新疆野核桃自然保护区不同坡向野核桃种群生命表与波动周期
2015-06-28焦子伟任艳利杨允菲李建东
张 维,焦子伟,任艳利,杨允菲,李建东
(1伊犁师范学院化学与生物科学学院,新疆伊宁835000;2东北师范大学草地科学研究所植被生态科学教育部重点实验室,长春130024)
新疆野核桃自然保护区不同坡向野核桃种群生命表与波动周期
张 维1,2,焦子伟1,任艳利1,杨允菲2*,李建东2
(1伊犁师范学院化学与生物科学学院,新疆伊宁835000;2东北师范大学草地科学研究所植被生态科学教育部重点实验室,长春130024)
为了掌握新疆野核桃种群的波动周期、预测未来种群结构变化,在新疆野核桃自然保护区的阳坡和阴坡分别设置样地大样本调查,以径级结构代表年龄结构编制野核桃种群静态生命表,绘制其期望寿命图、年龄结构图、存活曲线和死亡率曲线,同时应用谱分析方法比较分析不同坡向生境野核桃种群的波动周期。结果表明:(1)野核桃种群不同坡向同一龄级和同一坡向不同龄级的期望寿命均存在波动,期望寿命在10龄级前为阳坡高于阴坡,10龄级后为阳坡低于阴坡。(2)阳坡和阴坡野核桃种群均为增长型年龄结构。(3)不同坡向生境野核桃种群的存活曲线均趋于Deevey-Ⅱ型,但阳坡种群各龄级的存活率均高于阴坡。(4)1龄级植株以高死亡率为代价发育为幼树;阳坡种群在其生活史中经历2个死亡高峰,最高峰在16龄级,阴坡种群经历4个死亡高峰,最高峰在9龄级。(5)谱分析表明,新疆野核桃种群的数量动态除受基波影响外,并表现明显的小周期波动,具有大周期内有小周期的多谐波迭加特征,且在11龄级的小周期波动与野核桃高生长特性有关,在17龄级的小周期波动可能与野核桃的生理特性有关。
野核桃;种群;静态生命表;谱分析;坡向
年龄结构是种群的重要属性之一,其对种群的繁殖能力和可能的发展趋势起着决定性作用[1-2]。分析种群的年龄结构不仅可以了解种群现在的结构状态,重建种群的干扰历史,还可以预测未来种群结构的变化动态[3-4],有助于揭示种子散布、种子萌发及幼苗建立等特征在时间尺度上对植物种群动态变化的影响[5-6]。生命表是种群统计的核心[7-8],种群生命表可以提供许多关于种群年龄结构和数量统计方面的信息[3-4,9]。复杂的周期现象可以由不同振幅和相应的谐波组成,谱分析是探讨种群天然分布的波动性和年龄更替过程周期性的数学工具,种群天然更新动态可通过种群不同龄级的株数分布波动来表现[10-12]。
新疆野核桃(Juglans cathayensis)属胡桃科(Juglandaceae)、胡桃属植物,系国家Ⅱ级渐危物种[13]。它是中亚第三纪“残遗”阔叶树种与更新世北方“移民”的结合物,在原苏联境内的西天山和帕米尔-阿赖山地有较大的天然群落[14],在亚洲,成片分布的野核桃仅在中国新疆伊犁河谷巩留县野核桃沟。野核桃是栽培核桃的直接祖先[14],其独特的系统位置,在胡桃属植物的系统分类、种质资源保存上具有重要地位,并在核桃育种及栽培核桃的起源、演化研究上具有重要价值[15]。前人对新疆新疆野核桃的地理特征[16-17]、林下土壤形成[18-19]、群落学特征[14]、林学特性[15]、资源现状[20]、种质资源分类[21]、种子表型变异[22]、复叶的生长及生物量分配特征[23-24]、幼苗生长节律[25]等已有研究,从种群生命表及谱分析角度来研究其种群数量动态及周期性波动的报道较少。
