长白山北坡生态交错带主要树种生长对气候变化和干扰的响应1)
2015-08-08覃鑫浩
覃鑫浩
(国家林业局调查规划设计院,北京,100714)
近100 a来,全球气温显著升高[1]。中国东北地区也不例外,温度增高了约2℃[2],大气模型预测,21世纪末温度还会升高约 4.6℃[3]。气候变暖会作用于树木的生理活动,直接影响树木的生长[4]。另外,气候变暖而引起的干旱、火灾、洪涝和飓风等极端气候(干扰)事件的频率变化,也会间接影响树木生长以及森林动态[1],因此,有必要加强树木生长和动态对气候变化响应的研究,为气候变化背景下的森林可持续利用和管理提供科学依据。
生态交错带的生境异质性较高,通常是一些树种的极限分布区,因此,交错带内树木的生长对环境变化极为敏感[5]。对交错带群落结构以及树木生长对环境变化(气候及干扰)响应的研究,能够揭示交错带的群落特点和动态变化规律,反映出气候变化对森林结构的影响。树木年轮学是研究树木生长对气候变化的重要方法,也是重建森林干扰历史的重要手段。
长白山北坡生态系统完整,植被类型丰富,沿海拔变化形成了众多的生态交错带。以往的研究多关注于岳桦—苔原交错带的林线动态以及各主要树种红松(Pinus koraiensis)、落叶松(Larix olgensis)、鱼鳞云杉(Picea jezoensis)在海拔分布上、下限,径向生长对气候变化的响应[6-9],而森林干扰历史的重建也多集中于阔叶红松林[10]。将生态交错带作为研究区域,探讨气候变化对树木生长和森林动态直接和间接作用的研究还比较缺乏。因而,文中关注于长白山北坡的阔叶红松林—云冷杉林生态交错带,研究了该交错带内优势树种红松径向生长与气候因子(温度和降水)的关系,同时利用另外两个主要树种鱼鳞云杉和臭冷杉(Abies nephrolepis)的年龄结构和生长变化百分率重建了交错带的干扰历史,从而揭示该区域影响树木生长的主要气候要素以及森林动态规律,以期为森林保护和林业管理应对气候变化提供数据资料和科学参考。
1 研究区概况
本研究试验地设立于长白山自然保护区内(41°31'~42°29'N,127°8'~128°53'E),长白山属于温带大陆性山地气候,夏季温暖多雨,冬季寒冷漫长。山顶和山下气候差异大,年平均气温分别为2.8℃和7.3℃,年平均降水量为750 mm。降水多集中在夏季(6—8月份),约占全年降水量的70%以上。长白山的地貌属于典型的火山地貌,土壤垂直带谱明显,自上而下为高山苔原土带(2 000 m以上)、亚高山疏林草甸土带(1 700~2 000 m)、山地棕色针叶林土带(1 100~1 700 m)和山地暗棕色森林土带(1 000 m以下)。
