杂色鲍精氨酸激酶基因的克隆及其在不同应激条件下的表达
2015-06-23李文辉朱友芳张子平王艺磊
李文辉,张 鑫,朱友芳,张子平,王艺磊*
(1.莆田市水产科学研究所,福建莆田351100;2.集美大学水产学院,农业部东海海水健康养殖重点实验室,福建厦门361021)
杂色鲍精氨酸激酶基因的克隆及其在不同应激条件下的表达
李文辉1,张 鑫2,朱友芳1,张子平2,王艺磊2*
(1.莆田市水产科学研究所,福建莆田351100;2.集美大学水产学院,农业部东海海水健康养殖重点实验室,福建厦门361021)
采用c DNA末端快速扩增(SMART-RACE)技术克隆了杂色鲍(Haliotis diversicolor)精氨酸激酶(Hd AK)基因;利用实时荧光定量PCR(qRT-PCR)技术检测了Hd AK基因在各组织中的表达量,并分析了高水温、低溶氧、高水温与低溶氧联合应激及副溶血弧菌(Vibrio parahaemolyticus)感染对鳃组织和血细胞Hd AK基因表达量的影响.结果显示:Hd AK基因的cDNA全长1 595 bp,编码354个氨基酸.所测定的各组织中Hd AK基因均有表达,与其他组织相比在肝胰腺中的表达量显著提高(p<0.05).在高水温、低溶氧、高水温与低溶氧联合应激及副溶血弧菌感染实验中,杂色鲍鳃组织和血细胞的Hd AK基因在实验的部分时段显著高于对照组,说明杂色鲍Hd AK不但参与能量代谢调节,而且参与了杂色鲍对高水温、低溶氧等不利环境的适应过程及机体免疫反应过程.研究结果可为揭示高水温、低溶氧、高水温与低溶氧联合应激及副溶血弧菌感染对杂色鲍的生理影响的机制提供理论依据.
杂色鲍;精氨酸激酶基因;高水温应激;低溶氧应激;高水温与低溶氧联合应激;副溶血弧菌感染
精氨酸激酶(arginine kinase,AK)为磷酸激酶家族中的重要一员,广泛存在于无脊椎动物中,是无脊椎动物体内调节能量代谢重要的酶之一[1],目前已有多种软体动物的AK cDNA序列信息,包括泥蜡(Tegillarca granosa)[2]、长牡蛎(Crassostrea gigas,登录号: AB118650)、池蝶蚌(Hyriopsis schlegelii,登录号: JF803293)、曼氏无针乌贼(Sepiella maindroni,登录号: HQ650787)、日本大鲍(Haliotis madaka,登录号: D26104)、光滑双脐螺(Biomphalaria glabrata,登录号: GQ453432)、黑斑海兔(Aplysia kurodai,登录号: AB059837)[3]等.现有研究表明,AK在水生动物机体适应低溶氧[4-5]等不良的环境变化、机体的免疫、机体防御病原感染[6-8]等方面发挥重要的作用.
杂色鲍(Haliotis diversicolor)为我国南方重要的水产养殖种类,但在夏季由于高水温、低溶氧及由弧菌引起的疾病严重制约了杂色鲍养殖的可持续发展[9],为此进行环境应激及病原感染对杂色鲍生理影响的研究是必要的,目前已进行了环境应激及病原感染与杂色鲍相关基因表达关系的研究,如高水温及低溶氧应激对核因子κB(NF-κB)、核因子抑制蛋庄(IκB)和Akirin2[10]、缺氧诱导因子-1α(H d HIF-1α)[11]、杂色鲍紫色酸性磷酸酶(Hd PAP)[12]、酪氨酸3-加单氧酶/色氨酸5-加单氧酶激活蛋庄ζ(H d 14-3-3ζ)[13]、同种移植炎症因子(H d AIF-1)[14]、热休克转录因子1(H d HSF1)和热休克蛋庄90 (Hd HSP90)[15]等基因的表达的影响;副溶血弧菌感染对Hd PAP基因[12]、H d AIF-1α[11]、NF-κB、IκB、Akirin2基因[10]和β-1,3-葡聚糖识别蛋庄基因(sa-βgrp)[16]表达的影响等.但杂色鲍AK(Hd AK)基因克隆及高水温、低溶氧、弧菌感染对Hd AK基因表达的影响未见报道.为了给环境应激及病原感染对杂色鲍生理影响机制的研究提供理论依据,本研究采用cDNA末端快速扩增(SMART-RACE)技术克隆了Hd AK基因并以该基因为研究对象,利用实时荧光定量PCR(qRT-PCR)技术检测了在高水温、低溶氧、高水温与低溶氧联合应激及副溶血弧菌(Vibrio parah-aemolyticus)感染时基因的表达量.