对于数量较少的濒危乔木树种,若无明显可鉴定的外部形态标志,判定其实际年龄是困难的。将林木依胸径大小分级可建立胸径级结构,采用胸径级结构进行种群年龄结构特征分析同样揭示了种群的年龄分布[26-31]。以新疆野核桃自然保护区的野核桃为研究对象,以径级结构代表年龄结构,编制野核桃种群静态生命表,统计分析其数量特征,同时应用谱分析方法分析种群的波动周期,为了解野核桃种群的现存状态,预测未来种群发展趋势及揭示其濒危机制提供科学依据,并为资源植物新疆野核桃的保护和管理提供参考。
1 研究区概况
野核桃自然保护区位于新疆巩留县的凯特明山山前峡谷。地理位置:82°15′28″~82°17′23″E,43° 22′56″~43°25′40″N,总面积1 180hm2。保护区分为主沟、东沟、中沟和西沟[20],每条沟大体走向由北向南,海拔逐渐升高,整个沟谷为东、南、西三面陡峭的山梁所环抱,坡度30°~50°。春季和夏季多雨,冬季积雪0.7~1.0m。年均温7.6℃,1月均温-3.3℃,7月均温19.7℃,极端最低气温-25.3℃,≥10℃的积温1 865.4~2 338.9℃,年均降水量580 mm,年均蒸发量1 200mm,相对湿度70%~80%,无霜期约150d[17]。野核桃分布在较强的逆温层带之中,据伊犁州气象台1965~1969年探空观测记录,1月平均逆温强度为9.5℃,最大为22.6℃;1月逆温层平均厚度为950m,最大为2 077m[17]。现存野核桃集中分布面积约45hm2,计5 500余株[20,22],胸径≥2cm,主要分布在海拔1 250~1 550 m的沟谷、山坡。野核桃为该保护区优势种,伴生乔木主要有新疆野苹果(Malus sieversii)、野杏(Armeniaca vulgaris)、天山桦(Betula tianschanica)、欧洲山杨(Populus tremula)等;林下灌木主要有金丝桃叶绣线菊(Spiraea hypericifolia)、截萼忍冬(Lonicera altmannii)、野蔷薇(Rosa multiflo-ra)、异果小檗(Berberis heteropoda)、阿尔泰山楂(Crataegus altaica)等;草本植物主要有密丛雀麦(Bromus benekeni)、大羊茅(Festuca gigantea)、小叶羊角芹(Aegopodium alpestre)、短柄草(Brachypodium sylvaticum)、水金凤(Impatiens noli-tangere)等。
2 研究方法
2.1 野外调查方法
2012年5~10月,采用样带法和样方法相结合的调查方法,在野核桃自然保护区4条沟谷的阳坡和阴坡各设置1个调查样地,共8个样地。其中,根据野核桃分布的垂直高度不同,在主沟和中沟的阳坡和阴坡各设置3条样带,在东沟和西沟的阳坡和阴坡各设置4条样带,设置样带时尽量涵盖保护区主要乔木群落(野核桃、野杏、野苹果、欧洲山杨、天山桦),样带宽20m,样带长度为从各沟谷谷底到野核桃分布的上限,在每条样带以20m×20m为样方。阳坡70个样方,阴坡84个样方,对野核桃进行每木调查,记录胸径(DBH≥2cm)、树高、冠幅、枝下高、坡向,以及DBH<2cm的幼苗和幼树的基径、株高、株数等指标。同时对每一样地的其它乔木采用10m×10m、灌木采用5m×5m、草本层采用1m×1m的样方进行物种种类、密度、盖度调查,每个样地乔、灌、草各随机调查样方20个,并记录整个样地生境条件。
2.2 径级划分
