水貂IFN-α、IFN-β及IFN-γmRNA 荧光定量RT-PCR 检测方法的建立及应用
2015-06-11张海玲鲁荣光孙彦刚马凡舒赵建军薛向红徐淑娟范思宁凌明玉李欣彤闫喜军
王 洋,胡 博,张海玲,鲁荣光,吕 爽,刘 昊,孙彦刚,马凡舒,赵建军,张 蕾,薛向红,史 宁,白 雪,徐淑娟,范思宁,凌明玉,李欣彤,闫喜军*
(1.中国农业科学院特产研究所经济动物疫病研究室,吉林长春130112;2.吉林农业大学动物科学技术学院,吉林长春130118)
干扰素(interferon,IFN)是机体抗病毒感染的主要细胞因子,具有调节机体免疫功能、抗病毒、抗肿瘤等多种作用,是机体防御系统的重要组成部分[1]。根据基因序列、染色体定位和受体特异性可以将IFN 分为3种类型,IFN-α和IFN-β属于Ⅰ型干扰素,是机体抗病毒感染的主要细胞因子,具有抑制病毒复制和间接的防护病毒感染的作用[2-3]。IFN-γ属于Ⅱ型干扰素,又称免疫干扰素,主要由活化的T 细胞和NK 细胞分泌,在病毒感染过程中具有活化免疫细胞的功能,即在免疫调节中发挥重要作用[4-5]。Ⅲ型干扰素是2003年发现的一种新的细胞因子,称为IFN-λ,其抗病毒机制与Ⅰ型IFN 相类似[6]。IFN 是机体内各种抗病毒因子的潜在活化剂,可在较短时间内使机体处于抗病毒状态,使机体对病毒的感染作出快速抵抗[7]。因此,对机体内IFN 进行定量分析,不仅对免疫机制研究、免疫功能分析、疫苗免疫效果评估、器官移植、过敏反应,而且对多种胞内病原感染的诊断等方面都具有极其重要的理论价值和应用价值。实时荧光定量PCR(realtime PCR)具有灵敏度高、特异性好和操作简便等特点,已成为核酸定量检测的常用手段,该技术在犬细胞因子的检测中也有应用[8]。本研究拟建立一种用于检测水貂IFN-α、IFN-β和IFN-γ mRNA 的荧光定量RT-PCR 方法,对水貂IFN-α、IFN-β和IFN-γ等细胞因子的表达水平作出评价,为水貂细胞因子的定量检测提供技术支持。
1 材料与方法
1.1 材料
1.1.1 实验动物和毒种 水貂由中国农业科学院特产研究所实验动物基地提供;水貂肠炎病毒强毒株(SMVP-11)由本课题组分离和保存,病毒效价为5×106TCID50/mL。
1.1.2 主要试剂和仪器 淋巴细胞分离液,天津市灏洋生物制品科技有限责任公司;Axy Prep Multisource Tota RNA Miniprep Kit、Axy Prep DNA Gel Extraction Kit和Axy Prep Plasmid Miniprep Kit,杭州AXYGEN 公司;一步法反转录试剂盒,北京全式金生物技术有限公司;SYBR Premix®Ex TaqTMⅡ和pMD18-T,大 连TaKaRa 公 司;Light Cycler®96荧光定量PCR 仪,罗氏公司。
1.2 方法
