大黄鱼在感染刺激隐核虫后MHCIIB基因的表达
2015-07-13陈健张东玲叶坤等
陈健 张东玲 叶坤等
摘要[目的] 为进一步研究大黄鱼抗刺激隐核虫的免疫防御奠定基础,也为海水鱼类养殖的免疫预防提供参考。[方法] 用孵化2 h内的刺激隐核虫感染大黄鱼,并运用Real-time PCR检测MHC IIB基因在各组织(鳃、皮肤、脾和头肾)中表达量的变化。[结果] MHC IIB基因在鳃、皮肤和脾脏中的表达水平呈现先上调后逐渐下降的趋势,在感染6和12 h后表达量显著升高(P<0.05);在头肾中,MHC IIB基因在感染后6 和12 h的表达量显著下调(P<0.05),在感染2 d后显著上升(P<0.05)。[结论] 大黄鱼MHC IIB基因在刺激隐核虫的免疫应答过程中可能发挥重要作用。
关键词大黄鱼;MHC IIB;刺激隐核虫
中图分类号S965.322文献标识码A文章编号0517-6611(2015)07-143-04
主要组织相容性复合体(Major histocompatibility complex,MHC)是目前已知存在于所有脊椎动物体内与机体免疫机能密切相关的基因群[1-2]。最常见的MHC主要由2类分子组成:MHC I和MHC II类。MHC I类分子负责将内源性抗原呈递给CD8+ T细胞,MHC II类分子则将外源性抗原呈递给CD4+细胞[3-4]。MHC II类分子由α链和β链组成,其中MHC IIB基因编码β链区域,是研究最为广泛的一类MHC II分子[5-6]。近年来,已有多种鱼类的MHC IIB基因被克隆和鉴定,如虹鳟(Oncorhynchus mykiss)[7-9]、真鲷(Pagrus major)[10]、鮸鱼(Miichthys miiuy)[11]、半滑舌鳎(Cynoglossus semilaevis)[12]、大菱鲆(Turbot)[13]、草鱼(Grass carp)[14]、建锂[15](Cyprinus carpio var.jian)、鲤鱼(Carp)[16-17]和斑马鱼(Zebrafish)[18-21]等,这些研究表明了MHC IIB基因的多态性和基因拷贝数目的差异性。MHC IIB基因或蛋白的组织分布和表达情况也有研究,在真鲷中MHC IIB基因在注射鳗弧菌后在肝、脾、头肾和肠中的表达5~72 h明显下降,96 h恢复[22];在大菱鲆中MHC IIB基因在注射鳗弧菌后在肝和头肾中的表达24~48 h呈下降趋势,在脾中24~72 h显著下降,96 h后在3个组织中的表达均上升[23]。前期的研究表明 MHC IIB基因参与由病毒和细菌所引起的鱼类免疫应答,但对其是否参与鱼类抗寄生虫的免疫应答少有报道。