野核桃种群胸径大小级别的划分参考王伯荪等[32]的方法,将阳坡野核桃种群划分为17个胸径级,阴坡种群划分为19个胸径级,第1径级为0~2 cm,即幼苗和幼树划为1个龄级,之后每级间隔5 cm,即2~7cm为第2径级、7~12cm为第3径级,依此类推。将径级从小到大的顺序看作时间顺序,即第1径级对应1龄级,第2径级对应2龄级,第3径级对应3龄级,如此类推,统计各龄级植株数。在进行年龄结构分析时,各龄级植株数均换算为单位面积(hm2)植株数。
2.3 静态生命表编制
静态生命表反映的是种群多个世代重叠的年龄动态历程中的一个特定时间,而不是对同生种群的全部生活史追踪,在生命表中会出现死亡率为负的情况[7]。Wratten[33]认为,生命表分析中产生一些负值与数据假设技术不符。对野核桃种群的调查材料采用匀滑技术[8]处理,以阴坡野核桃种群为例,具体方法如下:调查数据在2和15龄级发生波动,其小于3和16龄级的存活数。根据特定时间生命表假设,年龄组合是稳定的,各年龄的比例不变。因此,认定2个区段,1~4龄级和13~19龄级,分别计算2个区段存活数的累积=T2/n2=33/7=4.7≈5,据此认为这是区段的组中值;据区段的第一个存活数和最后一个存活数的差数(547-243=304和12-1=11)及区段的间隔数(4和7),可以确定每一相邻年龄组存活数之间的差数为76和2左右。经匀滑修正后得ax′,再据此编制种群静态生命表。
2.4 波动周期分析
种群数量波动周期采用谱分析法,谱分析是Fourier级数的展开。Fourier证明复杂的周期现象可以由不同振幅和相应的谐波组成[34],写成正弦波形式为:Nt=A0+∑ni=1AkSin(ωkt+θt),式中,A0为周期变化的平均;Ak(k=1,2,3,…,p)为各谐波的振幅,标志其所起作用的大小,其值的差异反映了各周期作用大小的差别;ωk及θt分别为谐波频率及相角;Nt为t时刻种群大小。种群动态的谱分析中各参数参考文献[11-12,34]的方法进行计算。
3 结果与分析
3.1 静态生命表分析
根据样方调查的野核桃,阳坡为1 538株,阴坡为1 927株,总共3 465株,其静态生命表见表1。阳坡野核桃种群最大为17龄级,对应DBH为81.5 cm,阴坡最大为19龄级,对应DBH为91.7cm,反映出阳坡野核桃种群相对缺乏大龄级树。从消失率反映出阴坡野核桃种群在低龄级(12龄级前)比阳坡的消失率大,而到高龄级(12龄级后)则比阳坡的消失率小,表明新疆野核桃在长成成年植株前,阴坡种群比阳坡种群更难通过环境筛的筛选,种群有较大的损失率。
3.2 期望寿命分析
以龄级为横坐标,以表1中各龄级野核桃的期望寿命(ex)为纵坐标作图。从图1可以看出,野核桃的期望寿命在不同坡向生境上均以1龄级最大,不同坡向同一龄级及同一坡向不同龄级野核桃的期望寿命均存在波动,表明野核桃种群在不同坡向经历环境筛的强度不同。在10龄级前,阳坡野核桃的期望寿命高于阴坡,而10龄级后为阳坡低于阴坡。种群的期望寿命总体上表现为低龄级较高,中龄级较低,中龄级后的几个龄级又相对较高,最后几个龄级较低。
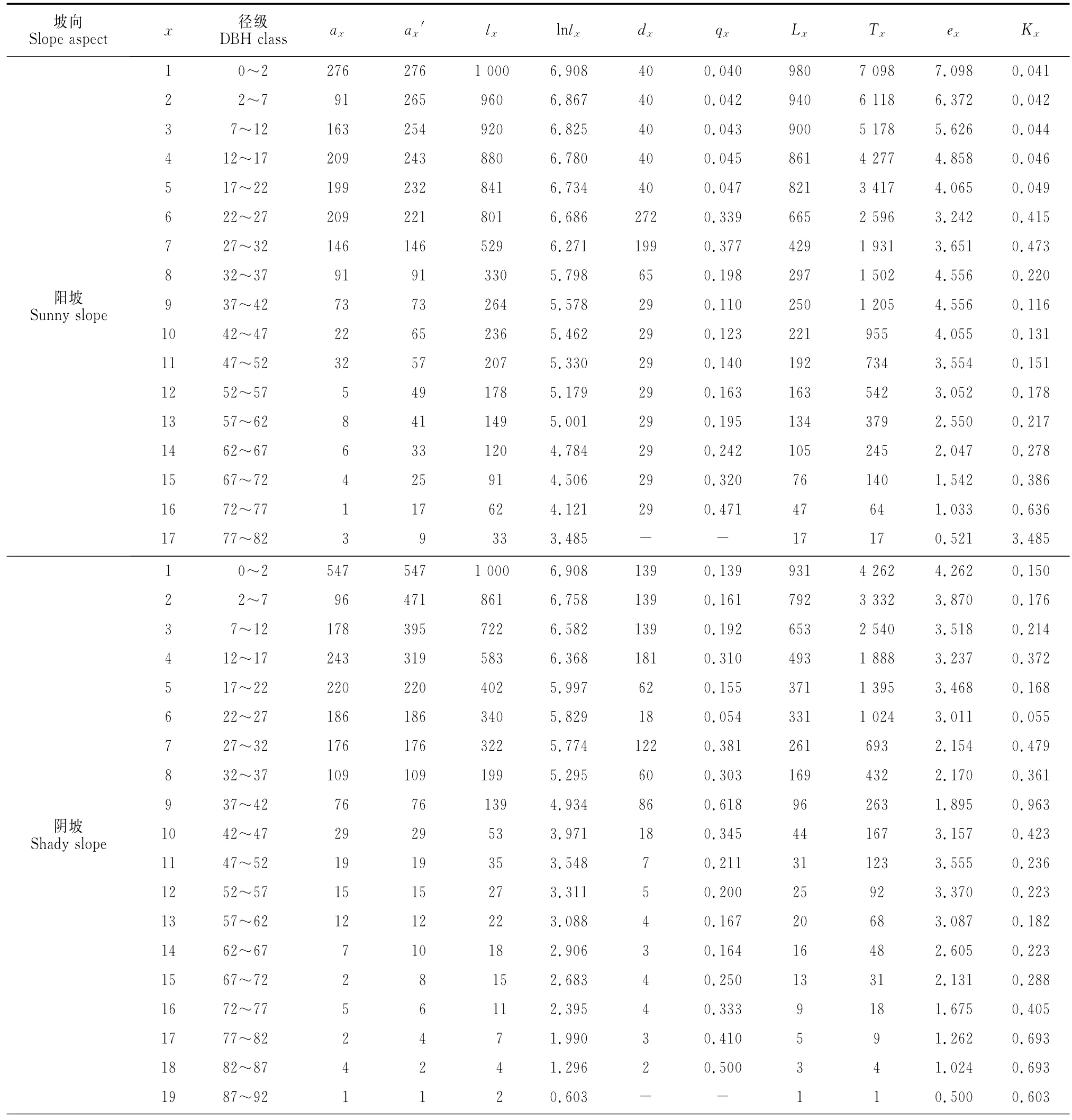
表1 不同坡向野核桃种群的静态生命表Table 1 Static life table of J.cathayensis populations in different slope aspects
3.3 年龄结构分析
