梅与山桃远缘杂交亲和性初步研究
2015-07-05王翠梅董然然王一兰陈瑞丹
王翠梅,董然然,王一兰,袁 涛,陈瑞丹
(花卉种质创新与分子育种北京市重点实验室,国家花卉工程技术研究中心,城乡生态环境北京实验室,北京林业大学 园林学院,北京100083)
梅(Prunus mume)是中国传统名花,同时也是中国第一种获得国际品种登录权的花卉,距今已有7 000多年的应用史和3 000多年的栽培史[1]。梅自古就以寒中开放的特性受到中国人民的喜爱,但梅花的抗寒能力并没有人们想象的那么强。梅的露地栽植范围大致以黄河为北界,以珠江为南界[2],因此中国北方人民无缘欣赏到梅花。中国梅花工作者自20世纪中期开始培育抗寒梅花品种,通过引种驯化、实生选种的方式株选出‘北京玉蝶’(P.mume‘Bei jingYudie’)和‘北京 小’梅(P.mume‘Beijing Xiaomei’)等真梅系抗寒梅花品种[3],可以在北京露地越冬。20世纪80年代开始的远缘杂交育种育成了‘燕杏’梅(P.mume‘Yanxing’)、‘花蝴蝶’(P.mume‘Hua Hudie’)、‘山桃白’(P.mume‘Shantaobai’)梅等杏梅系品种[4-5],并成功引种‘淡丰后’(P.mume‘Dan Fenghou’)等13 个品种。陈瑞丹以培育芳香抗寒梅花品种为目标,通过杂交育种成功培育出芳香抗寒的梅花新品种‘香瑞白’梅(P.mume‘Xiang Ruibai’)[6]。但 这 些 品 种 虽 抗 寒 性强,但开花较晚,在北京及以北地区自然花期在3月下旬至4月初,不能体现梅早花的特性,降低了其观赏性。因此,如何提早抗寒梅花的花期是目前育种工作者迫切需要解决的问题。
种间远缘杂交是植物种质创新和品种改良的重要途径之一[7],远缘杂交后代有明显的杂种优势,通过远缘杂交获得抗逆性强、观赏价值高的植物新品种是育种家追求的目标,‘燕杏’梅、‘花蝴蝶’梅、‘山桃白’梅等都是通过远缘杂交获得的,抗寒性极强。但不同物种间存在一定的生殖隔离,远缘杂交常表现为杂交不亲和性和杂种不育性,难以获得杂种后代[8]。为提早抗寒梅花的花期,使北国人民也能感受到梅花的韵味,本研究开展了梅与山桃、‘白花’山桃的杂交育种工作。选择自然结实率高且抗寒(能在北京地区露地栽植)的真梅品种‘江梅’、杏梅品种‘淡丰后’为母本。山桃(P.davidiana)和‘白花’山桃(P.davidiana‘Alba’)抗寒性强,是北京地区花期最早的梅花同属植物,且前人已获得山桃和‘小绿萼’梅(P.mume‘Xiao Lv,e’)的杂交种‘山桃白’梅[5],因此选择山桃和‘白花’山桃作为本试验的父本。对杂交结实率、授粉后花粉在柱头的萌发以及花粉管的生长过程进行观察,探讨各杂交组合的亲和性,并对未成熟胚进行培养,为梅花的远缘杂交育种工作提供基础。
1 材料和方法
1.1 试验材料
杂交试验于2014年3月下旬至4月上旬在青岛梅园进行,荧光观察和胚培养试验在北京林业大学实验室进行。杂交母本树为青岛梅园生长健壮的‘江梅’、‘淡丰后’梅,父本为北京林业大学校园内多年生且生长健壮的山桃和‘白花’山桃,杂交组合为‘江梅’×山桃、‘江梅’ב白花山’桃,‘淡丰后’×山桃、‘淡丰后’ב白花’山桃。授粉在3月底~4月上旬,上午9:30~11:00,天气晴好,无风或微风,温度10 ℃~20 ℃。
1.2 授粉方法
1.2.1 花粉的采集和贮藏 3月初山桃、‘白花’山桃开花前,采集气球期花蕾,在室内通风条件下自然阴干散粉后,收集花粉置于装有变色硅胶的干燥管中,-20 ℃条件下冰冻保存。
1.2.2 花粉生活力的测定 授粉前采用悬滴培养法测定花粉的活力。培养基为5%蔗糖+10μg/mL硼酸,24 ℃下培养6h。测得山桃花粉发芽率为55%、‘白花’山桃为60%,均可用于授粉。
