家蝇精氨酸激酶基因克隆及其在害虫防治上的应用
2015-07-24于雪曹新茹高一夫唐婷柳峰松
于雪,曹新茹,高一夫,唐婷,柳峰松
(河北大学 生命科学学院,河北 保定 071002)
精氨酸激酶(arginine kinase,ATP:N-phosphotransferase EC2.7.3.3)广泛存在于无脊椎动物如昆虫、甲壳动物和软体动物体内,是无脊椎动物能量代谢最重要的酶类之一.当动物体内的ATP充足时,可以将能量储存在磷酸精氨酸的高能磷酸键中;当动物消耗大量的ATP时,精氨酸激酶催化磷酸精氨酸分解生成ATP.精氨酸激酶在无脊椎动物生命活动中发挥重要作用,是无脊椎动物特有的磷酸原激酶,该酶及其能量代谢途径与哺乳动物不同,因此,精氨酸激酶可以作为防控害虫的一个有效而安全的靶点.
RNA 干扰(RNA interference,RNAi)是指外源或内源双链RNA(double strand RNA,dsRNA)介导的特异性降解靶向mRNA,导致转录后水平的基因沉默(post-transcriptional gene silencing,PTGS)[1].RNA干扰具有抑制转座子活动、抗病毒入侵、调控基因表达等作用,是重要的监控机制.Guo等[2]利用反义RNA手段探索线虫(Caenorhabditis elegans)par1基因的功能时发现了RNAi现象.Fire等[3]详尽地解释了这种现象是由于制备反义RNA 时混入了微量的dsRNA 引起的,认为dsRNA 是引发RNA 沉默的关键因子,并把这种现象称为RNAi.目前,RNAi技术已经在多种昆虫上得到应用,dsRNA 进入昆虫体内的方法主要有投喂、浸泡、注射、转基因和病毒介导等,这些方法各有特点,投喂法最具前景[4-8].
家蝇(Musca domestica)俗称苍蝇,其繁殖能力强,生长速度快,世界各地广泛分布,与人类的关系密切.家蝇携带多种病原菌,并能将病原菌传给人类或牲畜,危害性较大[9].本研究以家蝇精氨酸激酶基因为靶点,通过投喂长dsRNA 来抑制家蝇体内精氨酸激酶的表达水平,大大影响了家蝇的存活率,以此揭示精氨酸激酶在家蝇体内的生物学功能,并为害虫防治提供了一种新思路.
1 材料与方法
1.1 试虫与试剂
1.1.1 供试昆虫
家蝇(Musca domestica)由本实验室保种和饲养,饲养温度为25 ℃,幼虫饵料成分:麸皮55g,灭活酵母粉3g和150mL水.成蝇饲养成分:水、红糖和奶粉.
1.1.2 菌株及质粒
大肠杆菌(Escherichia coli)DH5α菌株、HT115菌株以及L4440质粒.
1.2 主要试剂和试剂盒
RNAiso Plus,pMD18-T Vector,限制性内切酶,T4DNA 连接酶购自大连宝生物公司,M-MLV 反转录酶,SYBR Green,Taq DNA 聚合酶均为北京全式金生物技术有限公司产品,引物合成自苏州金唯智生物技术有限公司.其他试剂均为国产或进口分析纯.
1.3 总RNA 的提取及cDNA的合成
依照宝生物RNAiso Plus使用说明书,按步骤提取家蝇3龄幼虫总RNA.核酸定量仪测定总RNA 纯度和浓度,经琼脂糖凝胶电泳检验完整性后,定总量为2μg的总RNA 为RT-PCR 模板,用通用引物Oligo(dT)反转录合成cDNA.
1.4 家蝇MdAK 基因的克隆
从本实验室测得的家蝇转录组数据库中搜索到一段长3 140bp的EST 序列(CL 939.Contig 1),通过Blastn程序在GenBank数据库中对该序列进行同源性分析,发现与昆虫精氨酸激酶基因相似性较高.根据此EST 序列用Primer premier 5.0设计正反向引物MdAK-F和MdAK-R(表1),以家蝇cDNA 为模板,进行PCR 扩增,PCR 反应条件为:94℃预变性5min;94℃变性30s,55℃退火40s,72℃延伸3min,30个循环;72 ℃延伸10min.PCR 产物经琼脂糖凝胶电泳分离、切胶回收后,连接pMD18-T 载体,转化至感受态细胞DH5α中,经菌落PCR 筛选阳性克隆后测序.