以龄级为横坐标,单位面积各龄级植株数为纵坐标,绘制野核桃种群年龄结构图。从图2可以看出,阳坡和阴坡1龄级植株数均最多,2龄级植株数均明显少于1龄级,表明野核桃种群在发育初期,幼龄级植株比较丰富,以高死亡率为代价从幼龄植株发育为成年植株,进而完成整个生活史。例如,阴坡1龄级植株每hm2有163株,2龄级仅有29株,死亡率达到82.2%,同时也反映出种群幼龄植株到成年植株的发育是不连续的。图2还显示,阴坡1龄级植株数远比阳坡多,但2龄级植株数却比阳坡少,表明野核桃幼龄植株在阴坡比阳坡有更大的死亡率。阳坡和阴坡种群各龄级的植株数量不同,但年龄结构相似,植株数量集中于低等和中等龄级,1~9龄级的植株数分别占阳坡、阴坡的94.7%和94.9%,即以幼年和青壮年树占绝对优势,为增长型年龄结构,从野核桃种群总体1~9龄级植株的比例94.8%也显示种群为增长型年龄结构。
3.4 存活曲线分析
以龄级为横坐标,种群标准化的存活数量为纵坐标,绘制野核桃种群的存活曲线。从图3可看出,与经典的存活曲线相比,阳坡、阴坡和种群总体的存活曲线均介于Deevey-Ⅱ型和Ⅲ型之间。采用数学模型检验的方法[12],阳坡、阴坡和种群总体的模型的F值及R2值均为指数函数大于幂函数(表2),可见,阳坡、阴坡野核桃种群和种群总体的存活曲线均趋于Deevey-Ⅱ型。图3还显示,在野核桃种群整个生活史中,阳坡种群各龄级的存活率均高于阴坡;阳坡和阴坡种群的存活曲线均存在波动,且阳坡种群波动幅度更大。表明野核桃种群在其生长发育过程中可能有几次大的非随机生境变迁,而且这些生境变迁对阳坡野核桃种群影响更大。
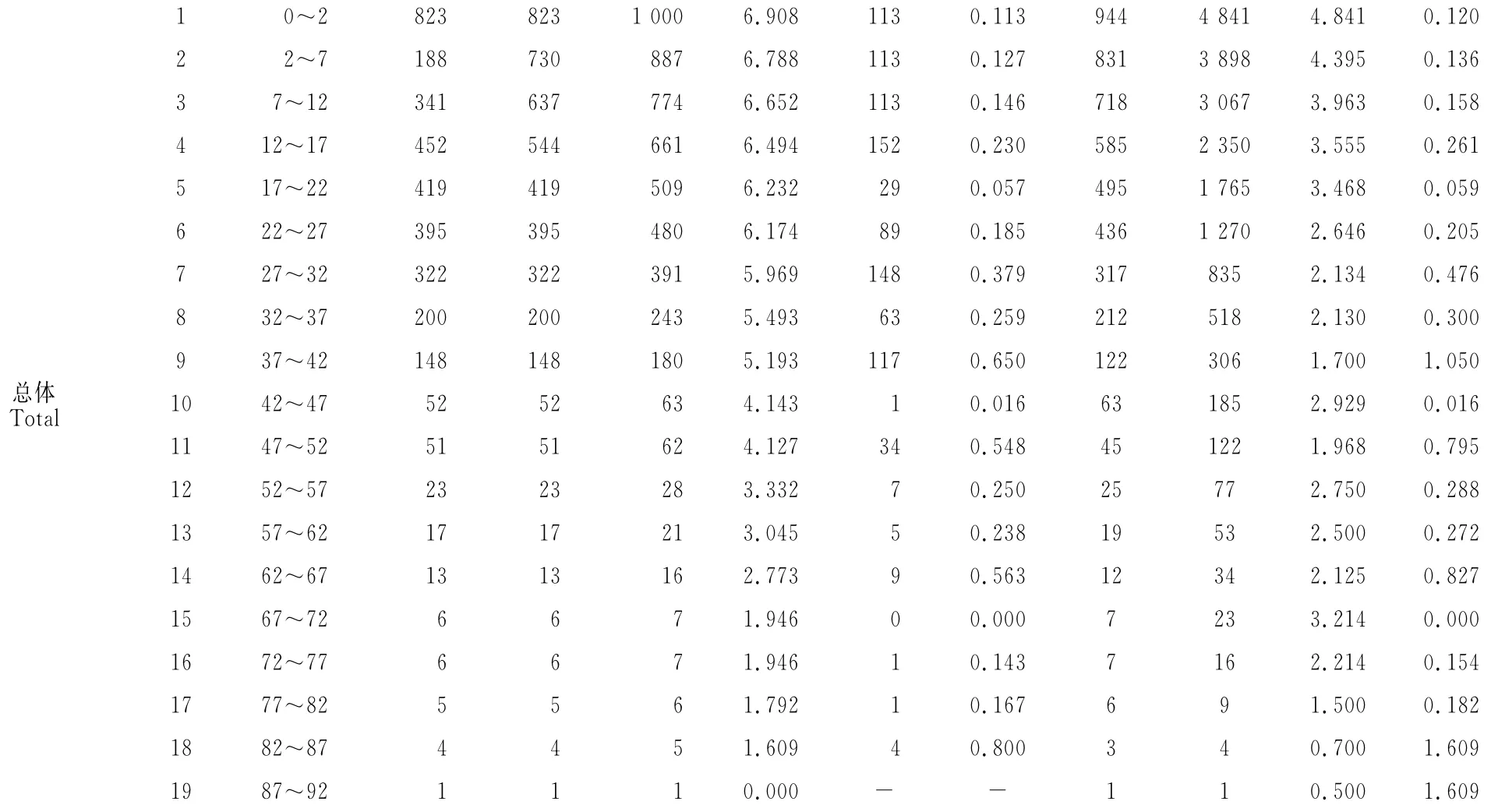
续表1 Continued Table 1
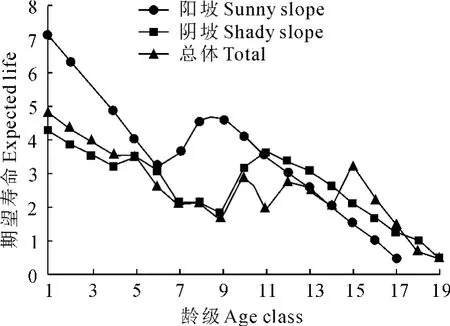
图1 不同坡向野核桃种群的期望寿命1~19.分别为不同龄级(径级),详见表1。下同Fig.1 Expected life of J.cathayensis populations in diffrent slope aspects 1-19represent different age classes(DBH classes),see Table 1.The same as below
3.5 死亡率曲线分析
以龄级为横坐标,死亡率(qx)为纵坐标,绘制野核桃种群死亡率曲线(图4)。阳坡种群存在7和16龄级2个死亡高峰,死亡最高峰在16龄级;阴坡种群存在4、7、9和18龄级等4个死亡高峰,死亡最高峰在9龄级,表明在不同坡向上,野核桃种群生活史中经历环境强烈筛选的龄级和次数不同。阳坡和阴坡有1个共同的死亡高峰7龄级,表明在阳坡和阴坡,野核桃种群在7龄级均受到环境筛的强烈作用。种群总体存在4、7、9、11、14和18龄级等6个死亡高峰,死亡最高峰在18龄级。表明野核桃种群在其生活史中,种群数量因野核桃自身生物学特性亦或外部环境因素多次发生较大变化。
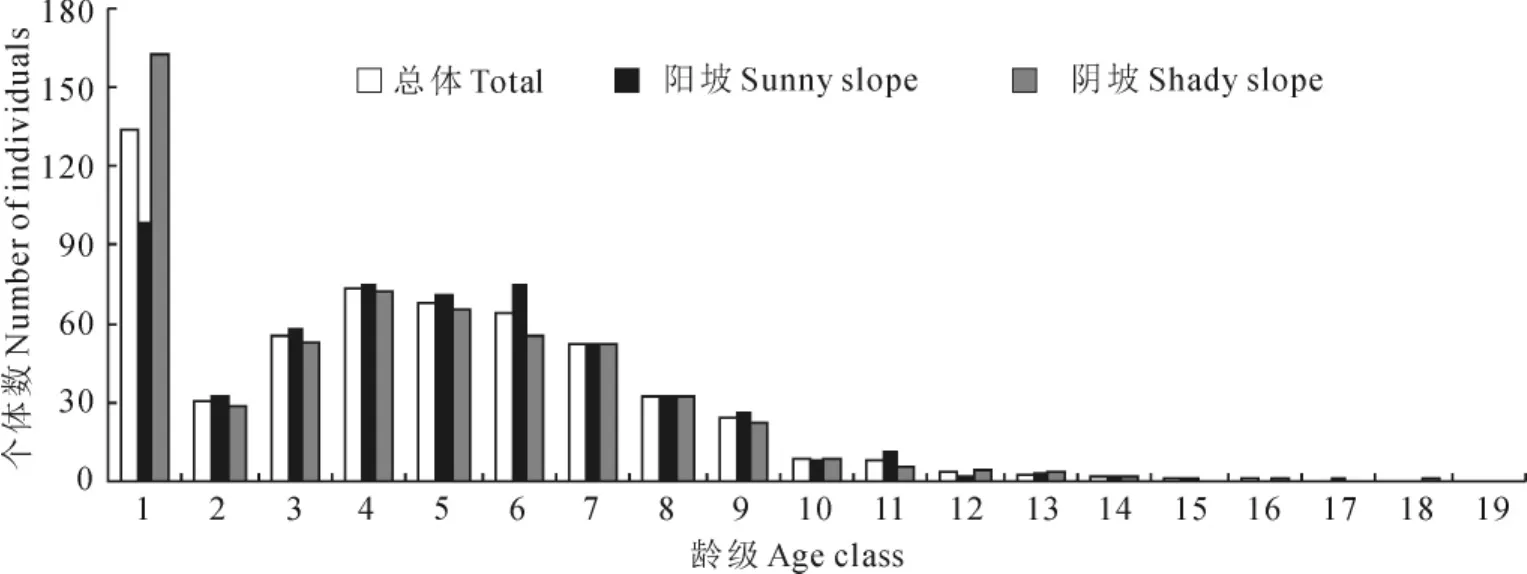
图2 不同坡向野核桃种群的年龄结构Fig.2 Age structures of J.cathayensis populations in different slope aspects

图3 不同坡向野核桃种群的存活曲线Fig.3 Survival curves of J.cathayensis populations in different slope aspects

表2 不同坡向野核桃种群存活曲线的检验模型Table 2 Test models of survival curves of J.cathayensis populations in different slope aspects