大黄鱼(Larimichthys crocea)是我国主要养殖海水鱼类之一。近年来,由于养殖密度过大和养殖环境恶化等原因,导致疾病频频暴发,严重威胁产业的发展。寄生虫病害以刺激隐核虫病引起的“白点病”最为严重,每年造成巨大的经济损失。对于小水体养殖的刺激隐核虫病,目前主要采取化学药物治疗[24]。对于网箱等开放养殖环境,一些物理方法和化学方法均起到一定效果,但它们都或多或少存在一些缺陷,如对鱼有毒性、污染环境、效果不确定以及不适用于大水体等[25-26]。因此,有必要寻找一种切实可行、有效的防治方法。由于免疫预防的安全性、低成本和环保等优势,免疫防治刺激隐核虫病目前被认为是一种相对有效的方法。笔者对感染刺激隐核虫后MHC IIB基因在各组织(鳃、皮肤、脾、头肾)中表达量的变化进行研究,探讨MHC IIB基因在宿主对刺激隐核虫免疫应答中发挥的作用,从分子角度初步揭示大黄鱼抗寄生虫的分子机制,为进一步系统研究大黄鱼抗刺激隐核虫的免疫防御奠定基础,同时也为海水鱼类养殖的免疫预防提供参考。
1材料与方法
1.1 材料
1.1.1
试验材料。大黄鱼:健康的大黄鱼采自福建宁德养殖海区,体重(130 ±15) g;
刺激隐核虫:以大黄鱼为宿主,宁德海水试验场传代培养获得幼虫。
1.1.2
主要试剂。Trizol、RNA保存液(RNAfixer)、逆转录试剂盒(Promega GoscriptTM Reverse Transcription System)、荧光定量试剂盒(SuperReal PreMix Plus)等。
1.2方法
1.2.1
刺激隐核虫感染大黄鱼。分为试验组和对照组,每组60条鱼。试验前将每组大黄鱼在室内水泥池内驯养10 d,盐度25‰~26‰。温度25 ℃。室内驯养后,试验组用刚孵化2 h内的刺激隐核虫感染大黄鱼,感染剂量为25 000幼虫/尾[27]。由于刺激隐核虫滋养体感染鱼体后脱落,在池底形成包囊,3 d后孵化出幼虫。为了避免重复感染,感染刺激隐核虫的大黄鱼3 d后移入新池。对照组除不给予刺激隐核虫感染外,其他试验方法均与试验组一致。试验组和对照组分别在首次感染后6 h、12 h、1 d、2 d、3 d和5 d采集大黄鱼皮肤、鳃、脾脏和头肾(每个时间点每组取5尾鱼)。
1.2.2
大黄鱼组织总RNA的提取及cDNA链的合成。
采用Trizol分别提取大黄鱼鳃、皮肤、脾脏和头肾4个组织的总RNA,DNase I处理残留 DNA 后,根据Promega GoscriptTM Reverse Transcription System说明书合成第一链cDNA。反转录PCR反应体系:RNA样品4 μg,Oligo(dT)15 Primer 1 μl,Random Primer 1 μl,Nuclease-Free Water补至10 μl体系,70 ℃变性5 min,冰上放置5 min;再加入Nuclease-Free Water 1.5 μl,Goscript 5×Reaction Buffer 4 μl,MgCl2 2 μl,PCR Nucleotide Mix 1 μl,Recombinant RNasin Ribonuclease Inhibitor 0.5 μl,Goscript Reverse Transcriptase 1 μl,共10 μl体系。混匀后,25 ℃ 5 min,42 ℃ 1 h,70 ℃ 15 min。合成后于-20 ℃保存备用。
1.2.3