1.2.1 引物设计与合成 根据GenBank已发表的MiGAPDH、MiIFN-α、MiIFN-β和MiIFN-γ基因的核苷酸序列(登录号分别为:EF392835.1、EU863613.1、EF581890.1和KJ888148.1)[9],用Primer 5.0软件设计荧光定量PCR特异性引物(表1)。引物由上海生工生物工程技术服务有限公司合成。
1.2.2 淋巴细胞总RNA 提取与cDNA 制备 采集水貂新鲜血液2.5mL置于肝素抗凝管,加入等体积生理盐水。取5mL淋巴细胞分离液加入无菌离心管中,将5mL稀释后的血液缓慢加入分离液的液面上,低速冷冻水平离心机400r/min离心20min。用吸管小心吸取中间层淋巴细胞,加入5mL生理盐水中,充分混匀,400r/min离心20min。弃去上清液,沉淀用5mL生理盐水洗2次,获得淋巴细胞。按RNA 提取试剂盒和一步法反转录试剂盒操作说明获得cDNA,置-20℃保存。

表1 MiGAPDH、MiIFN-α、MiIFN-β和MiIFN-γ引物Table 1 The primers of MiGAPDH,MiIFN-α,MiIFN-βand MiIFN-γ
1.2.3 质粒标准品的制备 以获得的cDNA 为模板,常规PCR 扩增MiGAPDH、MiIFN-α、MiIFN-β和MiIFN-γ基因。反应体系为10×ExTaq buffer 3 μL,dNTP Mixture 1.2 μL,TaKaRa ExTaq 0.6μL,上、下 游 引 物 各0.5 μL,cDNA 2 μL,ddH2O 补足30μL。PCR 扩增程序为:95℃4min;95℃25s,55℃30s,72℃1min,33个循环;72℃延伸7min。15g/L琼脂糖凝胶电泳检测PCR 产物,凝胶回收产物连接于pMD18-T 载体,连接产物转化到Trans 5α感受态细胞,培养并挑取单克隆菌落于液体LB 培养基37℃摇振培养12h,提取质粒经鉴定正确后,紫外线分光光度计测定OD 值,换算成拷贝数并进行倍比稀释。
1.2.4 反应条件的优化 在其他条件不变的情况下,通过比较Ct值、荧光强度及熔解曲线是否有引物二聚体峰来确定最佳退火温度。在优化的退火温度条件下,对引物浓度(5mmol/L~20mmol/L)进行优化。
1.2.5 荧光定量PCR 标准曲线的建立 以10倍梯度稀释已知拷贝数的重组质粒,荧光定量PCR 检测,反应完成后,应用Light Cycler 96 荧光定量PCR 仪自带分析软件自动生成标准曲线。
1.2.6 荧光定量PCR 敏感性、重复性和特异性试验 将101拷贝/μL~107拷贝/μL 的质粒标准品作为模板进行荧光定量PCR 检测,确定反应的灵敏性。将4种质粒标准品稀释成3个浓度梯度,进行组内、组间重复检测,每个稀释度3 个重复。按1.2.2的方法进行总RNA 的提取和cDNA 的合成,用MiGAPDH、MiIFN-α、MiIFN-β 和MiIFN-γ 的各自引物进行荧光定量RT-PCR 检测,并对荧光定量PCR 产物进行熔解曲线分析。