图4 不同坡向野核桃种群的死亡率曲线Fig.4 Curves of mortality of J.cathayensis populations in different slope aspects
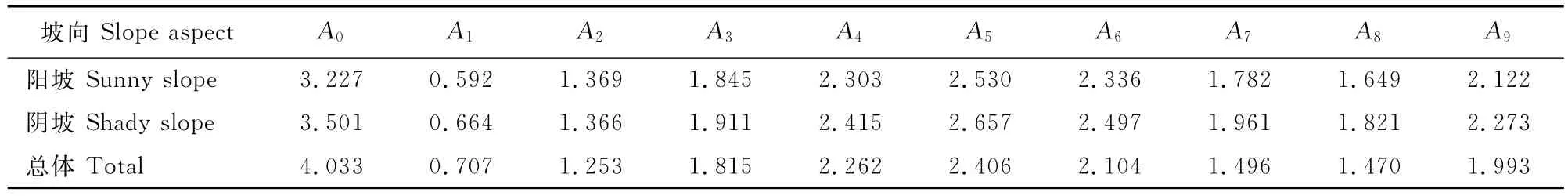
表3 不同坡向野核桃种群的周期性波动Table 3 Periodic fluctuations of J.cathayensis populations in different slope aspects
3.6 种群数量波动周期
将野核桃各龄级个体数在计算时进行对数化处理,即以Xt′=ln(ax+1)代换公式中的Xt。各个波形的振幅Ak值见表3。A0为基波振幅,A1~A9为谐波振幅。由于阳坡的龄级为17个,为了便于比较分析,采用内插法补充到18龄级,阳坡和阴坡的总波序K计算到9。
从振幅Ak值来看,各谐波不论是种群总体,还是阳坡和阴坡均表现为振幅存在周期性波动,且基波振幅A0为最大。基波表现了基本周期的波动,其周期长度为种群本身所固有,由种群波动特性所决定,表明野核桃种群数量动态变化明显受整个生命周期生物学特征的控制,但由于所调查野核桃种群年龄还不够长,最大径级DBH为91.7cm,时间系列长度不能表现出明显的固有波动周期长度。野核桃种群数量动态除受基波的影响外,还显示出明显的小周期波动,阳坡、阴坡和种群总体的小周期波动均发生在A6、A9,前者的波动约在空间序列DBH的49.5cm处,对应于11龄级,后者的波动约在空间序列DBH的79.5cm处,对应于17龄级。
4 讨 论
生命表显示,阴坡野核桃种群比阳坡大2个龄级,DBH相差达到10cm,推测其关键影响因子是水分,野核桃生长到最后几个龄级时,树高、冠幅等均较大,维持其生存和开花结实需要更多的水分,而阳坡的蒸发量远大于阴坡,野核桃需要的水分相对不足,致死大龄级数难以存活。这或许蕴含野核桃种群在不同坡向上的生长与物质分配策略,因为植物在生长发育过程中,总是不断调整其生长和物质分配策略,但也要遵循生长与物质分配相协调的原则[34]。
在不同坡向生境上,新疆野核桃种群的期望寿命在第10龄级是一个转折点,10龄级前种群期望寿命的关键影响因子可能是光照,阳坡较阴坡的光照相对充足,作为喜暖阔叶树野核桃有更大的期望寿命;10龄级后的关键影响因子可能是水分和养分,随着野核桃树体和树冠长大,需要更多的营养物质,而阳坡与阴坡相比,水分和养分相对不足,其期望寿命缩短。种群总体的期望寿命有几次较大波动,低龄级野核桃期望寿命较高,是因为野核桃植株处于幼年和青年期,树体和冠幅不大,生存空间比较充裕,种内和种间竞争不强,植株的期望寿命比较高;种群到中龄级,由于生存需要的光照、水分、养分和空间等不能满足所有植株需求,种内和种间竞争强烈,种群的自疏和它疏作用增强,致使植株的期望寿命减小;中龄级后几个龄级的植株是在种群进行自身数量调节后的稳定生活期,生存空间和养分、水分等又相对充足,期望寿命比较高;最后几个龄级植株已到生理衰退期,期望寿命随龄级增大而变小。
统计分析表明新疆野核桃种群为增长型年龄结构,理论上该野核桃种群在今后一定时期内,其种群数量将会逐渐增加。但是,近几年野核桃高昂的价格(300~400元/kg)致使人为对种子过度采摘,所剩不多的种子又被啮齿动物(野核桃林内松鼠比较多)捕食、腐烂以及其它原因,目前野核桃种群的种子最后能转化为幼苗和幼树的数量很少,2012年和2013年在野外调查时较难发现1和2年生幼苗,且野核桃由幼苗向幼株生长过程中,有一道较强的环境筛,植株很难通过而大量死亡,新疆野核桃种群的前景堪忧。