大黄鱼MHC IIB组织表达分析。根据大黄鱼MHC IIB序列,设计特异引物(表1),以β-actin作为内参基因,通过RT-PCR分析大黄鱼MHC II基因的表达模式。利用 LightCycler 480 荧光定量仪(Roche Sweden)进行实时荧光定量 PCR 反应。实时荧光定量PCR反应体系:2×SuperReal PreMix Plus 10.0 μl,引物F/R(10 μmol/L)0.6 μl,cDNA模板 8.4 μl,50×ROX Reference Dye 0.4 μl。实时荧光定量PCR反应程序:95 ℃ 15 min;95 ℃ 10 s,60 ℃ 20 s,72 ℃ 32 s,40个循环。利用Fold=2-ΔΔCT计算基因的相对表达量,然后用试验组数值除以对照组数值得到试验组相对于对照组的表达量,使用 SPSS 15.0 统计软件的 Duncans 检验法分析各组数据差异的显著性。
表1所用引物序列
引物序列(5′ -3′) 目的
MHCII B QFCTGGCAGACGCTGATTGGTmRNA表达
MHCII B QRTCTCTCAGACTCAGGCAGGGAC
β-actin-FTTATGAAGGCTATGCCCTGCCmRNA表达
β-actin-RTGAAGGAGTAGCCACGCTCTGT
2 结果与分析
2.1荧光定量引物的特异性检测
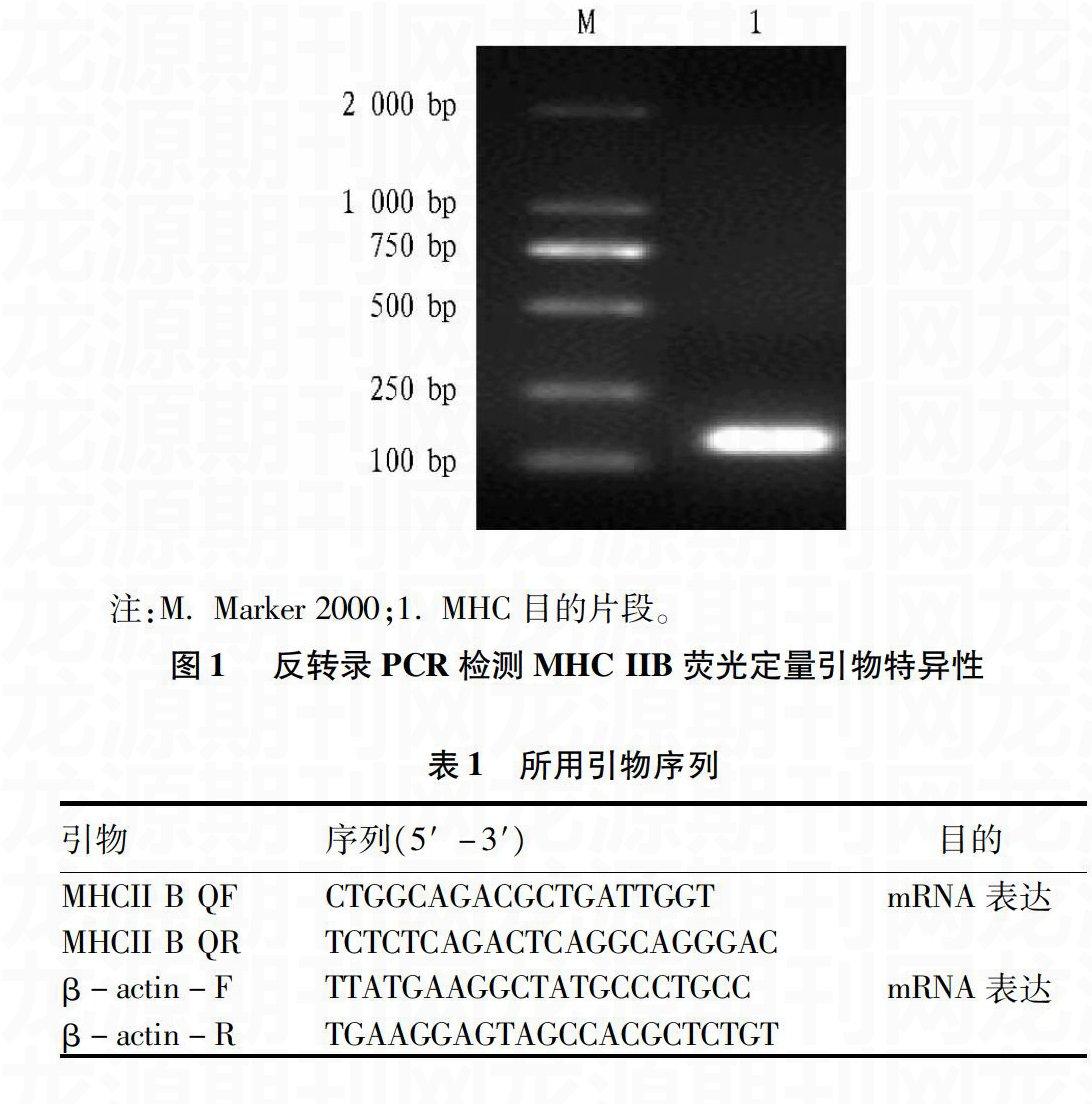
为了检验引物的特异性,反转录PCR扩增MHC IIB基因片段,PCR产物用1% 琼脂糖凝胶电泳进行检测,结果发现其与预期目的基因片段(150 bp)大小一致(图1),测序结果证实序列正确,说明设计合成的引物合格。
注:M. Marker 2000;1. MHC目的片段。
图1 反转录PCR检测MHC IIB荧光定量引物特异性
试验组与对照组共8组不同检测体系,以各自cDNA为模板,经过qRT-PCR检测后得到扩增曲线(图2a)和熔解曲线(图2b)。分析发现,所得曲线均为单一峰,无非特异性扩增产物和引物二聚体产生,说明设计合成的引物特异性良好。
2.2刺激隐核虫感染大黄鱼诱导MHC IIB的表达分析
大黄鱼受刺激隐核虫感染后,通过qRT-PCR检测大黄鱼MHC IIB基因在4种组织(鳃、皮肤、脾和头肾)中的表达水平。从图3可以看出,在鳃组织中,受刺激隐核虫感染后,试验组大黄鱼MHC IIB表达量在6和12 h显著上升(P<0.05),分别为对照组的3.09倍和3.19倍。皮肤在感染12 h后MHC IIB基因的表达量显著上升(P<0.05),其表达量为对照组的6.12倍;在感染3 d后MHC IIB基因的表达量显著下调(P<0.05),其表达量仅为对照组的10.1%。
在脾脏,试验组大黄鱼MHC IIB基因的表达量在6和12 h显著上升(P<0.05),分别为对照组的3.82倍和4.89倍,此后逐步恢复到对照组水平。头肾在受刺激隐核虫感染后,表达量开始下降,在6和12 h MHC IIB基因的表达量显著下调(P<0.05),分别为对照组的61.9%和66.0%,感染2 d后MHC IIB基因的表达量显著上升(P<0.05),其表达量为对照组的5.21倍。
3 讨论