1.2.7 临床样品检测 选取细小病毒抗体阴性水貂18只,随机选取15只水貂,口服接种水貂肠炎病毒(浓度5×106TCID50/mL),10 mL/只。其余3只水貂作为正常对照,口服正常细胞培养液。在不同的攻毒时间采集水貂血液,按照1.2.2进行RNA提取及cDNA 合成。采用建立的检测方法,对Mi-IFN-α、MiIFN-β及MiIFN-γmRNA 相对表达量进行检测,并对结果进行分析。
2 结果
2.1 质粒标准品的制备
由水貂外周血淋巴细胞提取的总RNA 经反转录成cDNA 后,经PCR 扩增,获得MiGAPDH(254 bp)、MiIFN-α(150bp)、MiIFN-β(170bp)和MiIFN-γ(104bp)基因的目的片段(图1),经胶回收、连接、转化、摇菌、提取质粒、酶切及测序鉴定,证实成功构建出带有目标片段的重组质粒,浓度依次为11μg/mL、3.5μg/mL、15.7μg/mL和20μg/mL,换算成拷贝数分别为3.4×1010拷贝/μL、1.12×1010拷贝/μL、5.0×1010拷贝/μL和6.54×1010拷贝/μL。用无菌水进行梯度稀释,制备成101拷贝/μL~107拷贝/μL的标准质粒,OD260nm/OD280nm 介于1.80~1.90 之间。
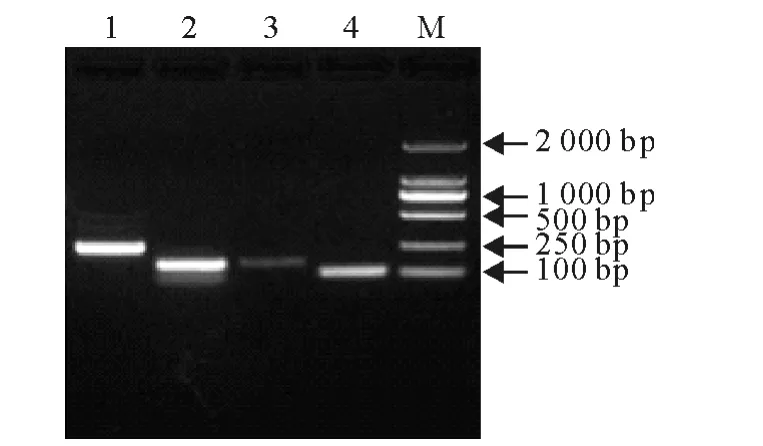
图1 目的片段PCR 产物琼脂糖凝胶电泳图Fig.1 Agarose gel electrophoresis of PCR products of target fragments
2.2 反应条件的优化
根据预试验对反应条件进行优化。退火温度55℃,引物浓度10pmol/μL,有最低的Ct值和最高的荧光强度,并且无二聚体。总体系20μL:SYBR Premix Ex TaqTM Ⅱ10 μL,上、下 游 引 物 各0.5μL,ddH2O 7μL,模板2μL。反应在Light Cycler 96荧光定量PCR 仪中进行,反应程序为:95℃30s;95℃6s,55℃30s,72℃30s,40个循环,每一次循环结束后收集荧光。
2.3 标准曲线的制作
以优化好的条件和程序对质粒标准品进行荧光定量PCR 扩增,经Light Cycler 96 荧光定量PCR软件分析得到MiGAPDH、MiIFN-α、MiIFN-β 和MiIFN-γ的动力学扩增曲线(图2)和标准曲线(图3)。标准曲线的相关系数R2均为1.00,各点间的线性关系良好,扩增效率高,标准曲线符合试验标准。
2.4 敏感性、特异性和重复性试验
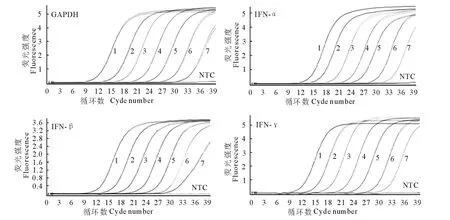
图2 细胞因子检测基因动力学扩增曲线Fig.2 Dynamics curve of MiGAPDH,MiIFN-α,MiIFN-βand MiIFN-γmRNA detection
2.4.1 敏感性试验 结果表明,MiGAPDH、Mi IFN-α、MiIFN-β和MiIFN-γ 的重组质粒拷贝数浓度在101~107拷贝/μL范围内扩增效果良好,最小检出下限为10 拷贝重组质粒/μL,阴性对照组(NTC)无信号。