1.2.3 杂交授粉方法 3月底~4月上旬梅花初开时,先对大蕾期母本用连被去雄法去雄,即用大拇指和食指将萼片、花瓣和雄蕊一次性撕掉的去雄方法,同时授以父本花粉并套袋挂牌,每朵花重复授粉2次。
1.3 花粉管生长行为观察
授粉后4、8、12、24、36、48、72、96、120、144、168 h取样,每组合随机选取授粉后的花柱20 个,用FAA 固定液(甲醛∶冰醋酸∶70%酒精=1∶1∶18,V/V)固定24h以上,4 ℃冰箱保存。
花粉在柱头上萌发和花粉管行为观察:经FAA固定液固定的雌蕊,先用蒸馏水冲洗3次,然后放入8mol/L NaOH 溶液中软化4h,蒸馏水冲洗干净后,放入含0.1mol/L K3PO4的0.1%的水溶性苯胺蓝溶液中染色1h。然后将花柱放于载玻片上,盖玻片轻轻压平。在Olympus荧光显微镜下观察记录花粉管生长情况。
1.4 杂交结实率比较及杂种幼胚拯救
授粉1 周后,拆除套袋,用纱网裹住授粉的大枝,防止果实掉落。之后每隔15d 统计1 次座果率,直至果实成熟。
2014年6月采收授粉后60d左右的果实,将梅果肉洗净,砸开种核,取出种仁,置于流水下冲洗30 min,在超净工作台上将种仁先后用70%酒精和1%次氯酸钠溶液消毒30s和25min,每次消毒后均用无菌水冲洗3~5次,去除内种皮,取带1/4子叶的幼胚 接 种 于①2/3WPM +0.5 mg/L IBA+2.0 mg/L 6-BA+0.5g/L 活性炭+40g/L 蔗糖+7 g/L琼脂和②WPM+0.5mg/L IBA+2.0mg/L 6-BA+0.5g/L 活性炭+40g/L 蔗糖+7g/L 琼脂,pH 5.8~6.0的培养基中,置于2 ℃~4 ℃的冰箱中低温黑暗处理90d,然后转入光照组培室[温度(23+2)℃,湿度60%左右,光照强度1 500~2 000 lx,光照14h/d]中培养,根据培养情况适时打开炼苗,炼苗后移栽至花盆中(7cm×7cm),移栽基质为珍珠岩∶进口泥炭土∶河沙=1∶1∶1(V/V),基质使用前用高温灭菌锅120 ℃灭菌2h。移栽后置于人工气候箱(白天25℃,晚上20℃,光照16h/d,湿度60%,光照80%)培养,观察记录杂交种子萌发情况。
结实率=(获得果实数/授粉花数)×100%
幼胚萌发率=(萌发的胚数/接种数)×100%
败育率=(败育的胚数/接种数)×100%
2 结果与分析
2.1 梅与山桃、‘白花’山桃种间杂交结实性
由表1可以看出,各杂交组合结实率都很低,在‘江梅’ב白花’山桃的杂交组合中,未能获得杂交果实,表现为种间远缘杂交不亲和性。‘江梅’和山桃杂交结实率只有0.7%,杂交亲和性很低。结实率较高的是以‘淡丰后’为母本的两个组合,分别为3.1%和7.4%。授粉后35d左右出现严重的落果现象,落果率高达90%,落果现象可能是由胚发育不正常造成的,且收获的果实大多不饱满,这种未成熟的果实即使沙藏后播种,也不能萌发。作者于2013年1月播种沙藏后的杂交种子,均未萌发,砸开种核发现里面的种仁已腐烂。分析原因认为,梅果实发育快、时间短,果实达到成熟时,胚还尚未成熟,尤其是远缘杂交由于亲和力的关系,胚后期发育不良,萌芽力很低。因此,杂交种子采取播种的方法在本试验中不能使用,必须采用更有效的试验方法进行胚拯救。
2.2 梅与山桃、‘白花’山桃授粉后花粉管行为观察
授粉后,山桃和‘白花’山桃的花粉粒在梅柱头上4~8h均能萌发,但花粉粒的萌发情况和花粉管的生长有很大差异。‘江梅’×山桃,4h 后观察到少量山桃花粉粒能够在‘江梅’柱头萌发,8h 后萌发量增加(图版Ⅰ,1),但12h后即观察到柱头乳突细胞出现胼胝质塞(图版Ⅰ,2),48h 后停止生长,花粉管顶端膨大、弯曲,未能进入子房(图版Ⅰ,3),不能完成授精作用;‘江梅’ב白花’山桃,‘白花’山桃的花粉粒在‘江梅’柱头上8h后仅有少量萌发,且伴随有大量的胼胝质沉积,花粉管末端发生弯曲、断裂、缠绕(图版Ⅰ,4),未能正常生长;‘淡丰后’×山桃,4h后即能大量萌发(图版Ⅰ,5),8h后观察到花粉管已开始深入花柱,48h 后到达花柱中部,但此后花粉管不再伸长(图版Ⅰ,6),未能进入子房;‘淡丰后’ב白花’山桃,4h后观察到花粉粒开始萌发(图版Ⅰ,7),12h 大量萌发,在花柱中延伸生长(图版Ⅰ,8),96h后观察到花粉管进入子房(图版Ⅰ,9)。