大黄鱼鳃和皮肤是刺激隐核虫感染的主要器官,也是鱼类免疫防御的第一道屏障,而脾脏和头肾是大黄鱼重要的免疫器官[28],因此选择这4个部位来研究大黄鱼在受刺激隐核虫感染后MHC IIB基因的表达变化。在皮肤和鳃,在感染初期(6 h或12 h)MHC IIB的表达量显著升高,12 h表达量差异达到最大值。这与斜带石斑鱼(Epinephelus coioides)被刺激隐核虫感染后, CD8 β、IL-1 β、C型凝集素和transferrin mRNA在皮肤和鳃表达水平上升的结果一致[29]。皮肤和鳃是鱼体粘膜组织,它们也是与水体病原直接接触的部位,与水体寄生虫最先接触,受感染后的炎症部位也最为明显。皮肤表皮上有粘液细胞和囊状细胞的分布,鳃组织的细胞中分布有大量淋巴细胞、中性粒细胞和上皮细胞等[30]。粘膜组织具有参与机体免疫应答的细胞基础,能够抵御寄生虫感染和引起相关免疫应答反应。目前,许多学者认为粘膜免疫系统可以不依赖系统免疫而独立完成抗原识别、结合和呈递,因此局部的免疫应答对抵御病原的入侵起着重要的作用[5]。
粘膜组织(皮肤和鳃)不仅仅
是物理屏障,其局部的免疫应答对病原体的抵御也相当重
要。MHC IIB是生物体抵抗病原体侵染的免疫基因,在鱼类的粘膜免疫抗原呈递中发挥着重要作用。
注:a.扩增曲线;b.熔解曲线。
图2 MHC IIB基因的扩增曲线和熔解曲线
注:a.鳃;b.皮肤;c.脾;d.头肾。*表示显著上升;§表示显著下降。
图3刺激隐核虫感染大黄鱼后MHC IIB基因在鳃、皮肤、脾脏和肾脏的表达分析
刺激隐核虫感染大黄鱼鳃和皮肤后,引起局部免疫应答,抗原向脾脏和头肾转移[28]。脾脏接收抗原刺激后,MHC IIB的表达量也显著上升,产生免疫应答以抵御病原入侵。脾脏是鱼类红细胞与中性粒细胞产生、贮存和成熟的主要场所,头肾可以产生红细胞和B淋巴细胞等,相当于哺乳动物的骨髓。头肾受感染后MHC IIB的表达量显著下降,直至感染2 d后表达量才显著回升。Xu TJ等研究发现半滑舌鳎(Cynoglossus semilaevis)注射鳗弧菌后。MHC IIB基因在脾组织中的表达量变化不大,在肝组织中24 h后的表达量呈下降趋势,但在96 h后表达量达到最高[11-12],此趋势与头肾中MHC IIB的表达趋势相一致。头肾位于头胸腔下侧,紧贴脊椎,应答抗原刺激较缓慢,到受病原感染后期才产生强烈免疫应答反应。这表明MHC IIB基因的表达与组织所含有的淋巴细胞有较密切的关系,从而表现免疫器官有较强的基因表达,而在鳃、皮肤等部位的表达主要是因为这些器官是鱼类最易接触病原体的部位。据此推断, MHC IIB基因在鱼类刺激隐核虫免疫应答中发挥着重要作用。
[19] ONO H,KLEIN D,VINCEK V,et al. Major histocompatibility complex class Ⅱgenes of zebra-fish[J]. Proc Natl Acad Sci USA,1992,89(24):11886-11890.
[20] S?LTMANN H,MAYER W,FIGUEROA F,et al. Organization of Mhc class ⅡB genes in the zebra-fish (Brachydanio rerio)[J]. Genomics,1994,23:1-14.
