姚江水系中华鳖不同组织3种同工酶的表达差异
2015-08-20郑小青梅依霞洪炆飞汪财生唐伟钱国英李彩燕
郑小青 梅依霞 洪炆飞 汪财生 唐伟 钱国英 李彩燕

摘要:采用聚丙烯酰胺垂直梯度凝胶电泳法,对姚江水系中华鳖(Pelodiscus sinensis)的6种组织(心脏、肝脏、脾脏、肾脏、肌肉、肺)中的乳酸脱氢酶(LDH)、酯酶(EST)、谷氨酸脱氢酶(GDH) 3种同工酶进行了初步研究,并与日本品系中华鳖的同工酶谱图进行了对比。结果表明:2个品系中华鳖的同工酶具有不同程度的组织特异性,LDH组织差异极为明显;姚江水系中华鳖相同组织LDH整体表达量与日本中华鳖类似,EST-1表达量高于日本中华鳖,GDH整体表达量低于日本中华鳖,肌肉中GDH-3表达量高于日本中华鳖。本研究得到了姚江群体中华鳖的标志性酶谱,并明确了其与日本品系中华鳖酶谱的差异和特殊性。
关键词:中华鳖;同工酶;表达;组织特异性;乳酸脱氢酶;酯酶;谷氨酸脱氢酶
中图分类号: S917.4 文献标志码: A 文章编号:1002-1302(2015)07-0244-03
中华鳖(Pelodiscus sinensis)又称水鱼、团鱼、元鱼、甲鱼等,是我国重要的特种经济动物和名特优水产养殖品种之一。近10余年来,在养鳖业蓬勃发展的同时,人们对野生资源的不合理开发加剧,中华鳖的生存形势持续恶化,原种资源不断减少。过度的食用消费、栖息地的破坏、化学污染等因素导致我国中华鳖种质资源遭受严重破坏,种群面临大规模锐减、濒临灭绝的境地,尤其是一些特殊地域种群的原种地没有引起足够的重视。如主要分布在宁波姚江流域的姚江水系中华鳖原种已处于濒危状态,野生种质资源稀缺。该群体中华鳖抗病力强、适应性好、肉质鲜美、胶质浓郁,深受消费者喜爱。姚江水系中华鳖除了具有中华鳖的基本固有特征外,主要特征为腹部有4处明显花斑,与其他品系中华鳖差异非常明显。目前,关于中华鳖同工酶的研究大多集中于黄河群体、太湖群体和台湾群体[1-3],而姚江水系中华鳖同工酶表达差异的研究尚未见报道。为了有效保护中华鳖的种质资源,保持地方种群优良的种质性状,应用聚丙烯酰胺垂直梯度凝胶电泳法对姚江水系中华鳖6种不同组织中的乳酸脱氢酶(LDH)、酯酶(EST)、谷氨酸脱氢酶(GDH) 3种同工酶进行了研究,并与日本品系中华鳖进行比较,以期在蛋白质水平上建立其种质资源的遗传标记,为该地方品系中华鳖的种质鉴定、遗传分析及良种选育提供科学依据。
1 材料与方法
1.1 试验材料
试验材料为三龄雄性中华鳖,其中姚江水系中华鳖取自浙江省余姚市明凤淡水养殖场,日本品系中华鳖取自绍兴中亚工贸园有限公司。