以‘江梅’为母本的两个杂交组合,虽然花粉粒能在梅柱头萌发,但均未观察到花粉管进入到子房,不能完成授精作用,存在授精前障碍;在以‘淡丰后’为母本的组合中,‘白花’山桃的花粉粒能够在雌蕊柱头萌发,沿花柱延伸,最后进入子房,完成授精作用,导致结实率低的原因是在果实发育中,胚败育导致大量落果。
2.3 幼胚培养萌发情况
由于2013年播种杂交种子均未萌发,因此本次试验种子处理采用胚培养的方法获得杂种苗。如表2中所示,接种数和结实数不一致的原因是杂交亲和性低、杂种存在严重败育的情况,并不是所有果实里都有饱满的胚,有的是干瘪的胚,有的是水渍状非正常的胚。远缘杂交极低的结实率以及胚败育导致最后能接种的幼胚极少。

表1 梅与山桃、‘白花’山桃种间杂交结实性比较Table 1 Fertility of interspecific crosses between P.mume and P.davidianaor P.davidiana‘Alba’
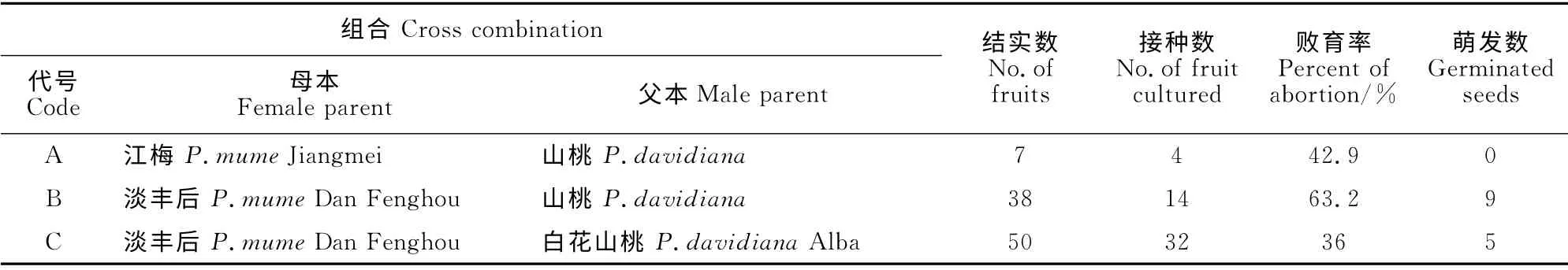
表2 梅与山桃 白花 山桃杂交幼胚培养萌发情况Table 2 Germination of immature embryo between P.mume and P.davidianaor P.davidiana‘Alba’
放置2 ℃~4 ℃的冰箱低温培养,每周观察一次萌发情况。低温培养50~55d后,杂种B、C胚开始萌动,幼芽呈现淡黄绿色,这与黑暗培养有关,未能进行光合作用,70d后胚根伸长约1cm。低温90 d后转入光照组培室培养,1~2d后,幼叶开始转绿、伸展,根伸长生长(图版Ⅰ,10、11)。5d后已长出数条二级根,此时打开三角瓶上的封口膜,炼苗5 d后移栽入盆,置于人工气候箱中生长,每天观察生长情况,视具体情况定时浇水。移栽2周后,杂种苗苗高达5~8cm(图版Ⅰ,12、13),杂种B 从叶形上已完全表现出父本的特性(图版Ⅰ,12),可以初步鉴定为‘淡丰后’和山桃的杂交后代。在人工气候箱中培养5~6月后,待植株生长健壮,且天气转暖后,换大盆移栽至露地生长。
通过未成熟胚培养获得了梅与山桃的杂种苗,说明胚培养是获得种间杂种苗的有效途径。
3 讨 论
种间远缘杂交是培育新品种的重要途径之一,但是种间杂交存在不亲和性以及杂种不育是远缘杂交育种中的两大障碍。
张启翔[9]曾观察了山桃花粉粒在‘小绿萼’梅、‘大羽’梅等真梅品种柱头上的萌发,发现山桃花粉和梅柱头之间不存在不亲和反应,都能在柱头表面萌发。本研究也观察到类似的现象,本试验4个杂交组合,花粉粒均能在雌蕊柱头上萌发,说明梅和山桃种间杂交柱头表面不存在不亲和反应,除了‘淡丰后’ב白花’山桃组合观察到花粉管进入子房,其他组合都未观察到花粉管进入子房,分析最后未能完成授精的原因,可能是和花柱中的某些物质发生识别障碍。Knox[10]通过研究杨属种间杂交提出了主动识别的概念,认为柱头识别出不亲和的花粉壁蛋白后在乳突细胞内迅速产生胼胝质,阻止花粉管继续生长进入子房。本试验通过荧光观察,发现山桃和‘白花’山桃的花粉授于‘江梅’柱头后,花粉管生长缓慢,均在花柱中产生大量胼胝质。分析原因可能是花粉粒与雌蕊柱头识别过程中,柱头存在抑制物质,阻碍花粉管的生长,诱导产生胼胝质[11],这是生物物种为了维持种间生殖隔离达到保存种性的目的[12]。不同杂交组合其结实率也存在差异,‘江梅’与‘白花’山桃杂交结果率为0,通过花粉管生长行为的观察,发现两者之间存在授精前障碍。结实率较高的为‘淡丰后’和山桃、‘白花’山桃之间的杂交,且不同亲本对杂种胚的萌发率也有影响。梁青等通过研究甜樱桃和中国樱桃的远缘杂交发现不同亲本对杂交座果率和杂种胚的萌发生长率均有显著影响[13],这可能是由遗传背景不同造成的。