野核桃种群的死亡高峰在阳坡有2个,阴坡有4个,且二者的死亡最高峰不同,反映出在不同坡向生境上,种群受环境筛的作用强度不同,或者种群选择的生活史对策不同。其它山地乔木树种在不同坡向生境上的死亡高峰是否也存在类似差异需要进一步研究。阳坡的死亡最高峰在16龄级(死亡率47.1%),这个死亡高峰是种群进入生理衰退期引起的;阴坡的死亡最高峰在9龄级(死亡率61.8%),是阴坡野核桃种群在DBH级达到37~42cm时,种内或种间竞争激烈,种群进行自疏或它疏作用增强而引起的;其次为18龄级(50.0%),是因为种群进入生理衰退期而导致高的死亡率。种群总体的死亡最高峰在18龄级(死亡率80.0%),表明野核桃一旦在幼苗、幼树通过环境筛的筛选后,大多数个体可以存活到正常寿命。
对一个植物有机体来说,生活在一个复杂的群落中,它死亡的原因也是复杂的,植物的生长不仅受所生长的生物环境的影响,而且还受气候条件及自然灾害等的影响[26]。新疆野核桃种群1龄级植株(胸径2cm以下,1和2年生幼苗很少)虽然比较丰富,但死亡率较高,可能与野核桃喜光特性有关,阳性植物在郁闭度大的林冠下更新是比较困难的[35]。野核桃的伴生乔木多为生长迅速的耐阴常绿或落叶阔叶植物,如天山桦、欧洲山杨等,这些阔叶植物可能是导致种群幼苗高死亡率的重要原因。对野核桃天然林进行适度的人为干扰(择伐、间伐),以降低林地的郁闭度,可以提高野核桃幼苗向成年植株生长的成功率。尽管环境筛对种群有重要影响,但在自然保护区,野核桃以幼年、青壮年树为主,为增长型年龄结构,且野核桃为长寿命树种,是自然保护区植被的主要优势种,若无强烈人为干扰或严重自然灾害,将在群落中长期保持优势种地位。
植物种群动态的谱分析一般针对衰退型种群进行。吴明作等[12]对栓皮栎(Quercus variabilis)的研究表明,谱分析也能应用于增长型种群数量动态的研究。林木植株的增长和生活空间的扩展使种群各年龄植株的组配情况不断产生变化,最后由种群自身所创造的环境条件又反过来调节和控制其种群的数量,产生波动过程,这是稳定植物群落中优势种群的特征。从谱分析结果可看出,新疆野核桃种群数量动态是存在周期性波动的,而且其所表现出的波动不是单一周期,具有在大周期内有小周期的多谐波迭加特征。野核桃种群的波动发生在11龄级,这个小周期波动与野核桃该时期的高生长特性相一致,野核桃经过多个层次的筛选,一部分植株最终达到主林层,林木开始产生分化,林分由郁闭变为稀疏,种群形成一个相对快速的扩张期,这和野外实地调查是一致的,野核桃的DBH和冠幅较大的样地内,其种群密度相对较小;野核桃种群的波动发生在17龄级,这个小周期波动可能与野核桃的生理特性有关,经过漫长的生长过程,到了17龄级的野核桃已经进入生理衰退期,使得该种群数量又出现波动。
[1] LI X K(李先琨),SU Z M(苏宗明),XIANG W SH(向悟生),et al.Study on the structure and spatial pattern of the endangered plant population of Abies yuanbaoshanensis[J].Acta Ecologica Sinica(生态学报),2002,22(12):2 246-2 253(in Chinese).