2.4.2 特异性试验 熔解曲线分析表明(图4),MiGAPDH、MiIFN-α、MiIFN-β和MiIFN-γ的质粒标准品只出现特异性单峰,无非特异性产物和引物二聚体的峰值出现,阴性对照组(NTC)没有杂峰,表明所建立的方法具有较好的特异性。
2.4.3 重复性试验 从MiGAPDH、MiIFN-α、Mi-IFN-β和MiIFN-γ质粒标准品中分别选取3个浓度梯度进行组内和组间重复性试验,结果显示,变异系数(CV)均小于4.5%,表明该方法具有较好的重复性(表2)。
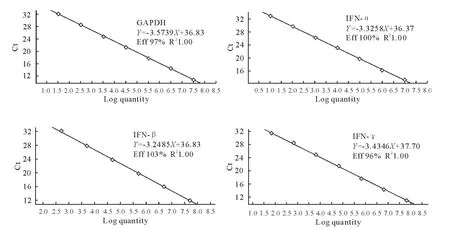
图3 MiGAPDH,MiIFN-α,MiIFN-β和MiIFN-γ实时荧光定量PCR 标准曲线Fig.3 Real-time PCR standard curves of MiGAPDH,MiIFN-α,MiIFN-βand MiIFN-γ
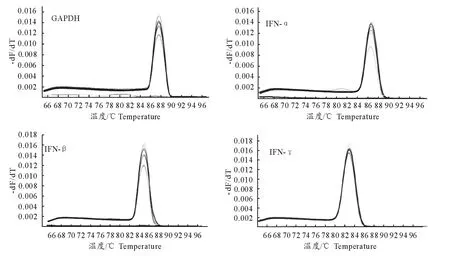
图4 MiGAPDH,MiIFN-α,MiIFN-β和MiIFN-γ实时荧光定量PCR 熔解曲线Fig.4 Real-time PCR melting curves of MiGAPDH,MiIFN-α,MiIFN-βand MiIFN-γ
2.5 临床样品检测
采集水貂感染肠炎病毒后不同时间点的血液样品,检测水貂感染肠炎病毒后不同时间外周血淋巴细胞 中MiIFN-α、MiIFN-β 及MiIFN-γ 的mRNA水平(图5)。结果表明,MiIFN-α、MiIFN-β和Mi-IFN-γmRNA 水平均在感染6d 达到最高峰值,MiIFN-α的量要比MiIFN-β和MiIFN-γ高两个数量级,并且在感染期(4d~6d)MiIFN-αmRNA 水平明显提高,MiIFN-α mRNA 量在2d 为1.73×102拷贝/μL,在6d 时MiIFN-α 相对表达量高达2.38×103拷贝/μL。
3 讨论
干扰素(IFN)是由受到病毒或其他干扰素诱生剂刺激巨噬细胞、淋巴细胞以及体细胞等而产生的一种具有抗病毒、抗肿瘤及免疫调节活性等多方面的生物学功能的糖蛋白[10-11]。科研人员从作用机理、功能结构和医疗作用等方面对干扰素展开了长期深入的研究[12]。随着人用干扰素制剂的广泛使用,兽用干扰素的相关研究也逐渐开展。谢丽君等[13]研究猪繁殖与呼吸综合征病毒(PRRSV)体外感染猪肺泡巨噬细胞(PAMs)对其产生Ⅰ型干扰素mRNA 转录水平的影响,结果PRRSV 可导致PAMs产生的Ⅰ型干扰素mRNA 转录水平降低,从而使宿主细胞及机体非特异性免疫功能受到抑制。Zhang H 等[14]研究证实,MiIFN-α抑制水泡性口炎病毒在WISH 细胞上的生长,并且MiIFN-α2可抑制犬瘟热病毒在Vero细胞上的生长。因此,对干扰素相对表达量的检测,可间接的对动物感染病毒性疾病的状态和机体的免疫活性做出初步评价。