1 材料与方法
1.1 材 料
1.1.1 杂色鲍
实验用杂色鲍体长(61.0±5.0)mm,质量(18.7 ±2.5)g,购回实验室后在25℃的海水循环系统中暂养,暂养期间每日喂1次海带(Laminaria japonica),暂养10 d后进行实验,实验用水符合海水养殖用水水质要求.
1.1.2 试 剂
本研究所用的逆转录酶M-MLV、SYBR Green Realtime PCR Master Mix、琼脂糖凝胶DNA回收试剂盒以及p MD19-T连接试剂盒分别购自Promega公司(美国)、捷瑞生物工程(上海)有限公司和宝生物(大连)公司.
1.1.3 所用引物
本研究用的引物全部由捷瑞生物工程(上海)有限公司合成.3′RACE outer primer和3′RACEinner primer序列分别为5′-GACCATCTCCGTCTCATCTCCA-3′和5′-GACATTCTGCCCCAGCAACTT-3′,head primer和toe primer序列分别为5′-ATGGGTACTGAAAAACAGAAGT-3′和5′-TCAATTCTGTTCTTGCTGTTCAC-3′,Hd AK RTF和Hd AK RTR序列分别为5′-ACGTCCTCGATGCGGTTATC-3′和5′-ACTGAACTTCTCGAAGGCGG-3′.Universal Primer Mix、Nested U-niversal primer、5′CDS primer、3′CDS primer、SMARTⅡ、β-actin-F和β-actin-R的序列与文献[12]相同.
1.2 方 法
1.2.1 高水温、低溶氧、高水温与低溶氧联合应激及副溶血弧菌感染实验
在高水温实验中,以水温25℃为对照组,实验组每小时升1℃至31℃进行0 h取样并开始计时;低溶氧应激、高水温与低溶氧联合应激及副溶血弧菌感染实验参照文献[10]的方法.低溶氧应激实验组溶解氧为2 mg/L,对照组为空气曝气(溶氧约为5.5 mg/L);高水温与低溶氧联合应激实验组升温及计时方法与高水温实验相同,溶解氧为2 mg/L,对照组水体经空气曝气(溶氧约为5.5 mg/L)、水温维持在25℃;副溶血弧菌感染实验组每只杂色鲍注射50μL浓度为6.7 ×107cfu/m L的副溶血孤菌,对照组注射50μL生理盐水.鲍血细胞是先天免疫防御系统的主要免疫细胞,鳃组织为水生动物与环境接触的第一屏障,因此本研究以杂色鲍血细胞、鳃组织为对象研究H d AK基因的表达量变化.实验组与对照组在每个采样时间点分别取5只,血淋巴经离心后弃上清,所得的血细胞保存于¯80℃冰箱中;鳃组织保存于液氮中,用于RNA的提取.
1.2.2 总RNA的提取及cDNA的合成
根据王艺磊等[17]实验方法,提取杂色鲍鳃组织以及血细胞总RNA,按照逆转录试剂盒的说明进行cDNA的合成.
1.2.3 Hd AK基因cDNA全长序列的克隆
采用SMART-RACE的方法获得H d AK基因的cDNA全长序列,利用head to toe引物验证其开放阅读框(ORF)的准确性.
1.2.4 Hd AK的生物信息学分析
用Blast(http:∥blast.ncbi.nlm.nih.gov/Blast. cgi)和ORF Finder(http:∥ww w.ncbi.nlm.nih.gov/ projects/gorf/orfig.cgi)等工具验证所得片段是否为目的基因;用Ex PASy(http:∥web.expasy.org/compute_pi/)推测Hd AK的等电点及分子质量;用SingalP 4.1 Server(http:∥www.cbs.dtu.dk/services/ SignalP/)判断有无信号肤序列;用NetPhos 2.0 Server(http:∥www.cbs.dtu.dk/services/Net-Phos/)对磷酸化位点进行预测;用Net NGlyc1.0 Server(http:∥w ww.cbs.dtu.dk/services/Net NG-lyc/)对糖基化位点进行预测;用BioEdit软件进行序列的多重比对;用MEGA5.05软件构建系统进化树.
