云南地区种子植物属多样性的纬度格局与区系起源的关系
2015-08-08丰帮艳冯建孟
丰帮艳, 冯建孟
(大理学院 农学与生物科学学院,云南 大理 671003)
0 引言
生物多样性沿地理梯度的变化一直是生物多样性研究的重要课题之一[1].其中,有关生物多样性纬度分布格局的研究受到广泛关注.虽然,物种多样性与纬度呈负相关关系的结论被很多人所接受[2-6];但也有学者认为,多样性在纬度梯度上呈现出单峰格局,即物种丰富度的峰值出现在中纬度地区[6-8].同时,为了解释物种多样性的纬度格局,人们也提出了若干个理论或假设,如“起源中心”假说[9-10]、“生态时间”假说[11-12]、“历史干扰”假说[12-15]以及“中域效应”假说[16]等.上述理论或假设的提出,为更深、更全面地理解物种多样性纬度格局的成因起了不可忽视的推动作用.但总体而言,有关物种多样性纬度格局的表现形式及其解释还存在着一定的争议[17-22],相关研究有待进一步深入.
植物区系是植物与自然地质历史长期适应、自身迁移和进化的结果.在一定程度上,植物区系可以理解为植物从其起源地扩散到其他地区的迁移历史、进化过程及其地理分布[23].因此,植物区系的起源及其扩散历史可能会在一定程度上影响植物多样性的纬度格局.例如,“起源中心”假说认为植物多样性起源于热带地区并向高纬度扩散;热带地区的植物类群在短时间内有更高的物种形成率和更低的灭绝率,导致其类群更年轻、更有生态优势,故在热带低纬度地区植物类群多样性更为丰富[10-12].同时,由于受生态位“保守主义”等因素的影响,使热带地区作为一个多样性泵[9],维持着相对较高的生物类群多样性.“历史干扰”假说认为高纬度地区多样性较低,可能是因为热带植物类群受到历史性冰川的影响,而没有足够的时间向纬度更高、更寒冷的气候区传播和扩散[12,15].所以,植物的区系地理起源可能是影响植物多样性纬度格局的重要因素之一.因此,开展植物区系起源与物种多样性纬度格局之间关系的研究有助于更好地验证或发展生物多样性的“起源中心”假说 和“历史干扰”假说,从而更好地理解物种多样性的纬度分布格局的形成机制.云南地区拥有复杂的地形、较低的纬度和特殊的生物地理位置,分布着丰富、多样的植物区系类型[23-24],是探讨植物区系起源与物种多样性纬度格局之间关系的理想区域.因此,笔者从区系地理起源的角度探讨云南地区植物多样性纬度分布格局的成因.有研究表明,更高的分类阶元更容易被发现和记录,故有助于减少调查强度因地区差异所导致的偏差[25-26].所以, 本文主要探讨云南地区属水平上的植物区系地理起源与植物多样性纬度分布格局之间的关系.
1 数据来源及方法
1.1 数据来源
研究区域概况见文献[23-24,27-29].
本研究中的云南地区种子植物的源数据主要来自于《云南植物志》(1-16卷)[29].在此基础上,也利用了云南自然保护区考察报告中的植物名录及其他地方性植物志进行补充和修正[30-37].同时,通过相关文献获得了云南地区县域行政区划矢量图及其面积和经、纬度中值信息[24].
1.2 研究方法
根据源数据中记载的每个物种的基本信息,对各物种的科、属、种的拉丁名、分类地位及分布信息等进行分析整理,并建立了以县(市)域为单位的植物物种分布数据库.以吴征镒(1991)中国种子植物属的区系类型[38]为参照,分别对研究区域内123个县(市)域所包含的种子植物属进行区系类型标识.分布区类型包括T1(世界分布)、T2(泛热带分布)、T3(热带亚洲和热带南美间断分布)、T4(旧世界热带分布)、T5(热带亚洲至热带大洋洲分布)、T6(热带亚洲至热带非洲分布)、T7(热带亚洲分布)、T8(北温带分布)、T9(东北和北美间断分布)、T10(旧世界温带分布)、T11(温带亚洲分布)、T12(地中海区、西亚至中亚分布)、T13(中亚分布)、T14(东亚分布)和T15(中国特有分布)等共15个类型.其中,T2~T15称为非世界分布区系类型,T2~T7为热带区系成分,T8~T14为温带区系成分.以此为基础,利用MS Excel分别统计了各县(市)域中各属区系类型所包含的属数.同时,还分别计算了各县(市)域内世界分布成分和各非世界分布区系类型属所占的比重以及温、热带区系成分属所占比重之和.利用各县(市)域的纬度中值和SPSS 17.0TM统计软件包分别对温、热带区系成分的比重之和进行纬度格局分析.