表1 PCR 引物Tab.1 Primers for PCR
1.5 MdAK 基因序列的生物信息学分析
应用Bioedit软件搜索MdAKcDNA 序列开放阅读框(open reading frame,ORF),并将核酸序列翻译成氨基酸序列.在ExPASy:ProtParam 程序中(http://au.expasy.org/tools/protparam.html)分析MdAK的分子质量和等电点.利用Blastp程序在NCBI网站上进行同源序列查找,对获得的昆虫精氨酸激酶的氨基酸序列利用Clustal软件进行多重序列比对.
1.6 L4440载体的构建和dsRNA的诱导
根据家蝇MdAK 的cDNA 序列,利用软件Primer设计出上下游引物RNAi-F和RNAi-R,引物5'末端分别加入BamHI和XhoI限制性酶切位点,将扩增片段与L4440载体经过双酶切回收后,连接获得重组质粒L4440-MdAK,将其转化至DH5α感受态细胞中,经过菌落PCR 筛选阳性克隆菌,提取小量质粒,重复双酶切,检验后测序验证.将构建好的干扰载体转化至HT115宿主,通过菌落PCR 筛选阳性转化子.阳性转化子接种100mL含有氨苄青霉素(终质量浓度60μg/mL)和四环素(终质量浓度50μg/mL)的LB(Lysogeny Broth)新鲜液体培养基,37 ℃摇床200r/min培养至OD600=0.6~0.8时加入2mol/L IPTG(isopropyl-β-D-thiogalactoside)至终浓度为0.4mmol/L,37 ℃,200r/min诱导3h.同时,诱导表达含空载L4440质粒的菌作为空白对照.提取菌体RNA 进行电泳检测,观察dsRNA 的表达情况.
1.7 RNAi实验
分别将经IPTG 诱导的含有L4440-MdAK 重组质粒和L4440 空载质粒的HT115 菌液稀释至OD600=0.1,用以配置家蝇幼虫饵料,80mL菌悬液加入60g麸皮,均匀搅拌,分别装在玻璃瓶中.向每个玻璃瓶中投放200只刚刚孵化的家蝇1龄幼虫,25 ℃培养箱中培养.之后每天观察幼虫数量变化并统计存活率,最后观察幼虫化蛹和羽化情况.每组实验重复3次.
1.8 MdAK 基因敲低效果分析
投喂表达MdAK dsRNA 的菌液后,于6,24,36h后各取6只家蝇幼虫,提取总RNA 并反转录,以βactin基因作为内参,进行实时荧光定量PCR 检测MdAK mRNA 表达量变化.设计目的基因定量引物MdAK-RT-F/R;内参基因定量引物β-actin-F/R.实时荧光定量PCR 程序:95 ℃2 min;95 ℃20s,60 ℃30s,30个循环.实时荧光定量结果用相对定量2-ΔΔCt法进行分析,用t检验来比较实验组和对照组的差异,P>0.05则为无显著差异,0.01<P<0.05为显著差异,P<0.01为极显著差异.
2 实验结果
2.1 MdAK 序列分析
通过RT-PCR 克隆得到一段全长为2 007bp的MdAKcDNA 序列,包含1 071bp的开放阅读框,可编码356个氨基酸残基(图1).利用ExPASy:ProtParam 程序分析,MdAK 蛋白理论分子质量为40.0ku,理论等电点为5.91.利用Blastp程序在NCBI网站上进行同源搜索,发现家蝇精氨酸激酶与黑腹果蝇(Drosophila melanogaster)的精氨酸激酶氨基酸序列相似性较高(identity=91%),于是将此基因命名为MdAK(Musca domesticaarginine kinase).精氨酸激酶氨基酸序列多重比对结果表明:不同物种的精氨酸激酶氨基酸中第271~277位的序列CPTNLGT,62 位天冬氨酸(Asp)和193 位精氨酸(Arg)保守性较高(图2).CPTNLGT 是精氨酸激酶的活性中心序列,271位的半胱氨酸(Cys)是活性中心必需氨基酸.