1.2.5 Hd AK基因在杂色鲍各组织及不同应激条件下的表达
以β-actin作为内参基因,H d AK基因表达量测定的q RT-PCR反应混合液为SYBR Green Realtime PCR Master Mix 10μL、10μmol/L的Hd AK RTF primer和Hd AK RTR primer各0.5μL以及cDNA模板9μL.反应条件为95℃1 min;后95℃15 s,60℃1 min,40个循环.利用SPSS 20.0进行显著性差异分析,p<0.05为差异显著.
2 结 果
2.1 Hd AK基因的克隆、序列分析及空间结构模拟
通过RACE的方法获得了H d AK(Gen Bank登类聚为一大支,而杂色鲍首先和日本大鲍聚为一小支,然后和其他软体动物单独聚为一大支.录号:KM433667)的cDNA全长为1 595 bp,其中, 5′非编码区(5′UTR)、3′UTR和ORF分别为49, 481和1 065 bp.编码的氨基酸为354个(图1). H d AK分子质量为39.7 ku,等电点6.24,不含信号肤序列.由图1可见,H d A K有5个丝氨酸磷酸化位点、5个苏氨酸磷酸化位点和6个酪氨酸磷酸化位点.
2.2 编码AK蛋白序列的多重比对以及系统进化树的构建
图2为多重序列比对结果,由图中可见AK蛋庄具有较高的保守性,但其氨基酸的长度仍有一定的差异.Blast分析得出,H d AK与同为鲍属的日本大鲍的AK相似性最高,达到了91%.
图3为系统进化树.图中可见,节肢动物和线虫

图1 杂色鲍Hd AK基因c DNA及其氨基酸序列Fig.1 The cDNA and amino acid sequence of Hd AK gene from H.diversicolor

图2 Hd AK和其他物种AK氨基酸序列的多重比对Fig.2 Multiple alignment of the AK amino acid sequence between H.diversicolor and other species

图3 Hd AK和其他物种AK氨基酸序列系统发育树Fig.3 Phylogenetic tree of the AK amino acid sequences between H.diversicolor and other species
2.3 杂色鲍不同组织Hd AK基因的表达
由图4可见,所测定的各组织中Hd AK基因均有表达,与其他组织相比在肝胰腺中的表达量显著提高(p<0.05).
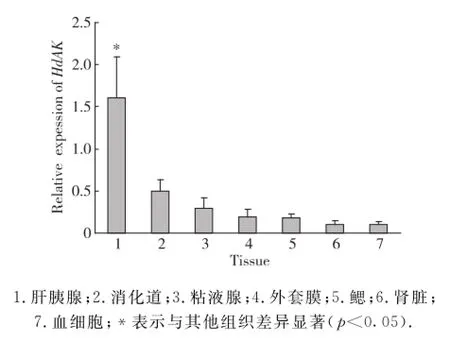
图4 Hd AK基因在杂色鲍各组织器官的表达情况Fig.4 Distribution pattern of Hd AK in different tissues of H.diversicolor
2.4 Hd AK基因在高水温、低溶氧、高水温与低溶氧应激以及副溶血弧菌感染后的表达
高水温应激开始阶段,Hd AK基因在杂色鲍鳃组织中的表达量不存在显著性差异.当应激时间达到4 h,H d AK基因的表达量显著性上调,之后又迅速恢复到和对照组相似的水平(图5(a)).在血细胞中, Hd AK基因的表达量分别在0 h和4 h表达量显著上调,随后又迅速恢复到对照组的表达水平(图5(b)).
qRT-PCR结果显示,低溶氧环境下,H d AK基因在鳃组织中的表达量分别在4,96和192 h显著上调,并且在192 h,表达量上升达到顶峰(图6(a)).在低溶氧诱导的起始阶段,H d AK基因在血细胞中的表达量未检测到显著性变化,当处理达到24 h,实验组的表达量显著上调,随后,在96和192 h虽然表达量有所下降但这一显著性差异依旧存在(图6(b)).