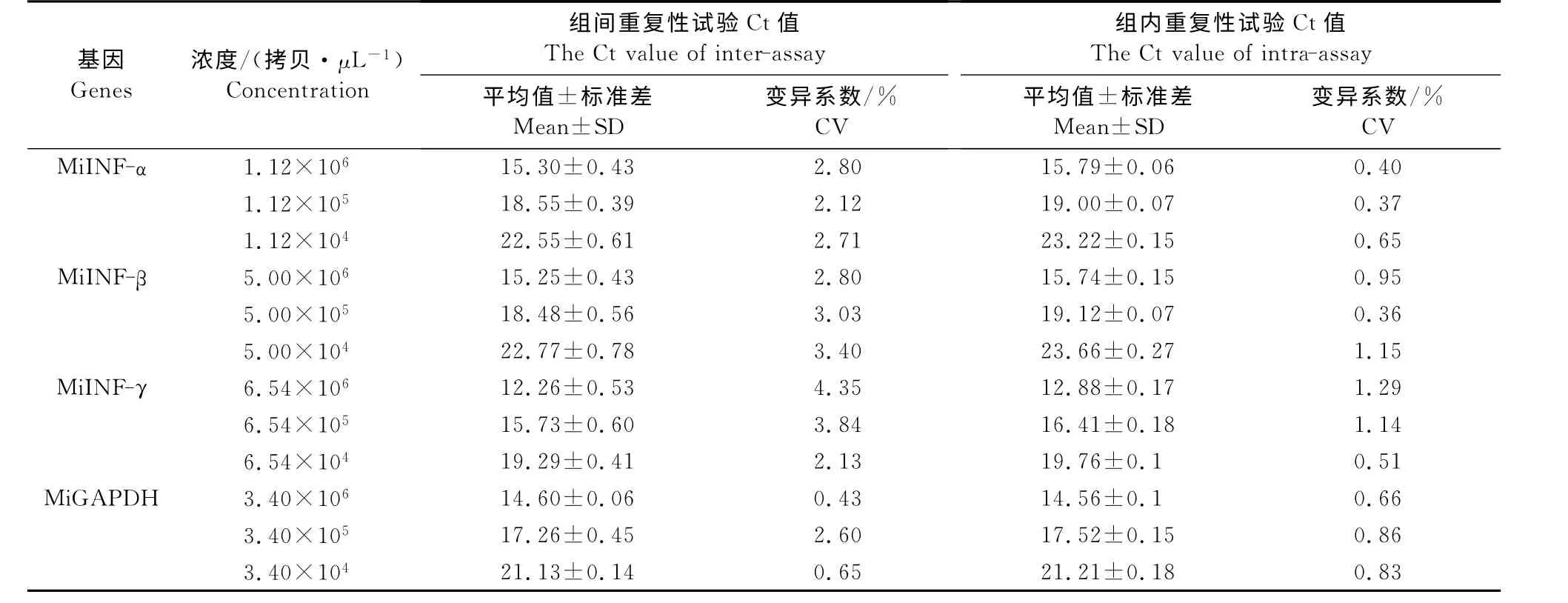
表2 实时荧光定量RT-PCR重复性试验Table 2 Inter-assay and intra-assay reproducibilities of the real-time RT-PCR
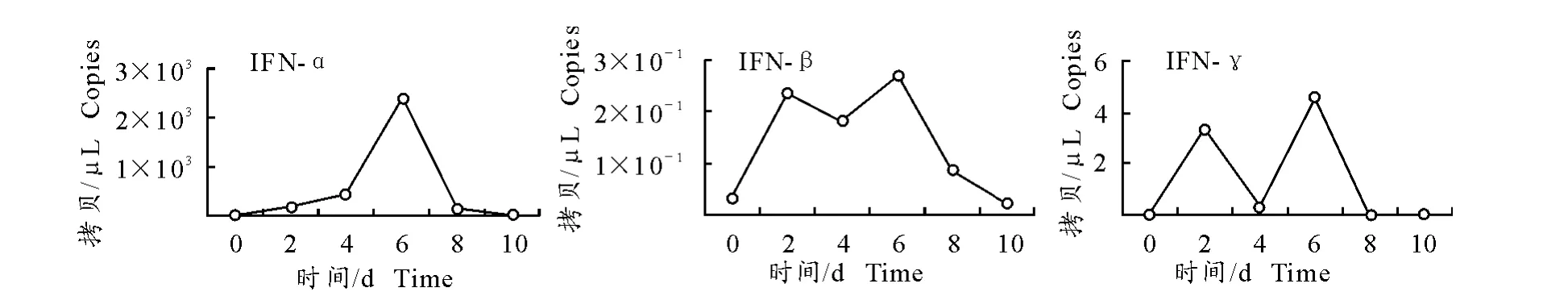
图5 不同时间点MiIFN-α、MiIFN-β和MiIFN-γmRNA 的动态表达Fig.5 Dynamic expression of MiIFN-α,MiIFN-βand MiIFN-γat different times
本研究以GAPDH 基因为内参基因,采用SYBR GreenⅠ荧光定量PCR 检测MiIFN-α、Mi-IFN-β和MiIFN-γ 基因的mRNA 转录水平,建立了检测水貂MiIFN-α、MiIFN-β和MiIFN-γmRNA实时荧光定量RT-PCR 检测方法。结果4 种基因标准曲线的R2均在0.995~1.000之间,扩增效率较高,标准曲线具有良好的线性关系;由熔解曲线可以看出,无非特异性产物和引物二聚体出现,表明所用的扩增4个基因的引物具有较好的特异性;扩增曲线呈现较好的S 型,检出下限为10 拷贝重组质粒/μL,敏感性良好;组内、组间重复性试验显示变异系数均小于4.5%,具有较好的重复性。应用本研究建立的检测方法,对水貂感染肠炎病毒后不同时间外周血淋巴细胞中IFN 的相对变化进行了检测。结果表明,病毒感染后6d3种IFN 的mRNA水平均达到最高峰值,而感染后4d到6d正是水貂肠炎病毒在水貂体内表达量达到峰值的时段[15],表明IFN mRNA 水平的改变与病毒感染有着密切的关系,体现出IFN 直接或间接的对病毒感染的抑制作用[16]。经过检测MiIFN-α相对表达量较MiIFNβ和MiIFN-γ高两个数量级,且在病毒感染期(4d~6d)MiIFN-α相对表达量明显提高,MiIFN-β和MiIFN-γ在病毒感染初期(2d)及感染期(4d~6d)均有两个峰值,综上说明MiIFN 与机体抗病毒感染过程有一定的相关性。
本研究建立的水貂MiIFN-α、MiIFN-β和Mi-IFN-γ3种细胞因子实时荧光定量PCR 检测方法,可以对IFN 在水貂机体内的动态变化水平进行初步评价,也可以为了解水貂病毒性疾病的发生、发展、转归情况及其机体保护性免疫反应动态规律提供支持,为水貂IFN mRNA 的定量分析提供了有效工具。
[1] Baccala R,Welch M J,Gonzalez-Quintial R,et al.Type I interferon is a therapeutic target for virus-induced lethal vascular damage[J].Proceed Nat Acad Sci,2014,111(24):8925-8930.