2.2 RNAi处理后精氨酸激酶基因的表达变化
将PCR 扩增得到的MdAK 基因片段克隆到L4440载体,由于克隆位点两侧各含有1个T7启动子,经IPTG 诱导后,插入片段被双向转录并在大肠杆菌HT115中形成dsRNA.提取细菌总RNA 进行电泳检测,发现L4440-MdAK 转化菌株较L4440 转化菌株明显多出1 条亮带(图3),证明MdAK 双链RNA 诱导成功.
利用投喂法对家蝇1龄幼虫MdAK 基因进行了RNAi实验,将能表达MdAKdsRNA 的工程菌和空载对照菌分别添加到幼虫饵料中进行投喂.为了检测MdAK 基因的敲低效果,对进行RNAi处理6,24,36h后的家蝇幼虫体内MdAK 基因进行定量研究,结果发现RNAi处理组MdAK 基因表达被有效抑制,且在处理后36h的敲低程度达到80%以上(图4).

图2 昆虫精氨酸激酶同源序列比对分析Fig.2 Multiple alignment of arginine kinases from insects
2.3 RNAi处理后家蝇存活率统计
RNAi处理家蝇幼虫1,2,3,4,5,6d后,统计家蝇的数量(图5).结果显示,实验组家蝇幼虫在RNAi处理后第4天起开始大幅死亡,第6天家蝇的死亡率达到80%以上,而对照组的死亡率较低,不到10%.同时发现RNAi处理后实验组家蝇化蛹率和羽化率也低于对照组,即使幼虫存活至羽化为成蝇,生长速度也要比对照组慢很多,个体明显要小于对照组,幼虫尾部常常发黑,活动能力很差.
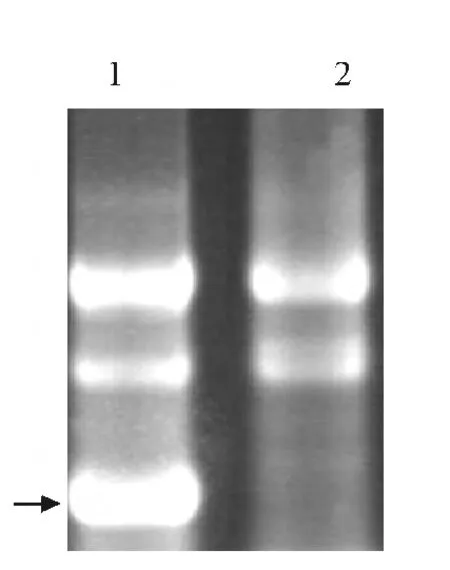
图3 HT115总RNA电泳结果Fig.3 Total RNA electrophoretogram of HT115

图4 RNAi干扰对精氨酸激酶基因mRNA表达的影响Fig.4 Relative expression level of MdAK mRNA at different time points after RNAi treatment

图5 RNAi处理后家蝇幼虫的数量变化Fig.5 Changes in mortality rate of housefly after RNAi treatment
3 讨论
精氨酸激酶是无脊椎动物体内主要的磷酸原激酶,其在无脊椎动物能量代谢上的重要作用已得到广泛证实.本研究利用RT-PCR 技术获得了长度为2 007bp的家蝇精氨酸激酶cDNA,编码356个氨基酸.不同昆虫的精氨酸激酶的氨基酸序列虽有不同,但有较高的同源性,相似性都在70%以上,这表明精氨酸激酶在进化上高度保守,同时也提示其功能的重要性.