长白山的植被带谱明显,自上而下为高山苔原带(2 000 m以上)、岳桦林(1 800~2 000 m)、云冷杉林(1 100~1 800 m)和阔叶红松林(500~1 100 m),每个植被带谱间都形成一定区域的生态交错带。主要树种包括了红松、鱼鳞云杉、臭冷杉、蒙古栎(Quercus mongolica)、青杨(Populus ussuriensis)、紫椴(Tilia amurensis)、水曲柳(Fraxinus mandshurica)、白桦(Betula platyphylla)、山杨(Populus davidiana)和色木槭(Acer mono)等。
2 材料与方法
2011年夏天,在长白山的阔叶红松林—云冷杉林交错带(海拔1 000~1 300 m)设置了6块样地(海拔每间隔50 m设置一块),样地面积为40 m×40 m。在群落调查时,将样地划分为5 m×5 m的小样方,记录样地内胸径≥1 cm树木种类、胸径和树高。通过轮枝测定法来确定鱼鳞云杉(422株)和臭冷杉(618株)幼苗(树高<1.3 m)的树龄。
在年轮取样时,将每块样地划分为4个20 m×20 m的亚样方,统一选取左下角(西南方向)的亚样方,用生长锥锥取亚样方内所有胸径大于3 cm的鱼鳞云杉和臭冷杉。为了准确地获取上述两个树种的树龄,每棵植株在树干基部钻取1个年轮条,保留过髓心的样品,共采集到有效的年轮样品鱼鳞云杉61个、臭冷杉106个。同时在海拔1 300 m的样地内,选取15株干形通直、无病虫害迹象且处于林冠层的红松,在胸高1.3 m处锥取年轮条,每棵植株锥取两根年轮条。
将年轮样品带回实验室晾干,安装在木槽上用砂纸打磨直至年轮边界清晰。在显微镜下进行年龄的测定,并进行交叉定年以确保每个样品年龄的准确性[11]。用LINTAB年轮分析仪测量宽度,精度为0.01 mm。最后使用软件COFECHA来检验定年结果[12],并进行修订。
数据分析:通过计算树种的重要值(IV)来判断树种在群落中的重要程度,重要值=(相对多度+相对显著度+相对频度)/3,将所有样地的重要值平均,来判断各树种在该交错带内的重要性。在分析树龄结构时,将鱼鳞云杉、臭冷杉幼苗和成树的树龄以10 a为一龄组进行统计,以公顷(hm2)为单位进行数量统计。
生长释压分析是研究森林干扰历史的有效方法,该方法通过计算年轮宽度序列的生长变化百分率,来判定生长释放(干扰事件)的发生,采用传统的生长变化百分率(Percentage Growth change,GC)来重建干扰历史[13]。具体公式如下:GC=[(M2-M1)/M1]×100%,其中GC表示前后10 a生长的百分比变化,M1和M2分别表示前10 a和后10 a的年轮宽度平均值,当GC超过100%被视为一个生长释放。以10 a为一个时间段(1820年开始),计算每个时间段内,臭冷杉和鱼鳞云杉样本发生生长释放事件的比例(即该时段内发生生长释放的样本量占总样本量的比例),将两个树种生长释放比例平均后来判断整个交错带受到干扰的程度。