1.2 样品制备
活鳖运回实验室后,迅速杀死取其心脏、肝脏、 肾脏、脾脏、肺和腿部肌肉,称质量后加入Tris-HCl缓冲液(pH值 75)进行匀浆,再离心20 min,取其上清液與甘油按1 ∶2混合后,直接电泳或于-80 ℃保存备用。
1.3 电泳
采用聚丙烯酰胺垂直梯度凝胶电泳法[4-7],操作步骤参考区又君等报道的方法[5]进行,电泳过程使用夹心式垂直板电泳槽(Mini-PROTEAN Tetra),取10 μL样品加样。分离胶浓度为7.5%(pH值8.8),浓缩胶浓度为4.5%(pH值6.8),电极缓冲系统为 Tris-Gly(pH值8.3),在160 V 恒压、冰浴中电泳2 h(除EST研究的电泳时间为1.5 h外)。
电泳后的凝胶染色参照何忠效等[6]和郭尧军[7]的操作方法,并加以修改(表1)。在电泳结束前,提前配制好染色液,其中EST染色液需过滤后使用。电泳后的凝胶置于染色液中直至显色,LDH、EST检测凝胶室温下约染色20 min,GDH检测凝胶置于37 ℃染色约4 h。经蒸馏水清洗染色后的显色凝胶置于光亮背景下拍照记录。
2 结果与分析
应用于种群生化遗传分析的同工酶谱,除酶带应清晰外,更重要的是反映出编码该酶的几个基因座位、存在几个等位基因、是几聚体的酶等特征性信息[8]。2个不同品系中华鳖的6种组织中3种同工酶活性强弱结果见表2。从表2中可以看出,2个品系中华鳖的心脏、肝脏、脾脏、肾脏中LDH、EST 2种同工酶均明显表达且含量较高,肺与肌肉中的LDH、EST、GDH 同工酶有所表达但相比于其他组织的同工酶含量则较少。其中姚江水系中华鳖的6种组织中属肝脏组织中同工酶的整体表达量最高,而日本品系中华鳖的6种组织中属肾脏组织中同工酶的整体表达量最高。
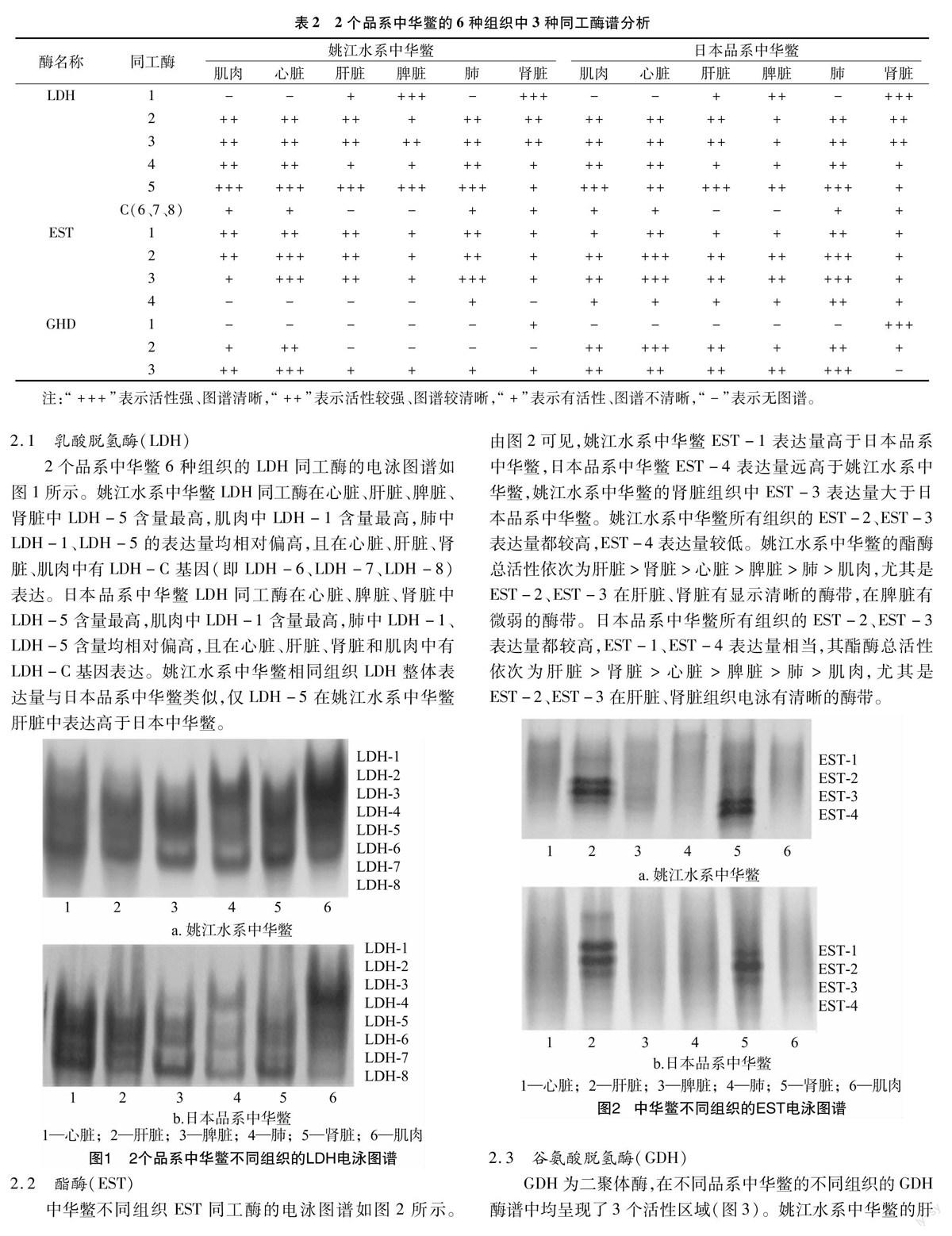
2.1 乳酸脱氢酶(LDH)
2个品系中华鳖6种组织的LDH同工酶的电泳图谱如图1所示。姚江水系中华鳖LDH同工酶在心脏、肝脏、脾脏、肾脏中LDH-5含量最高,肌肉中LDH-1含量最高,肺中LDH-1、LDH-5的表达量均相对偏高,且在心脏、肝脏、肾脏、肌肉中有LDH-C基因(即LDH-6、LDH-7、LDH-8)表达。日本品系中华鳖LDH同工酶在心脏、脾脏、肾脏中LDH-5含量最高,肌肉中LDH-1含量最高,肺中LDH-1、LDH-5含量均相对偏高,且在心脏、肝脏、肾脏和肌肉中有LDH-C基因表达。姚江水系中华鳖相同组织LDH整体表达量与日本品系中华鳖类似,仅LDH-5在姚江水系中华鳖肝脏中表达高于日本中华鳖。