远缘杂交获得的果实由于亲和力差的原因,胚后期发育不良,即使通过沙藏后播种也未萌发,砸开种核发现种仁已腐烂。本试验获得杂交果95个,种仁不饱满或干瘪率为47.4%,因此很难用常规播种方法获得杂种苗。胚培养技术的发展有效地解决了这个问题[14],通过胚拯救已获得杏和桃、李等的杂种苗[15-16]。本试验也有力地证明了胚培养技术在远缘杂交育种中的重要作用,通过胚培养共获得杂种苗13株,表明幼胚拯救是获得种间杂种苗、提高杂交成功率的重要途径。
[1] 陈俊愉.中国梅花[M].海口:中国海南出版社,1996.
[2] CHEN J Y陈俊愉.The study of Chinese Mei flowerⅠSource area and cultivation history of Mei flowerJ.Acta Horticulturae Sinica(园艺学报),1962,1(1):69-78(in Chinese).
[3] CHEN J Y(陈俊愉),ZHANG Q X(张启翔),LI ZH J(李振坚),et al.Studies on introducing and breeding hardy Mei cultivars together with extension prospects[J].Journal of Beijing Forestry University(北京林业大学学报),2003,25(特刊):1-5(in Chinese).
[4] ZHANG Q X(张启翔).Distant hybridization and cold-resistance breeding of Mei flower(Ⅰ):study on flowering and pollination habit &distant hybridization of Mei flower[J].Journal of Beijing Forestry University(北京林业大学学报),1987,9(1):69-79(in Chinese).
[5] ZHANG Q X(张启翔).Distant hybridization and cold-resistance breeding of Mei flower(Ⅱ):study on frost-resistance of distant hybrids and parents[J].Journal of Beijing Forestry University(北京林业大学学报),1988,10(4):53-69(in Chinese).
[6] 陈瑞丹,张启翔,陈俊愉.通过杂交育种培育出芳香抗寒梅花新品种[C]//张启翔.中国观赏园艺研究进展.北京:中国林业出版社,2005:1-5.
[7] DAI H J(戴华军),ZHU ZH B(朱正斌),SHEN X L(沈雪林),et al.Essences and approaches of distant hybridization in crops breeding[J].Genomics and Applied Biology(基因组学与应用生物学),2010,29(1):144-149(in Chinese).
[8] DENG Y M,TENG N J,CHEN S M,et al.Reproductive barriers in the intergeneric hybridization between Chrysanthemum grandiflorum(Ramat.)Kitam.and Ajania przewalskii Poljak.(Asteraceae)[J].Euphytica,2010,174:41-50.