[2] WANG ZH F(王峥峰),AN SH Q(安树青),ZHU X L(朱学雷),et al.Distribution pattern of tree populations in tropical forest and comparison of its study methods[J].Chinese Journal of Applied Ecology(应用生态学报),1998,9(6):575-580.
[3] HARPER J L.Population Biology of Plants[M].London:Academic Press,1977:599-643.
[4] STEWART G H,ROSE A B.The significance of life history strategies in the developmental history of mixed beech(Nothofagus)forest,New Zealand[J].Vegetation,1990,87:101-114.
[5] ARMESTO J J,CASSASSA I,DOLLENZ O.Age structure and dynamics of Patagonian beech forests in Torresdel Paine National Park,Chile[J].Vegetation,1992,98:13-22.
[6] SKOGLUND J,VERWIJST T.Age structure of woody species populations in relation to seed rain,germination and establishment along the river Dalalven,Sweden[J].Vegetation,1989,82:25-34.
[7] BEGON M,MORTIMER M.Population Ecology:A Unified Study of Animal and Plants[M].London:Blackwell Science Ltd,1981.
[8] 江 洪.云杉种群生态学[M].北京:中国林业出版社,1992:8-26.
[9] STEWART G H.The dynamics of old-growth Pseudotsugaforests in the western Cascade Range,Oregon,USA[J].Vegetation,1989,82:79-94.
[10] GUO J P(郭晋平),WANG SH H(王石会),KANG R L(康日兰),et al.Age structure and dynamics of natural spruce(Picea wilsonii)forests in Guancen Mountains[J].Acta Ecologica Sinica(生态学报),1997,17(2):184-189(in Chinese).
[11] WU Y G(伍业钢),XUE J X(薛进轩).Periodic fluctuation of Korean pine population of broad leaved Korean pine forest[J].Chinese Journal of Ecology(生态学杂志),1988,7(1):19-23(in Chinese).
[12] WU M Z(吴明作),LIU Y C(刘玉萃).Periodic fluctuation and stability of Quercus variabilis population[J].Chinese Journal of Ecology(生态学杂志),2000,19(4):23-26(in Chinese).
[13] 国家环境保护局,中国科学院植物研究所.中国珍稀濒危保护植物名录(第1册)[M].北京:科学出版社,1987:19.
[14] ZHANG X SH(张新时).On the eco-geographical characters and the problems of classification of the wild fruit-tree forest in the Ili valley of sinkiang[J].Acta Botanica Sinica(植物学报),1997,15(2):239-253(in Chinese).
[15] WANG L(王 磊),CUI N R(崔乃然),ZHANG H F(张汉斐).Research report of Xinjiang wild walnuts[J].Arid Zone Research(干旱区研究),1997,14(1):17-27(in Chinese).
[16] XU D Y(徐德炎).Study on eco-climatic features of Xinjiang wild walnuts[J].Chinese Journal of Ecology(生态学杂志),1989,8(4):24-27(in Chinese).
[17] XU D Y(徐德炎),ZHU X ZH(朱晓专).Study on the survive and multiply ecological conditions of Xinjiang wild walnuts[J].Forest by-Product and Speciality in China(中国林副特产),1991,(4):1-6(in Chinese).
[18] LIU L CH(刘立诚),PAI Z L(排祖拉),XU H J(徐华君).The formation characteristics and taxonomy of the soil under the wild-walnut forest in Ili vally[J].Journal of Xinjiang University(新疆大学学报),1998,(2):60-65(in Chinese).
[19] SHE D Y(佘定域).Formation and characteristics of wild walnut forest soils in Gongliu County[J].Arid Zone Research(干旱区研究),1994,11(2):11-15(in Chinese).
[20] DONG Y ZH(董玉芝),ZHU X H(朱小虎),CHEN H(陈 虹),et al.Investigation and analysis on the wild walnut in Gongliu,Xinjiang[J].Chinese Journal of Plant Genetic Resources(植物遗传资源学报),2012,13(3):386-392(in Chinese).
[21] WANG L(王 磊),LI X(李 霞),YANG L(杨 辽),et al.Study on the germplasm resources quantitative classification of Xinjiang wild walnuts[J].Northern Horticulture(北方园艺),1998,(1):2-5(in Chinese).
[22] ZHANG W(张 维),LUO X Z(罗新泽),ZHANG N(张 娜),et al.Phenotypic variability and growth characteristics of wild walnut seeds in Xinjiang,Northwest China[J].Chinese Journal of Ecology(生态学杂志),2013,32(9):2 281-2 288(in Chinese).
[23] ZHANG W(张 维),REN Y L(任艳利),ZHAO Y(赵 玉),et al.Biomass plasticity and allocation rules for modules of compound leaves with a different number of leaflets on Juglans cathayensis in Xinjiang[J].Journal of Northeast Forestry University(东北林业大学学报),2012,40(7):37-40(in Chinese).
[24] ZHANG W(张 维),ZHAO Y(赵 玉),ZHANG X F(张相锋),et al.Phenotypic variability and growth regulation on the compound leaf of Juglans regiain Yili of Xinjiang[J].Journal of Northeast Normal University(东北师大学报),2011,43(1):113-117(in Chinese).