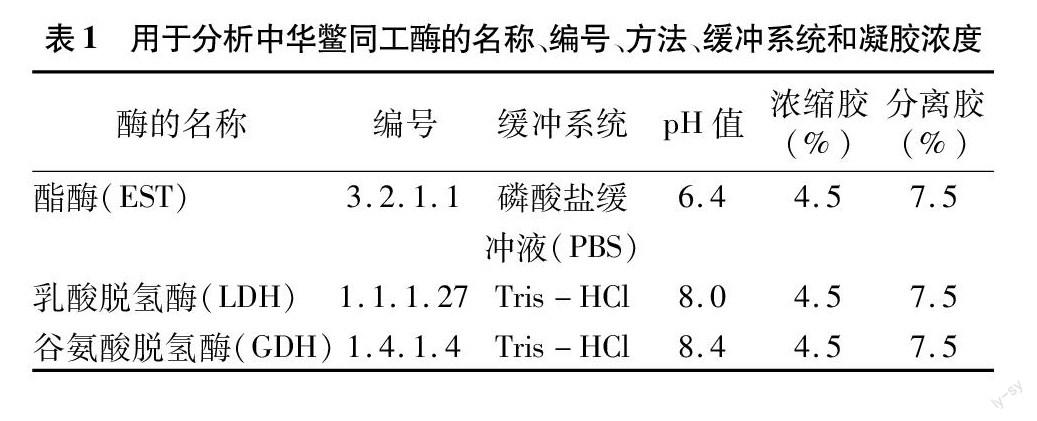
2.2 酯酶(EST)
中华鳖不同组织EST同工酶的电泳图谱如图2所示。由图2可见,姚江水系中华鳖EST-1表达量高于日本品系中华鳖,日本品系中华鳖EST-4表达量远高于姚江水系中华鳖,姚江水系中华鳖的肾脏组织中EST-3表达量大于日本品系中华鳖。姚江水系中华鳖所有组织的EST-2、EST-3表达量都较高,EST-4表达量较低。姚江水系中华鳖的酯酶总活性依次为肝脏>肾脏>心脏>脾脏>肺>肌肉,尤其是EST-2、EST-3在肝脏、肾脏有显示清晰的酶带,在脾脏有微弱的酶带。日本品系中华鳖所有组织的EST-2、EST-3表达量都较高,EST-1、EST-4表达量相当,其酯酶总活性依次为肝脏>肾脏>心脏>脾脏>肺>肌肉,尤其是 EST-2、EST-3在肝脏、肾脏组织电泳有清晰的酶带。
2.3 谷氨酸脱氢酶(GDH)
GDH为二聚体酶,在不同品系中华鳖的不同组织的GDH酶谱中均呈现了3个活性区域(图3)。姚江水系中华鳖的肝脏、心脏、肾脏、肌肉组织中GDH表达量较高,其他组织表达量低,且在肌肉中有GDH-1、GDH-3的表达。日本品系中华鳖在肝脏、脾脏、肾脏、肌肉中GDH表达量均较高,其中肝脏中的GDH表达量最高;GDH-1在肌肉中大量表达而在其他组织几乎不表达;肌肉中未检测到GDH-2、GDH-3,而其他5种组织中都有所表达。日本鳖GDH表达量整体上高于姚江水系中华鳖,但肝脏和心脏组织中2个品系中华鳖的GDH表达量相近,肌肉组织中姚江水系中华鳖GDH-3的表达量高于日本中华鳖。
3 结论与讨论
已有学者采用同工酶电泳分析了不同群体中华鳖的组织表达特异性[1-3,9-11]。如杨弘等采用垂直淀粉胶电泳对中华鳖脑、眼、心脏、肌肉、肝脏、肾脏和卵巢7种组织中的6种同工酶系统(LDH、ADH、IDH、ME、GAPDH、EST)进行研究,构建了中华鳖同工酶酶谱,揭示了LDH在中华鳖进化上的保守性[2]。姚雁鸿等运用垂直聚丙烯酰胺凝胶电泳技术对中华鳖和台湾鳖的肝脏、肾脏、眼、肌肉等几种组织中的LDH、EST同工酶进行比较研究,结果发现在中华鳖组织中有组织特异性、无多态性的LDH同工酶在台湾鳖的组织中却既有组织特异性又有多态性[3]。
3.1 乳酸脱氢酶
乳酸脱氢酶(LDH)是糖代谢途径中一种重要的氧化还原酶,存在于机体所有组织细胞的胞质内,能催化丙酮酸与乳酸之间的相互转化,是研究最广泛的一种同工酶[1-3,9-12]。已经证明LDH同工酶系统具有明显的种族及组织特异性[9,12]。心肌是含氧量丰富的组织,细胞以有氧代谢为主,催化乳酸生产丙酮酸的LDH-5占优势。骨骼肌是相对缺氧的组织,催化丙酮酸还原生产乳酸的LDH-1占优势,使骨骼肌因激烈运动造成的相对缺氧条件下仍能得以酵解获得能量[13]。