[21] S?LTMANN H,FIGUEROA F,O'HUIGIN C,et al. Zebrafish Mhc class Ⅱ alpha chain-encoding genes: polymorphism,expression,and function[J]. Immuno- genetics,1993,38(6):408-420.
[22] CHEN SL,ZHANG YX,XU MY,et al. Molecular polymorphism and expression analysis of MHC class Ⅱ B gene from red sea bream (Chrysophrys ma- jor)[J]. Dev Comp Immunol,2006,30(4):407-418.
[23] ZHANG YX,CHEN SL. Molecular identification,polymorphism,and expression analysis of mjor histocompatibility complex class ⅡA and B genes of
turbot (Scophthalmus maximus)[J]. Marine Biotechnology(NY),2006,8:611-623.
[24] 邱军强,刘玮,杨先乐,等. 刺激隐核虫免疫学研究进展[J]. 中国病原生物学志,2011,6(10):787-797.
[25] 徐润林,白庆笙,李贵生,等. 刺激隐核虫感染幼虫的超微结构研究[J]. 中山大学学报论丛,1995(1):142-145.
[26] 刘振勇,林小金,谢友佺,等. 大黄鱼刺激隐核虫病继发细菌感染致死原因的研究[J]. 福建水产,2010(1):47-48.
[27] 钟正苹,但学明,李安兴,等. 斜带石斑鱼CD4基因克隆和组织表达分析[J]. 四川动物,2008,30(6):875-881.
[28] 刘云,姜国良,姜 明,等. 牙鲆鳃淋巴样组织内免疫相关细胞的超微结构[J].青岛海洋大学学报,2001,31(6):872-876.
[29] LI YW,DAN XM,ZHANG TW,et al. Immune-related genes expression profile in orange- spotted grouper during exposure to Cryptocaryon irritans[J].
Parasite Immunol,2011,33: 679- 687.
[30] FAST MD,SIMS DE,BURKA JF,et al. Skin morphology and humoral non-specific defence parameters of mucus and plasma in rainbow trout,coho and
Atlantic salmon[J]. Comp Biochem Physiol,2002,132: 645-657.