高水温与低溶氧联合应激条件下,在鳃组织中在4和192 h时实验组显著高于对照组(p<0.05)(图7 (a)).在血细胞中,在0和4 h,实验组显著高于对照组(p<0.05)(图7(b)).
在副溶血弧菌感染后,在实验组杂色鲍鳃组织中Hd AK基因的表达量分别在3,6和12 h时显著高于对照组,24 h这一显著性差异消失(图8(a)).在血细胞中,处理3 h,H d AK基因的表达量显著上调,之后的时间内,虽然实验组H d AK基因的表达量有所下降,但这一显著性变化在6和12 h仍然存在,处理24 h,随着Hd AK基因的表达量的继续下调,这一显著性差异消失,并恢复到与对照组无显著性差异的水平(图8(b)).

图5 高水温应激后各时期Hd AK基因在鳃组织(a)及血细胞(b)中的表达Fig.5 The change of Hd AK after thermal stress in gills(a)and haemocytes(b)
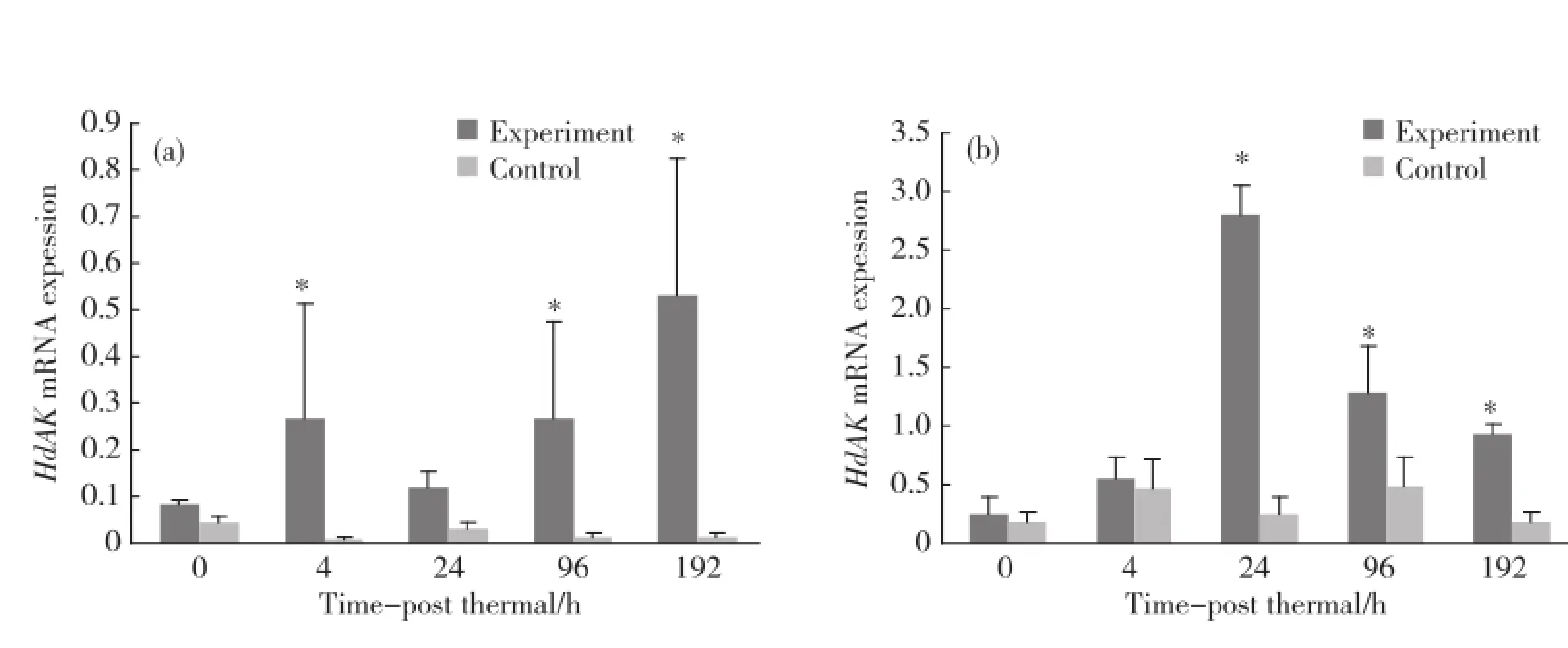