中华鳖作为水陆两栖的动物,能适应较大的含氧量波动,既能适应水环境,也能适应短时间的陆上生活,故其肺部LDH-1、LDH-5都有较多表达。肝脏、肾脏、脾脏中LDH-5占優势的原因与心肌中LDH-5占优势的原因类似,且不同组织其他的酶带均有所差异,推测LDH可作为姚江水系中华鳖组织特异性鉴定的同工酶。
日本品系中华鳖和姚江品系中华鳖的心脏、肝脏、肾脏、肌肉组织都有LDH-C基因不同程度的表达,其表达量与相应组织在中华鳖体内的活动度有一定的线性关系,推测LDH-C基因的表达可增加产能。此外,在姚江水系中华鳖的肝脏中LDH-5占优势,而在日本品系中华鳖中却没有出现该现象,这可能与日本中华鳖个体的脂肪含量较高有关(数据另文发表)。以上2个推测还有待进一步研究验证。
3.2 酯酶
酯酶(EST)是催化酯类化合物水解进入中间代谢的重要水解酶,在细胞内执行最基本的生理功能[10],其结构多为单体,由1个亚基组成[4]。EST还能水解大量正常存在的非生理性酯类化合物,可能与机体的解毒功能有关。本研究发现,姚江水系中华鳖EST整体表达量低于日本鳖,猜测原因可能与中华鳖的生存环境有关。
酯酶并非生理性修饰酶,故可以EST酶谱作为遗传指标[14-15]。从酶谱整体的角度看,姚江水系中华鳖和日本品系中华鳖的EST酶谱均呈现组织特异性。 酯酶EST-4在日本品系中华鳖中表达而在姚江水系中华鳖中基本不表达,而姚江水系中华鳖肾脏中的酯酶EST-3表达量高于日本品系,且姚江水系中华鳖肝脏和肾脏中EST条带数目多、活性强,具有种属特异性,均可作为姚江水系中华鳖的种属鉴定指标。
3.3 谷氨酸脱氢酶
谷氨酸脱氢酶(GDH)是动物体内参与谷氨酸新陈代谢的关键酶之一[12-15],它以NAD、NADP或者同时利用两者作为辅酶,参与到谷氨酸的合成与分解中,但在大多数生物体内反应是向着合成谷氨酸的方向进行的[16]。目前水产动物同工酶GDH的研究少见报道。本研究发现,GDH-3在日本品系中华鳖肌肉中不表达,而在姚江水系中表达,表明GDH-3的表达可能会导致某种生化途径改变,从而影响生物的代谢途径。由于谷氨酸是重要的鲜味氨基酸,故猜测GDH-3的高表达可能是姚江水系中华鳖与日本中华鳖相比味道更为鲜美的原因,但其相关性有待于进一步确认。GHD是氨基酸代谢中的重要酶类,尤其在非必需氨基酸的合成和转化中起着重要作用[17];因此GDH-3的表达是否直接影响氨基酸含量也有待于进一步研究。