精氨酸激酶是无脊椎动物体内不可缺少的、调节能量代谢的酶,且精氨酸激酶仅存在于无脊椎动物体内,哺乳动物体内肌酸激酶参与的代谢途径与无脊椎动物体内精氨酸激酶参与的途径不同.所以,精氨酸激酶可作为害虫防治的潜在靶点.家蝇分布广,繁殖能力强,免疫力强,生长周期短,适于实验室研究.本研究,以家蝇作为实验对象,通过投喂表达dsRNA 的工程菌来敲低家蝇体内精氨酸激酶基因的表达水平.大肠杆菌HT115为RNaseⅢ缺失突变菌株,有助于dsRNA 的稳定表达.基因定量结果显示,投喂表达MdAK dsRNA 的大肠杆菌后,精氨酸激酶mRNA 表达水平明显低于对照组,同时发现家蝇的生长发育受到了很大的影响,且幼虫存活率显著降低.
dsRNA 分子较单链RNA 分子相对稳定,喷洒到植物叶片上的dsRNA 能存留几天[10-11].这样使得RNAi技术具有潜力成为害虫防治的有力工具.本研究证实dsRNA 可有效干扰家蝇体内精氨酸激酶基因的表达,并能有效杀死家蝇,且该方法仅对节肢动物等含有精氨酸激酶的生物有作用,不会影响其他生物的正常生长和发育,具有较高的安全性.化学农药的使用给环境和人类健康带来了不可估量的影响,寻求新的害虫防治手段具有重要的意义.随着昆虫系统生物学和基因工程等学科的快速发展,针对害虫的特有基因设计特异性药物已经成为了害虫生物防治的重要研究方向,具有广阔的应用前景.
[1] 王子见 吴秀萍 邓洪宽,等.RNA 干扰技术在动物寄生线虫研究中的局限性[J].中国寄生虫学与寄生虫病杂志,2008,26(6):476-479.WANG Zijian,WU Xiuping,DENG Hongkuan,et al.Limitations of the application of RNA interference in the study of animal parasitic nematodes[J].Chinese Journal of Parasitology and Parasitic Diseases,2008,26(6):476-479.
[2] GUO Su,KEMPHUES K J.Par-1,agene required for establishing polarity in C.elegans embryos,encodes a putative Ser/Thr kinase that is asymmetrically distributed[J].Cell,1995,81(4):611-620.
[3] FIRE A,XU Siqun,MONTGOMERY M K,et al.Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans[J].Nature,1998,391(6669):806-811.
[4] BAUM J A,BOGAERT T,CLINTON W,et al.Control of coleopteran insect pests through RNA interference[J].Nat Biotechnol,2007,25(11):1322-1326.
[5] DZITOYEVA S,DIMITRIJEVIC N,MANEV H.Intra-abdominal injection of double-stranded RNA into anesthetized adult Drosophila triggers RNA interference in the central nervous system[J].Mol Psychiatry,2001,6(6):665-670.
[6] EATON B A,FETTER R D,DAVIS G W.Dynactin is necessary for synapse stabilization[J].Neuron,2002,34(5):729-741.
[7] TURNER C T,DAVY M W,MACDIARMID R M,et al.RNA interference in the light brown apple moth,Epiphyas postvittana(Walker)induced by double-stranded RNA feeding[J].Insect Mol Biol,2006,15(3):383-391.
[8] UHLIROVA M,FOY B D,BEATY B J,et al.Use of Sindbis virus-mediated RNA interference to demonstrate a conserved role of Broad-Complex in insect metamorphosis[J].Proc Natl Acad Sci U S A,2003,100(26):15607-15612.
[9] SCOTT J G,LIU N,KRISTENSEN M,et al.A case for sequencing the genome of Musca domestica(Diptera:Muscidae)[J].J Med Entomol,2009,46(2):175-182.
[10] TENLLADO F,BARAJAS D,VARGAS M,et al.Transient expression of homologous hairpin RNA causes interference with plant virus infection and is overcome by a virus encoded suppressor of gene silencing[J].Mol Plant Microbe Interact,2003,16(2):149-158.
[11] TENLLADO F,MARTINEZ-GARCIA B,VARGAS M,et al.Crude extracts of bacterially expressed dsRNA can be used to protect plants against virus infections[J].BMC Biotechnol,2003,3:3.