遵循树木年轮学传统的分析和统计方法[11],运用ARSTAN软件采用步长40 a的样条函数去除生长趋势(受树龄、遗传以及干扰影响),并对去趋势的序列以双重平均法合成标准年表(STD)。又以时间序列的自回归模型对去趋势的序列进行再次标准化,以双权重平均法将差值序列合成差值年表(RES)。文中选取保留气候高频信号较多的差值年表进行红松生长与气候关系的分析。差值年表的公共区间为1930—2010年,可靠年表区间为1923—2010年,年表的标准差为 0.23,平均敏感度为 0.18,一阶自相关系数为-0.04,树与树之间的相关系数为0.61,信噪比为 6.80,样本总体代表性为 0.89,第一主成分所占方差量为41.6%。
应用DENDROCLIM2002软件,采用响应函数来分析树木生长与气候因子的关系[14],分析了年轮宽度与上一年6月份到当年9月份气候变量(月平均温度和月降水量)的关系,气候数据来源于长白山地区松江气象站。
3 结果与分析
3.1 群落物种组成
群落中重要值最大的4个树种为臭冷杉、鱼鳞云杉、红松和落叶松(表1)。臭冷杉的数量多和出现频率高决定了其在群落中的优势地位;红松和落叶松在群落中的优势主要体现在个体大,故胸高断面积大;鱼鳞云杉在个体数量以及出现频率都有一定的优势。
3.2 年龄结构和生长释放
臭冷杉的年龄结构呈多峰型,在1900—1910年、1930—1950年和1990—2000年存在较多的更替个体(表2)。大多数臭冷杉个体出现在1930年之后。鱼鳞云杉也具有类似的年龄结构,种群建立的高峰时期发生在1820—1830年、1860—1870年、1930—1950年和1990—2000年,其更新主要发生在近80 a。在过去180 a,生长释放的频率较高,在每一个时期都有发生(表2)。61个鱼鳞云杉和106个臭冷杉的年轮样本中,共表现出97次生长释放。生长释放比例超过15%的事件有2次,分别发生在1920—1930年和1980—1990年,与种群更新高峰的时期同步。
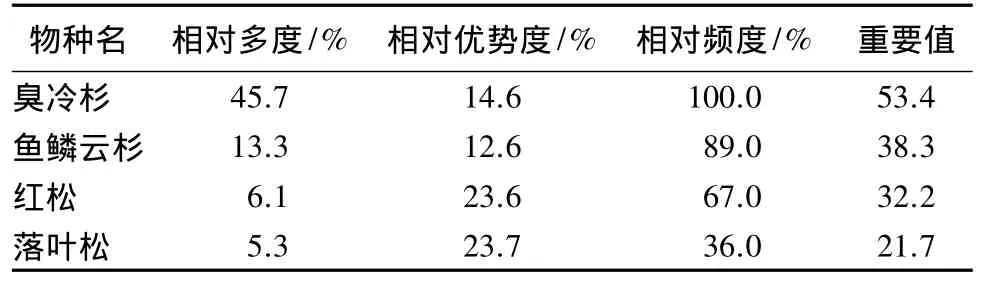
表1 长白山北坡生态交错带主要树种的重要值
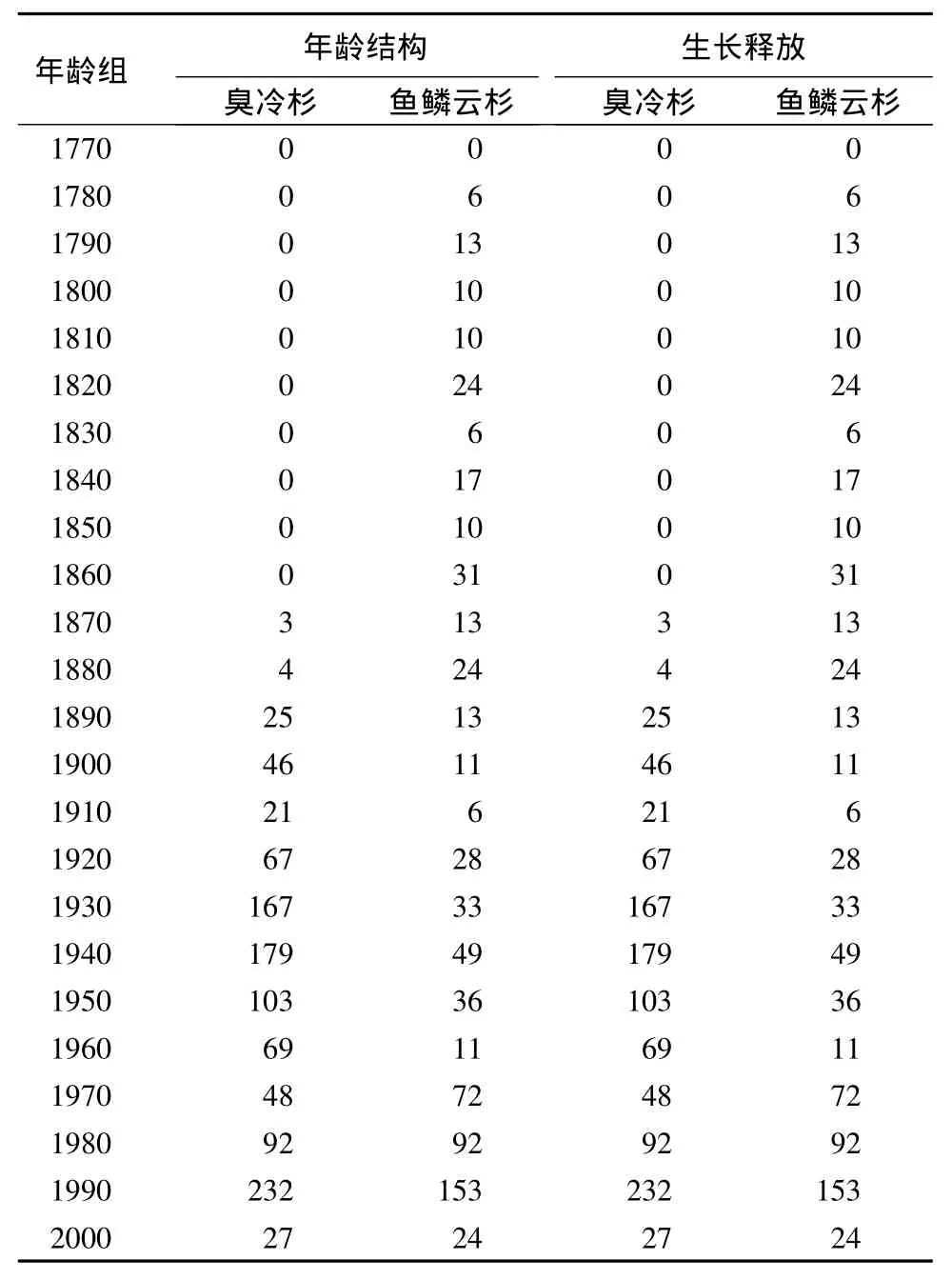
表2 长白山生态交错带臭冷杉和鱼鳞云杉的年龄结构和生长释放 株·hm-2
3.3 红松生长对气候变化的响应