图6 低溶氧应激后各时期Hd AK基因在鳃组织(a)及血细胞(b)中的表达Fig.6 The change of Hd AK after hypoxia stress in gills(a)and haemocytes(b)
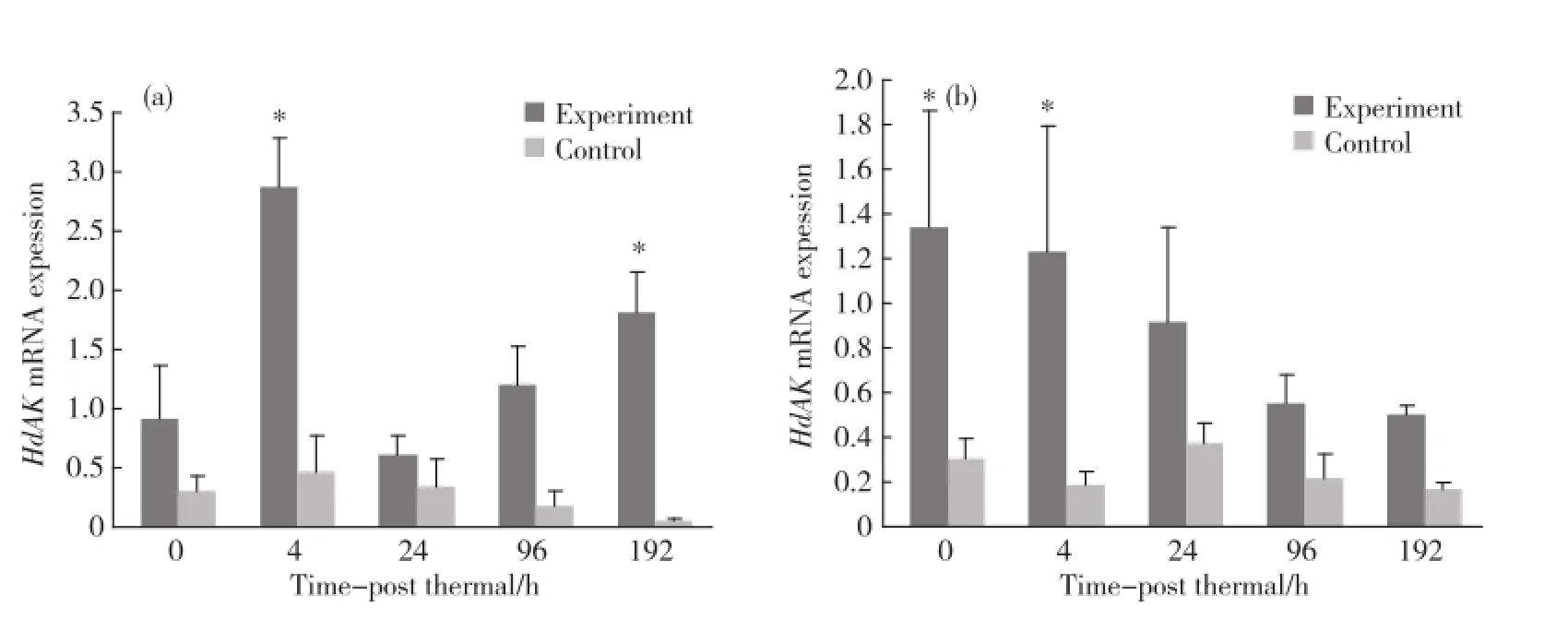
图7 高水温与低溶氧联合应激处理后Hd AK基因在鳃组织(a)及血细胞(b)中的表达Fig.7 The change of Hd AK after thermal&hypoxia stress in gills(a)and haemocytes(b)
3 讨 论
Hd AK的cDNA全长为1 595 bp,其中,5′UTR为49 bp,3′UTR为481 bp以及1 065 bp的ORF,编码354个氨基酸,Hd AK基因推导的氨基酸序列与其他动物的AK氨基酸序列有较高的相似度,特别是与日本大鲍的相似性达到91%,说明AK具较高的保守性.但氨基酸的多重比对表明不同物种AK氨基酸的长度也存在一定的差异.