统计热带属所占比重多于温带属所占比重的县(市)域数量,并计算其占123个县(市)域的比重;以123个县(市)域的温、热带植物区系比重为数据集,利用SPSS17.0统计软件包对热、温带属的比重进行了Mann-Whitney检验,以此比较在纬度梯度上温、热带属区系比重的大小.同时,笔者分别以各县(市)域温、热带区系属数和总属数为分子,以各县(市)域面积的自然对数为分母,利用公式(1)[25]分别获得了各县(市)域的温、热带区系属密度和总属密度, 以此为基础,利用SPSS17.0TM统计软件包探讨上述属密度在纬度梯度上的分布格局.
D=S/lnA,
(1)
其中:D为属密度,A为县(市)域面积,S为丰富度(即属的数量).
2 结果
2.1 属水平上温、热带区系成分的相对组成
研究表明,研究区域内种子植物由2 338个属组成.其中,热带属1 345个,占属总数的57.5%;温带属749个,所占比重为32.0%.热带属数量明显多于温带属的数量.从空间分布来看,Mann-Whitney检验结果表明,属于热带区系成分的属数总体上明显多于温带区系成分的属数(P<0.001);同时,研究结果也表明,研究区域内123个县 (市) 域中有114个县 (市) 域的热带属所占比重大于温带属所占比重,占总县 (市) 域总数的92.6%.因此,无论从总体组成来看,还是从123个县 (市) 域的具体分布来看,属于热带区系成分的属数均明显多于属于温带区系成分的属数,即热带区系成分在研究区域内占主导地位.
2.2 温、热带属比重的纬度分布格局
从图1可看出,随着纬度的升高,热带区系成分属的比重之和呈现出显著下降的趋势(R2=0.77,P<0.001).由图2可知,温带区系成分属的比重之和随纬度的升高呈显著增加的趋势(R2=0.78,P<0.001).总体而言,在由南向北的纬度梯度上,热带属所占比重逐渐递减、温带属所占比重则呈相反的趋势.

图1 热带区系比重之和的纬度变化趋势Fig.1 Latitudinal patterns of the sum of the proportions of tropical elements

图2 温带区系比重之和的纬度变化趋势Fig.2 Latitudinal patterns of the sum of the proportions of temperate elements
2.3 属密度的纬度分布格局
由图3可知,热带属密度随纬度升高呈下降的趋势(R2=0.44,P<0.001).从图4可以发现,温带属密度随纬度增加呈增加趋势(R2=0.25,P<0.001);图5则表明总属密度与纬度之间存在着明显的负相关关系(R2=0.09,P<0.001).总体上,在由南向北的纬度梯度上,热带属多样性呈减少趋势、温带属多样性呈增加趋势,而总体多样性则呈递减趋势.

图3 热带属密度的纬度变化趋势Fig.3 Latitudinal patterns of tropical genus density
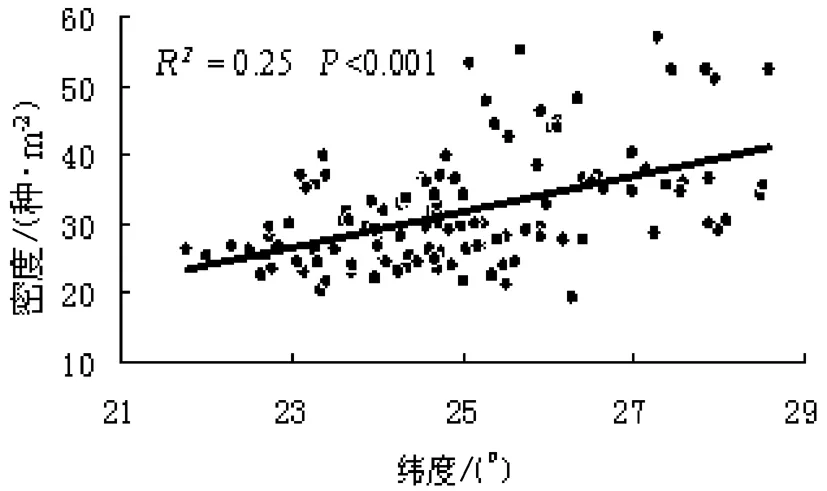
图4 温带属密度的纬度变化趋势Fig.4 Latitudinal patterns of temperate genus density

图5 所有属密度的纬度变化趋势Fig.5 Latitudinal patterns of total genus density
3 讨论
研究结果表明,无论在比重方面,还是在密度方面,热带区系属与纬度呈明显的负相关关系,而温带区系属与纬度呈正相关关系;同时,在地理 (纬度) 梯度上和总体组成上,热带属所占比重总体上明显多于温带属.另外,值得注意的是,总体属多样性(密度)也呈递减趋势.这可能意味着,由于热带区系属的主导地位, 而且其在纬度上呈递减趋势,所以在一定程度上导致了总体属密度在纬度梯度上的递减格局.