[2] Ivashkiv L B,Donlin L T.Regulation of type I interferon responses[J].Nat Rev Immunol,2014,14(1):36-49.
[3] Stifter S A,Feng C G.Interfering with immunity:Detrimental role of type I IFNs during infection[J].J Immunol,2015,194(6):2455-2465.
[4] Qiao Y,Giannopoulou E G,Chan C H,et al.Synergistic activation of inflammatory cytokine genes by interferon-γ-induced chromatin remodeling and Toll-like receptor signaling[J].Immunity,2013,39(3):454-469.
[5] Pollard K M,Cauvi D M,Toomey C B,et al.Interferon-γ and systemic autoimmunity[J].Discov Med,2013,16(87):123-131.
[6] Dickensheets H,Sheikh F,Park O,et al.Interferon-lambda(IFN-λ)induces signal transduction and gene expression in human hepatocytes,but not in lymphocytes or monocytes[J].J Leukocyte Biol,2013,93(3):377-385.
[7] Liu S Y,Sanchez D J,Aliyari R,et al.Systematic identification of typeⅠand typeⅡinterferon-induced antiviral factors[J].Proceed Nat Acad Sci,2012,109(11):4239-4244.
[8] 潘福星,王冬冬,曹志伟,等.比格犬IFN-α和IFN-βmRNA SYBR GreenⅠ荧光定量RT-PCR 检测方法的建立[J].中国预防兽医学报,2014,36(4):297-300.
[9] Svitek N,von Messling V.Early cytokine mRNA expression profiles predict morbillivirus disease outcome in ferrets[J].Virology,2007,362(2):404-410.
[10] Sadler A J,Williams B R G.Interferon-inducible antiviral effectors[J].Nat Rev Immunol,2008,8(7):559-568.
[11] Dussurget O,Bierne H,Cossart P.The bacterial pathogen Listeria monocytogenes and the interferon family:type Ⅰ,typeⅡand typeⅢinterferons[J].Front Cell Infect Microbiol,2014,4:1-12.
[12] 王 照,王 磊,李天松,等.动物干扰素的研究进展[J].中国动物检疫,2012,29(2):71-73.
[13] 谢丽君,邵志勇,刘欢欢,等.猪繁殖与呼吸综合征病毒感染对猪肺泡巨噬细胞产生Ⅰ型干扰素mRNA 转录水平的影响[J].中国兽医科学,2014,44(10):1047-1052.
[14] Zhang H,Zhao J,Chai X,et al.Cloning,expression and antiviral activity of mink alpha-interferons[J].BMC Vet Res,2015,11(1):1-8.
[15] Uttenthal A,Larsen S,Lund E,et al.Analysis of experimental mink enteritis virus infection in mink:in situ hybridization,serology,and histopathology[J].J Virol,1990,64(6):2768-2779.
[16] Davidson S,Maini M K,Wack A.Disease-promoting effects of type I interferons in viral,bacterial,and coinfections[J].J Interferon Cytokine Res,2015,35(4):252-264.