[25] ZHANG W(张 维),JIA F Q(贾风勤),NASEN B T(纳森巴特),et al.Population dynamics and growth of Juglans cathayensis seedlings of different slope aspects in the west Tianshan valley in Xinjiang,China[J].Chinese Journal of Ecology(生态学杂志),2014,33(10):2 596-2 602(in Chinese).
[26] BI R CH(毕润成).Ecological feature and floral analysis of Juglans mandshurica community in Huoshan Mountain of Shanxi Province[J].Chinese Journal of Applied Ecology(应用生态学报),1999,10(6):650-656(in Chinese).
[27] LI X X(李晓笑),WANG Q CH(王清春),CUI G F(崔国发),et al.Structure and dynamic characteristics of a wild population of endangered Abies fanjingshanensis[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2011,31(7):1 479-1 486(in Chinese).
[28] LIU G F(刘贵峰),CHENG W Y(程伟燕),LIU Y P(刘玉平),et al.Population structure and dynamic of domiant species of main forest community in Daqinggou Nature Reserves[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2014,34(4):843-850(in Chinese).
[29] YUAN H B(袁宏波),ZHANG J CH(张锦春),CHU J M(褚建民),et al.Characteristics of the age structure of three typical plant populations in Kumtag Desert[J].Acta Bot.Boreal.-Occident.Sin.(西北植物学报),2011,31(11):2 304-2 309(in Chinese).
[30] HU E CH(胡尔查),WANG X J(王晓江),ZHANG W J(张文军),et al.Age structure and point pattern of Butula platyphyllain Wulashan Natural Reserve of Inner Mongolia[J].Acta Ecologica Sinica(生态学报),2013,33(9):2 867-2 876(in Chinese).
[31] LI Y L(李艳丽),YANG H(杨 华),KANG X G(亢新刚),et al.Population structures and dynamics of Abies nephrolepis and Picea koraiensis in the Changbai Mountains of Northeastern China[J].Journal of Beijing Forestry University(北京林业大学学报),2014,36(3):18-25(in Chinese).
[32] 王伯荪,李鸣光,彭少麟.植物种群学[M].广州:广东高等教育出版社,1995.
[33] WRATTEN S D,FRY G I A.Field and Laboratory Exercises in Ecology[M].London:Edward Arnad,1996:98-103.
[34] HONG W(洪 伟),WANG X G(王新功),WU CH ZH(吴承祯),et al.Life table and spectral analysis of endangered plant Taxus chinensis var.mairei population[J].Chinese Journal of Applied Ecology(应用生态学报),2004,15(6):1 109-1 112(in Chinese).
[35] FENG L(封 磊),HONG W(洪 伟),WU CH ZH(吴承祯),et al.Study on the dynamics of the endangered plant population of Tsuga tchekiangensis[J].Chinese Journal of Wuhan Botanical Research(武汉植物学研究),2003,21(5):401-405(in Chinese).
(编辑:潘新社)
Life Tables and Fluctuation Cycle of Juglans cathayensis Populations on Different Slope Aspects in Wild Walnut Natural Conservation Area of Xinjiang,China
ZHANG Wei1,2,JIAO Ziwei1,REN Yanli1,YANG Yunfei2*,LI Jiandong2
(1School of Chemistry and Bioscience,Yili Normal College,Yining,Xinjiang 835000,China;2Ministry of Education Key Laboratory of Vegetation Ecology,Institute of Grassland Science,Northeast Normal University,Changchun 130024,China)
For grasping fluctuation cycle of Juglans cathayensis populations and forecasting change of the population structure,we conducted a large sample survey on sunny and shady slopes in valley of conservation area in Xinjiang,Using the diameter class structure representing the age structure,we drew up static life table,life expectancy chart,age structure plot,survival curve,and death rate curve chart of J.cathayensis populations.Also,we compared the fluctuation cycle of J.cathayensis populations on different slope aspects by using spectrum analysis method.The results showed that:(1)life expectancies of J.cathayensis populations were fluctuant,both on different slopes of the same age and the same slope of different ages.Life expectancy before 10th age class was higher on sunny slope than that of shady,but after 10th age classit was lower than that of shady slope.(2)J.cathayensis populations on different slope aspects showed increasing age structure.(3)Survival curve of J.cathayensis populations on different slope aspects tended to be Deevey-Ⅱtype,and the survival rate of each age class on sunny slope was higher than that on shady slope.(4)The 1st age plant developed into saplings at the cost of high mortality.The populations in sunny slope had two death peaks in their life history,with the highest peak in 16th age class.However,the shady populations had four death peaks,with the highest peak in 9th age class.(5)Spectrum analysis showed that the quantitative dynamics of J.cathayensis populations were influenced by fundamental wave and also showed obvious small periodic fluctuation,and they had characteristics of multi harmonic superposition that small periodic fluctuation in big one.In the meantime,the small periodic fluctuation of 11th age class was related to high growth characteristic of J.cathayensis,but it might be related to physiological property in 17th age class.
Juglans cathayensis;population;static life table;spectrum analysis;slope aspect
Q948.15+4
A
10.7606/j.issn.1000-4025.2015.06.1229
1000-4025(2015)06-1229-09
2014-12-08;修改稿收到日期:2015-04-10
国家自然科学基金(31160072);新疆教育厅普通高等学校重点实验室项目(2013YSHXYB07)
张 维(1973-),男,在读博士研究生,副教授,主要从事植物种群生态学研究。E-mail:zhangw891@nenu.edu.cn
*通信作者:杨允菲,教授,博士生导师,主要从事植物种群生态学和草地生态学研究。E-mail:yangyf@nenu.edu.cn