本研究利用区系地理理论,可以在一定程度上同时解释纬度梯度上植物多样性的递减格局和递增格局.起源于热带地区,喜温的植物类群的多样性随纬度上升呈递减趋势,这可能与该区系类群起源于低纬度地区,其北上扩散的进程需要漫长的时间和长期的低温适应有关,这导致了该区系类型的植物多样性在低纬度地区相对饱和,多样性较为丰富;而中高纬度则相对不饱和,多样性相对较低.对起源于温带地区、耐寒的植物类群而言,其从中高纬度地区南下扩散的进程,同样需要漫长的时间和长期的对温暖环境的适应,故该区系类型植物多样性在中高纬度地区是相对饱和的,多样性较为丰富;但在低纬度地区则相对不饱和,多样性相对较低.植物总体属多样性的纬度分布格局则可能是温、热带植物属多样性纬度格局的叠加,并由于热带属多样性的纬度递减格局而呈递减趋势.因此,我们认为植物区系地理理论可能是解释生物多样性纬度格局的重要理论或假说之一,但不可否认其相关研究还有待进一步深入.
“起源中心”假说和“历史干扰”假说均被用于解释生物类群多样性的纬度递减格局,但不能同时解释生物类群多样性在纬度梯度上的递减格局和递增格局.本研究既考虑了生物类群的热带或低纬度起源,兼容了生物类群的温带起源或中高纬度起源,同时也结合了生物类群的扩散理论,故可以有效地解释云南地区植物类群在纬度梯度上的递减和递增格局.因此,本研究在一定程度上丰富和发展了“起源中心”假说和“历史干扰”假说.
Rapoport法则认为随着纬度的升高,气候变化幅度增加,生物为适应这一梯度变化,其生态位在高纬度地区相对较宽,但是在低纬度地区,物种的生态位相对较窄,故产生“拯救效应”,导致低纬度地区物种多样性丰富,而高纬度地区物种多样性相对贫乏[21].但本研究则表明,随着纬度的升高,起源于热带地区的植物属多样性呈递减趋势,而起源于温带地区的植物属多样性则呈递增趋势.这可能是因为,对于喜温的热带起源的植物类群而言,其起源地的气候变化幅度相对较小,对纬度梯度上气候因子的变化幅度相对较为敏感,故随着纬度升高,热带起源的植物生态位宽度呈增加趋势,并因为“拯救效应”导致其多样性呈递减趋势.而对于温带区系起源的植物类群而言,其起源地的气候变化幅度相对较大,而且本身的生态位也相对较宽,故可能对纬度梯度上气候变化幅度的变化较为适应或较为不敏感,故未出现明显的“拯救效应”,进而,未出现该植物类群的多样性随纬度升高而降低的趋势.因此,从纬度较高的起源地向低纬度地区扩散时,其多样性呈递减趋势.也意味着,Rapoport法则可能更适用于起源于热带地区的物种多样性纬度格局的解释.
4 结论
结果表明,热带植物类群主要集中在低纬度地区,而温带植物类群则主要集中在中高纬度地区.从保护对象的植物类群组成角度来看,低纬度地区植物多样性的主要保护对象可能是热带区系植物类群,而对于纬度相对较高的地区而言,其主要保护对象则应是温带区系植物类群.但如果从保护对象的生物类群丰富度的角度来看,我们发现研究区域的低纬度地区拥有相对更为丰富的种子植物类群,故可能是云南地区植物多样性保护较为重要的区域.