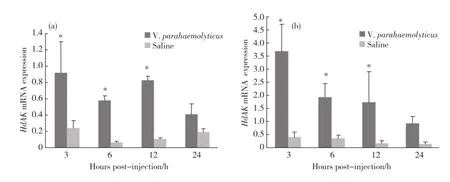
图8 副溶血弧菌感染后Hd AK基因在鳃组织(a)及血细胞(b)中的表达Fig.8 The change of Hd AK after V.parahaemolyticus challenge in gills(a)and haemocytes(b)
一般来说,AK的酶活性中心区域都具有7个氨基酸残基序列(CPTNLGT),酶活性中心位点都是由高度保守的半胱氨酸残基(C)组成[18],但有些种类的氨基酸残基序列第3位苏氨酸(T)被丝氨酸(S)代替,并不影响其功能.本研究中的杂色鲍H d AK活性中心氨基酸残基序列即为CPSNLGT(266~272),与日本大鲍、光滑双脐螺、黑斑海兔[3]、曼氏无针乌贼的序列相同.
已有的研究结果表明,在大多数无脊椎动物体内各组织中,AK基因均有表达[19],本次测定结果也证实了这一点,在本次测定的7种组织中,AK基因均有表达.在三疣梭子蟹(Portunus trituberculatus)、凡纳滨对虾(Litopenaeus vannamei)等甲壳类在肝胰腺中AK基因表达量显著低于鳃组织和血细胞[8,20],而Hd AK在肝胰腺中表达量则显著高于鳃组织和血细胞,这可能与甲壳类游泳生活而杂色鲍的匍匐生活有关.Hd AK在肝胰腺中表达量最高说明肝胰腺在调节能量代谢、适应不良的环境变化、防御病原侵袭等方面发挥了很大的作用.
日本囊对虾(Marsupenaeus japonicus)在缺氧条件下,肌肉中AK基因的表达量明显上调,肌肉中磷酸精氨酸的含量明显下降[5];红鲍(H.rufescens)在缺氧时AK上升[4];凡纳滨对虾在WSSV感染时肌肉的AK的表达量上调[7];三疣梭子蟹在溶藻弧菌(V. alginolyticus)感染后肌肉的AK的表达量显著上调[8];感染WSSV病毒的细角滨对虾(P.stylirostris),AK基因的表达量高出对照组2.84倍[6],这些研究表明,AK基因不但在生物体适应不利环境中起作用,而且还参与机体的免疫反应[18].本研究以杂色鲍血细胞、鳃组织为研究对象检测其在高水温、低溶氧、高水温与低溶氧联合应激和副溶血弧菌感染各采样时间点Hd AK的cDNA的表达量变化情况,结果表明在各实验中杂色鲍血细胞、鳃组织的Hd AK的cDNA的表达量在部分时段显著高于对照组,说明该基因参与了高水温、低溶氧等不利环境的适应过程及免疫反应过程,Hd AK对环境应激及副溶血弧菌感染反应敏感,可作为判断环境因素及细菌感染是否对杂色鲍产生应激的依据之一.
高水温及低溶氧应激抑制了杂色鲍H d PAP基因的表达[12],提高了H d 14-3-3ζ[13]、Hd AIF-1[14]、Hd HIF-1α[11]、Hd HSF1和Hd HSP90[15]、NF-κ、BIκB、NF-κB和Akirin2[10]等基因的表达水平;在副溶血弧菌感染下Hd PAP[12]表达抑制,AIF-1、NF-κB、IκB、NF-κB、Akirin2[10]和sa-β-grp[16]等基因表达上调.以上研究结果表明,杂色鲍在高水温、低溶氧、高水温与低溶氧联合应激及副溶血弧菌感染时可诱导参与多种生物学过程的基因表达水平发生变化,本实验的结果显示杂色鲍在高水温、低溶氧、高水温与低溶氧联合应激以及副溶血弧菌感染应激时Hd AK基因表达水平上升,基因表达水平的变化增加了杂色鲍的能量供应,杂色鲍对高水温、低溶氧、高水温与低溶氧联合应激以及副溶血弧菌感染的响应是一个耗能的过程.
在高水温及高水温与低溶氧联合应激实验中,血细胞H d AK基因0 h表达量显著上升,低溶氧应激实验中在0 h血细胞Hd AK基因表达量却没有上升,可见血细胞H d AK基因在实验升温过程中已产生应激反应,每小时升1℃由25℃升至31℃的升温方式已影响到杂色鲍的正常生理代谢.这一结果与鳃部Hd AIF-1基因在高水温实验升温过程表达量显著上调的趋势相似[14].