[9] ZHANG Q X(张启翔).Study on interspecific pollination and parental pollen germination under 0-2℃low temperature of Prunus mume[J].Journal of Beijing Forestry University(北京林业大学学报),1986,(1):84-90(in Chinese).
[10] KNOX R B.Role of pollen-wall proteins as recognition substances in interspecific incompatibility in poplars[J].Nature,1972,237:383-390.
[11] JIA A P(贾爱平),WANG F(王 飞),YAO CH CH(姚春潮),et al.Interspecific and intraspecific cross compatibility of kiwi fruits’[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2010,3(9):1 809-1 814(in Chinese).
[12] WATANA BE K.Successful ovary culture and production of F1hybrids and androgenic haploids in Japanese Chrysanthemumspecies[J].The Journal of Heredity,1977,68:317-320.
[13] LIANG Q(梁 青),CHEN X S(陈学森),ZHANG L J(张立杰),et al.Effects of parental cultivars on cross compatibility and embryo rescue in interspecific crosses between Prunus avium and Prunus pseudocerasus[J].Journal of Fruit Science(果树学报),2006,23(3):388-391(in Chinese).
[14] DAVID W R.The use of embryo culture in fruit breeding[J].Hort Science,1900,25:393-398.
[15] ARANCHA A,DAORDEN M E,GARCIA E,et al.Significant effect of accidental pollinations on the progeny of low setting Prunusinterspecific crosses[J].Euphytica,2006,147:389-394.
[16] LIU W,CHEN X S,LIU G J,et al.Interspecific hybridization of Prunus persica with P.armeniacaand P.salicinausing embryo rescue[J].Plant Cell Tiss Organ Cult,2007,88:289-299.
Explanation of plate:
PlateⅠ Fig.1-9.Pollen germination and pollen tube growth in different cross combinations between P.mume and P.davidiana:Fig.1.P.mume‘Jiangmei’×P.davidiana,pollen germinated 4hafter pollination,×200;Fig.2.P.mume‘Jiangmei’×P.davidiana,callose accumulated 8hafter pollination,×100;Fig.3.P.mume‘Jiangmei’×P.davidiana,the end of pollen tube bended and swelled,×100;Fig.4.P.mume‘Jiangmei’×P.davidiana‘Alba’,pollen tube bended,twined and fractured 48hafter pollination,×100;Fig.5.P.mume‘Dan Fenghou’×P.davidiana,a lot of pollen germinated on the stigma,×100;Fig.6.P.mume‘Dan Fenghou’×P.davidiana,pollen tube arrived at the middle of style 48hafter pollination,×100;Fig.7.P.mume‘Dan Fenghou’×P.davidiana‘Alba’,pollen germinated 4hafter pollination,×200;Fig.8.P.mume‘Dan Fenghou’×P.davidiana‘Alba’,pollen tube prolongated in the style 12hafter pollination,×100;Fig.9.P.mume‘Dan Fenghou’×P.davidiana‘Alba’,pollen tube stretched into the ovary 96hafter pollination,×400.Fig.10-13.Embryo culture and hybrid:Fig.10.P.mume‘Dan Fenghou’×P.davidiana‘Alba’;Fig.11.P.mume‘Dan Fenghou’×P.davidiana;Fig.12.Hybrid of P.mume‘Dan Fenghou’×P.davidiana;Fig.13.Hybrid of P.mume‘Dan Fenghou’×P.davidiana‘Alba’.

图版Ⅰ 1~9.不同杂交组合花粉萌发和花粉管生长过程荧光观察:1.‘江梅’×山桃,4h花粉粒萌发,×200;2.‘江梅’×山桃,8h后胼胝质沉积,×100;3.‘江梅’×山桃,花粉管末端弯曲、膨大,×100;4.‘江梅’ב白花’山桃,48h后花粉管弯曲、缠绕、断裂,×100;5.‘淡丰后’×山桃,4h后花粉粒在柱头大量萌发,×100;6.‘淡丰后’×山桃,48h后花粉管进入花柱中部,×100;7.‘淡丰后’ב白花’山桃,4h后花粉萌发,×200;8.‘淡丰后’ב白花’山桃,12h花粉管在花柱中延伸,×100;9.‘淡丰后’ב白花’山桃,96h后进入子房,×400。10~13.胚培养及杂种苗:10.‘淡丰后’ב白花’山桃杂种胚培养情况;11.‘淡丰后’×山桃杂种胚培养情况;12.‘淡丰后’×山桃杂种苗;13.‘淡丰后’ב白花’山桃杂种苗。
