活创造了良好的栖息环境。虽然2014年虫口密度低,刻槽产卵率可能也较低,但这种适宜的低潮高温条件可能会导致极高的1龄幼虫越冬存活率[20,28]。加之 2013 年的虫口密度较大(图 2),该批幼虫在2015年5—6月份才老熟,因此相当数量的蛹有可能还在林木的木质部蛀道里,2015年很可能是云斑天牛大发生的年份。林场应以2013年幼虫空间格局矢量图(图2)为依据,尽快重点检查各林分幼虫聚集点林木的健康状况,做好防控的准备。
[1]陈世骧,谢蕴贞,郑国藩.中国经济昆虫志:天牛科[M].北京:科学出版社,1959:84-85.
[2]刁志娥.白蜡树害虫发生危害及防治研究[D].泰安:山东农业大学,2005.
[3]李建庆,杨忠岐,张雅林,等.杨树上云斑天牛种群的空间格局及抽样技术[J].昆虫学报,2009,52(8):860-866.
[4]杨桦,杨茂发,杨伟,等.杨树云斑天牛幼虫空间分布格局及生息坑道[J].四川农业大学学报,2010,28(2):148-152.
[5]Karimzadeh R,Hejazi M J,Helali H,et al.Analysis of the spatiotemporal distribution of Eurygaster integriceps(Hemiptera:Scutelleridae)by using spatial analysis by distance indices and geostatistics[J].Environmental Entomology,2011,40(5):1253-1265.
[6]赵成章,李丽丽,王大为,等.黑河上游天然草地蝗虫空间异质性与分布格局[J].生态学报,2012,32(13):4166-4172.
[7]章一巧,刘永华,宗世祥,等.基于GS+的大黄枯叶蛾卵的空间分布关系[J].林业科学,2013,49(10):100-105.
[8]Graham A.The current status of the legume fossil record in the Caribbean region[M]//Herendeen P S,Dilcher D L.Advances in legume systematics:the fossil record.Kew:The Royal Botanic Gardens,1992:161-167.
[9]房用,刘德玺,马昭让,等.滨海盐碱地大果沙棘适应及耐盐碱试验[J].东北林业大学学报,2005,33(2):38-39.
[10]Lecoustre R,Fargette D,Fauquet C,et al.Analysis and mapping of the spatial spread of African cassava mosaic virus using geostatistics and kriging technique[J].Phytopathology,1989,79(9):913-920.
[11]Liebhold A M,Rossi R E,Kemp W P.Geostatistics and geographic information systems in applied insect ecology[J].Annual Review of Entomology,1993,38:303-327.
[12]刁志娥,丁福波.云斑天牛在白蜡树上的发生与防治研究[J].华东昆虫学报,2004,13(2):49-52.
[13]钱范俊,袁俊杰,杜夕生.云斑天牛产卵刻槽在杨树树干上的分布规律[J].中南林学院学报,1997,17(3):82-85.
[14]张蓉,冷允法,朱猛蒙,等.基于地统计学和GIS的苜蓿斑蚜种群空间结构分析和分布模拟[J].应用生态学报,2007,18(11):2580-2585.
[15]陈顺立,杜瑞卿,高宛莉,等.影响武夷山景区松墨天牛种群动态变化的因素分析[J].昆虫学报,2010,53(4):427-435.
[16]Faccoli M.Effect of weather on Ips typographus(Coleoptera Curculionidae)phenology,voltinism,and associated spruce mortali-ty in the southeastern Alps[J].Environmental Entomology,2009,38(2):307-316.
[17]习妍,牛树奎.气候要素对松材线虫病疫情的影响研究[J].林业资源管理,2008(4):70-76.
[18]党志浩,陈法军.昆虫对降雨和干旱的响应与适应[J].应用昆虫学报,2011,48(5):1161-1169.
[19]季荣,谢宝瑜,李哲,等.基于GIS和GS的东亚飞蝗卵块空间格局的研究[J].昆虫学报,2006,49(3):410-415.
[20]赵静,赵鑫,王玉军,等.烟盲蝽及其天敌蜘蛛空间格局的地统计学分析[J].生态学报,2010,30(15):4196-4205.
[21]Jönsson A M,Harding S,Bärring L,et al.Impact of climate change on the population dynamics of Ips typographus in southern Sweden[J].Agricultural and Forest Meteorology,2007,146(1/2):70-81.
[22]翁少容.松墨天牛发生面积与气温及雨量的关系[J].福建林业科技,2006,33(1):94-109.
[23]董兆克,戈峰.温度升高对昆虫发生发展的影响[J].应用昆虫学报,2011,48(5):1141-1148.
[24]Intergovernmental Panel on Climate Change.Climate change 2007:the physical science basis,working group I contribution to the fourth assessment report of the IPCC[M].New York:Cambridge University Press,2007.
[25]Cárcamo H A,Herle C E,Otani J,et al.Cold hardiness and overwintering survival of the cabbage seedpod weevil,Ceutorhynchus obstrictus[J].Entomol Exp Appl,2009,133(3):223-231.
[26]安志芳,张爱民,刘向东.低温胁迫对灰飞虱种群生长发育与繁殖的影响[J].应用昆虫学报,2011,48(5):1309-1313.
[27]江苏农学院植保系昆虫教研组.昆虫的扩散和迁移[J].昆虫知识,1976(1):28-30.
[28]Bale J S,Masters G J,Hodkinson I D,et al.Herbivory in global climate change research:direct effects of rising temperature on insect herbivores[J].Global Change Biology,2002,8(1):1-16.