鳃组织H d AK基因表达量在高水温、低溶氧、高水温与低溶氧联合应激及副溶血弧菌感染时出现显著上升开始的时间分别为4,4,4和3 h,而血细胞H d AK基因表达量则分别在0,24,0和3 h时显著上升.由此可以得出,血细胞Hd AK基因对高水温、高水温与低溶氧联合应激的反应比鳃组织Hd AK基因敏感,与血细胞Hd AK基因相比鳃组织Hd AK基因对溶氧的变化更为敏感,鳃组织H d AK基因和血细胞Hd AK基因在弧菌感染时敏感性没有显著的差异.在高水温应激实验中,鳃组织Hd AIF-1基因在升温过程中表达量出现显著性上调[14],Hd HIF-1α基因在升温过程中表达量没有显著上调,直至水温稳定在31℃后1 h才出现显著上调[11],本次实验中鳃组织Hd AK基因在水温升至稳定在31℃后4 h才出现显著上调.这些结果说明同一基因在不同的组织中及同一组织中不同的基因对同一种应激的敏感性不同,这可能与各自功能不同有关.
总之,本研究对H d AK的cDNA序列进行克隆和分析,以血细胞和鳃组织为研究对象,对H d AK基因在高水温、低溶氧、高水温与低溶氧联合应激及副溶血弧菌感染后的表达规律进行测定,结果显示Hd AK参与了高水温、低溶氧等不利环境的适应过程及免疫反应过程,研究结果可为揭示高水温、低溶氧、高水温与低溶氧联合应激及副溶血弧菌感染对杂色鲍的影响机制提供理论依据.
[1] Ellington W R.Phosphocreatine represents a thermodynamic and function improvement over other muscle phosphagens[J].Journal of Experimental Biology,1989, 143:177-194.
[2] 李松伟,李晔,李成华,等.泥蜡(Tegillarca granosa)精氨酸激酶的基因克隆与表达[J].海洋与湖沼,2011,42(1): 119-123.
[3] Suzuki T,Inoue N,Higashi T,et al.Gastropod arginine kinases from cellana grata and Aplysia kurodai[J].Comparative Biochemistry and Physiology.Part B:Biochemistry&Molecular Biology,2000,127(4):505-512.
[4] Shofer S L,Willis J A,Tjeerdema R S.Effects of Hypoxia and toxicant exposure on arginine kinase function as measured by31P-NMR magnetization transfer in living abalone[J].Comparative Biochemistry and Physiology. Part C:Pharmacology,Toxicology and Endocrinology, 1997,117(3):283-289.
[5] Abe H,Hirai S,Okada S.Metabolic responses and arginine kinase expression under hypoxic stress of the kuruma prawn Marsupenaeus japonicus[J].Comparative Biochemistry and Physiology.Part A:Molecular&Integrative Physiology,2007,146(1):40-46.
[6] Astrofsky K M,Roux M M,Klimpel K R,et al.Isolation of differentially expressed genes from white spot virus (WSV)infected Paci fic blue shrimp(Penaeus stylirostris)[J].Archives of Virology,2002(7):1799-1812.
[7] Ma F F,Liu Q H,Guan G K,et al.Arginine kinase of Litopenaeus vannamei involved in white spot syndrome virus infection[J].Gene,2014,539(1):99-106.
[8] Song C W,Cui Z X,Liu Y,et al.Cloning and expression of arginine kinase from a swimming crab Portunus trituberculatus[J].Molecular Biology Reports,2012,39(4): 4879-4888.
[9] 蔡俊鹏,杨洪志.南方杂色鲍苗大规模死亡病原菌的分离鉴定及其回归感染试验[J].海洋湖沼通报,2006,3: 54-59.
[10] Zhang X,Huang Y,Cai X,et al.Identification and expression analysis of immune-related genes linked to Rel/ NF-κB signaling pathway under stresses and bacterial challenge from the small abalone Haliotis diversicolor [J].Fish&Shell fi sh Immunology,2014,41(2): 200-208.