在红松分布的上限(1 300 m),其径向生长和当年7月份温度存在显著相关关系,和降水没有关系,说明温度是影响红松上限分布生长的主要因子(表3)。
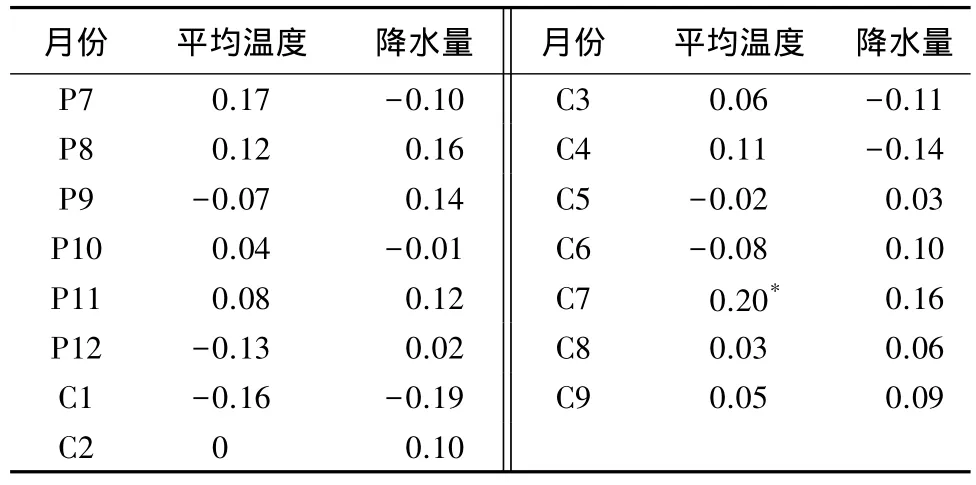
表3 长白山生态交错带红松分布上限径向生长与气候因子的相关性
4 结束语
生长释放和年龄结构分析表明,该生态交错带在过去的200 a里经历了两次尺度较大的干扰(1920—1930年和1980—1990年),其余时段属于小尺度的干扰。1920—1930年和1980—1990年生长释放的比例高于15%,而在干扰发生后的1930—1940年和1990—2000年更新出现了高峰值,对干扰事件形成了同步的呼应。这两次干扰事件发生的时期和以往研究的结果一致[10,15],引起生长释放的因素可能为风干扰。1986年长白山西坡记录到了一次飓风事件,造成了大面积的林木风倒[15],而1920—1930年的干扰事件虽然没有直接的飓风记录,但是在调查样地内未发现火烧的痕迹或者记录的其他干扰事件(病、虫害)的发生,因而,推断是由风干扰引起的。
中国的东北地区,风干扰是影响群落动态的重要因子,对物种共存起着重要的作用[16-18]。本研究表明,干扰能够促进鱼鳞云杉和臭冷杉的更新,其他研究表明,干扰也能促进该地区先锋种落叶松的更新和生长以及红松生长[17],从而使得这些树种能够在群落内自我维系以及进入林冠层,形成以落叶松、红松、鱼鳞云杉和臭冷杉为主要树种的稳定群落结构[16]。
在红松分布上限,其生长主要受夏季温度(7月份)影响,这和其他研究结果一致[8,19]。7 月份正值红松生长的旺季,较高的温度能够促进树木的生理活动,提高光合作用效率,从而有利于红松的生长。而红松在上限的生长则不受降水的限制,树木在其海拔上限的生长往往都和降水无显著关系[20]。
本研究表明,气候变暖有利于长白山交错带红松的生长,其他研究表明,温度的增加总体上有利于该交错带内鱼鳞云杉和落叶松的生长[8,21],从而直接对交错带的群落组成和结构产生影响。但是这种影响也会受到极端气候事件引起的干扰(例如,火山爆发、火灾和飓风等),尤其是气候变化间接引起的风干扰频度以及强度的变化,对长白山地区森林群落的动态起着重要作用。因此,在某个区域进行气候变化对森林群落动态研究时,应综合考虑直接和间接两个方面的作用,这在将来森林结构和组成的研究方面具有很大的应用空间。
[1] Nemani R R,Keeling C D,Hashimoto H,et al.Climate-driven increases in global terrestrial net primary production from 1982 to 1999[J].Science,2003,300:1560-1563.