参考文献:
[1]赵金良,蔡完其,李思发.中华鳖同工酶的组织特异性研究[J]. 上海水产大学学报,1998,7(4):311-316.
[2]杨 弘,夏德全,吴婷婷,等. 中华鳖同工酶研究[J]. 中国水产科学,1997,4(4):55-59.
[3]姚雁鸿,余来宁,方耀林,等. 中华鳖和台湾鳖的同工酶初步比较[J]. 淡水渔业,2002,32(6):44-45.
[4]朱蓝菲.鱼类同工酶和蛋白质的聚丙烯酰胺梯度凝胶电泳法[J]. 水生生物学报,1992,16(2):183-185,197.
[5]区又君,齐旭东,李加儿.波纹唇鱼不同组织5种同工酶表达的差异[J]. 南方水产,2009,5(2):51-55.
[6]何忠效,张树政. 电泳[M]. 北京:科学出版社,1990:287-298.
[7]郭尧君. 蛋白质电泳实验技术[M]. 北京:科学出版社,1999:70-80.
[8]王可玲,尤 锋,徐 成,等. 5种海水鱼同工酶表达的组织特性及其电泳的初步分析[J]. 海洋与湖沼,1996,27(6):626-631,682.
[9]李永通,向应海,杨业勤. 中国大鲵及鳖不同组织LDH同工酶的比较研究[J]. 动物学杂志,1992,27(1):28-31.
[10]王金星,赵小凡,廖正根. 休眼与非休眠期中华鳖乳酸脱氢酶同工酶的比较分析[J]. 海洋湖沼通报,1995(2):43-47.
[11]叶玉珍,王小虎,吴清江. 孵化温度对中华鳖4种同工酶和肌肉蛋白基因表达的影响[J]. 华中农业大学学报,2004,23(4):385-388.
[12]肖调义,陈清华,陈开健,等. 四种黄颡鱼乳酸脱氢酶同工酶电泳的研究[J]. 上海水产大学学报,2004,13(1):72-74.
[13]艾晓杰,杨丽娥,程美蓉. 白香猪消化器官乳酸脱氢酶同工酶的比较研究[J]. 四川畜牧兽医,2005(8):23-24.
[14]王启瑞. 缘蜻科部分种类酯酶同工酶的分子系统学研究[D]. 西安:陕西师范大学,2001:10-16.
[15]朱新平,林礼堂,夏仕玲. 鲮鱼、麦瑞加拉鲮鱼及露斯塔野鲮酯酶同工酶的电泳研究[J]. 淡水渔业,1992(5):30-31.
[16]王亚平,姜宇海. 谷氨酸脱氢酶在黄疸鉴别诊断中的应用[J]. 南京军医学院学报,2003,25(4):294-295.
[17]曹志华,高贵琴,张巧玲. 鲤鱼肠道及肠内微生物谷氨酸脱氢酶同工酶及活性的初步研究[J]. 淡水渔业,2001,31(5):52-53.