[11] Cai X H,Huang Y T,Zhang X,et al.Cloning,characterization,hypoxia and heat shock response of hypoxia inducible factor-1(HIF-1)from the small abalone Haliotis diversicolor[J].Gene,2014,534(2):256-264.
[12] 黄贻涛,蔡秀红,张子平,等.杂色鲍紫色酸性磷酸醋酶基因克隆及应激下的表达[J].中国水产科学,2013,20 (5):939-949.
[13] 张鑫,黄贻涛,蔡秀红,等.杂色鲍14-3-3ζ基因的克隆及其在应激下的表达[J].水产学报,2014,38(4): 491-501.
[14] 黄贻涛,蔡秀红,张子平,等.杂色鲍同种移植炎症因子1的克隆及其在应激下的表达[J].水产学报,2013,37 (6):830-839.
[15] Huang Y T,Cai X H,Zou Z H,et al.Molecular cloning, characterization and expression analysis of three heat shock responsive genes from Haliotis diversicolor[J]. Fish&Shell fi sh Immunology,2014,36(2):590-599.
[16] 王宝珍,张子平,王艺磊,等.杂色鲍β-1,3-葡聚糖识别蛋庄基因的克隆和表达[J].海洋科学,2011,35(8): 67-75.
[17] 王艺磊,张子平.日本对虾精巢和卵巢全长cDNA文库的构建[J].动物学杂志,2003,38(2):9-13.
[18] 王华兵,徐豫松.家蚕精氨酸激酶基因的克隆、基因结构与表达分析[J].中国农业科学,2006,39(11): 2354-2361.
[19] 苏晓峰,陆国清,程红梅.精氨酸激酶蛋庄及分子生物学的研究进展[J].生物技术通报,2011(4):26-30.
[20] 姚翠莺,冀培丰,孔鹏,等.凡纳滨对虾精氨酸激酶的多克隆抗体制备及组织特异性表达分析[J].水产学报, 2009,33(6):1026-1030.
Cloning and Differential Expression of Arginine Kinase Gene in Haliotis diversicolor under Different Stresses
LI Wen-hui1,ZHANG Xin2,ZHU You-fang1,ZHANG Zi-ping2,WANG Yi-lei2*
(1.Putian Municipal Institute of Fishery Science,Putian 351100,China;2.Key Laboratory of Healthy Mariculture for East China Sea,Ministry of Agriculture,Fisheries College,Jimei University,Xiamen 361021,China)
:The full length cDNA of arginine kinase gene(Hd AK)from Haliotis diversicolor was cloned for the first time by SMART-RACE method.The m RNA expression levels of Hd AK in examined tissues and in gill and haemocytes under thermal stress,hypoxia exposure,thermal plus hypoxia stress and the injection of Vibrio parahaemolyticus were analyzed by real-time quantitative PCR(qRT-PCR).Results showed that the full length c DNA sequence of Hd AK is 1 595 bp and encodes 354 amino acids.The qRT-PCR results showed that Hd AK gene was detected in all examined tissues and with the highest expression level in hepatopancreas(p<0.05).Under different stresses,the m RNA expression levels in either gills or haemocytes of Hd AK gene were signi fi cantly up-regulated in some phases.These results indicated that Hd AK is involved in response to energy metabolism and plays roles in response to thermal,hypoxia stress and the injection of V.parahaemolyticus.These results will facilitate the study of the physiological mechanisms of these stresses on H.diversicolor.
Haliotis diversicolor;arginine kinase gene;thermal stress;hypoxia stress;thermal plus hypoxia stress;Vibrio parahaemolyticus infection
Q 785;S 968.3
A
0438-0479(2015)03-0315-09
10.6043/j.issn.0438-0479.2015.03.004
2014-10-16 录用日期:2015-01-14
国家自然科学基金(41176152);莆田市科技局项目(2014N07);集美大学创新团队基金(2010A001)
*通信作者:ylwang@jmu.edu.cn
李文辉,张鑫,朱友芳,等.杂色鲍精氨酸激酶基因的克隆及其在不同应激条件下的表达[J].厦门大学学报:自然科学版,2015,54(3):315-323.
:Li Wenhui,Zhang Xin,Zhu Youfang,et al.Cloning and differential expression of arginine kinase gene in Haliotis diver
sicolor under different stresses[J].Journal of Xiamen University:Natural Science,2015,54(3):315-323.(in Chinese)