[2] Qian Weihong,Zhu Yafen.Climate change in China from 1880 to 1998 and its impact on the environmental condition[J].Climatic Change,2001,50(4):419-444.
[3] He Hongshi,Hao Zhanqing,Mladenoff D J,et al.Simulating forest ecosystem response to climate warming incorporating spatial effects in north-eastern China[J].Journal of Biogeography,2005,32(12):2043-2056.
[4] Way D A,Oren R.Differential responses to changes in growth temperature between trees from different functional groups and biomes:a review and synthesis of data[J].Tree Physiology,2010,30(6):669-688.
[5] 王婷,于丹,李江风,等.树木年轮宽度与气候变化关系研究进展[J].植物生态学报,2003,27(1):23-33.
[6] 邵雪梅,吴祥定.利用树轮资料重建长白山区过去气候变化[J].第四纪研究,1997,17(1):76-85.
[7] Yu Dapao,Gu Huiyan,Wang Jiandong,et al.Relationships of climate change and tree ring of Betula ermani tree line forest in Changbai Mountain[J].Journal of Forestry Research,2005,16(3):187-192.
[8] 高露双,王晓明,赵秀海.长白山过渡带红松和鱼鳞云杉径向生长对气候因子的响应[J].植物生态学报,2011,35(1):27-34.
[9] 陈力,尹云鹤,赵东升,等.长白山不同海拔树木生长对气候变化的响应差异[J].生态学报,2014,34(6):1568-1574.
[10] 阳含熙,谢海生.长白山红松混交林干扰历史的重构研究[J].植物生态学报,1994,18(3):201-208.
[11] Fritts H C.Tree rings and climate[M].London:Academic Press,1976.
[12] Liang Eryuan,Shao Xuemei,Hu Yuxi,et al.Dendroclimatic evaluation of climate-growth relationships of Meyer spruce(Picea meyeri)on a sandy substrate in semi-arid grassland,north China[J].Trees Structure and Function,2001,15(4):230-235.
[13] Nowacki G J,Abrams M D.Radial-growth averaging criteria for reconstructing disturbance histories from presettlement-origin oaks[J].Ecological Monographs,1997,67(2):225-249.
[14] Biondi F,Waikul K.Dendroclim 2002:A C++program for statistical calibration of climate signals in tree-ring chronologies[J].Computer& Geosciences,2004,30(3):303-311.
[15] 侯向阳,韩进轩,谢海生.长白山红松林干扰节律研究[J].生态学报,2000,20(3):409-411.
[16] Liu Qijing.Structure and dynamics of the subalpine coniferous forest on Changbai Mountain,China[J].Plant Ecology,1997,132(1):97-105.
[17] Zhang Yun,Drobyshev I,Gao Lushuang,et al.Disturbance and regeneration dynamics of a mixed Korean pine dominated forest on Changbai Mountain,north-eastern China[J].Dendrochronologia,2014,32(1):21-31.
[18] 朱良军,金光泽,王晓春.典型阔叶红松林干扰历史重建及干扰形成机制[J].植物生态学报,2015,39(2):125-139.
[19] 李广起,白帆,桑卫国.长白山红松和鱼鳞云杉在分布上限的径向生长对气候变暖的不同响应[J].植物生态学报,2011,35(5):500-511.
[20] Tranquillini W.The physiological ecology of the alpine timberline[M].New York:.Springer-Verlag,1979.
[21] 于大炮,王顺忠,唐立娜,等.长白山北坡落叶松年表及其与气候变化的关系[J].应用生态学报,2005,16(1):14-20.
