Post-fire regeneration dynamics in whitebark pine(Pinus albicaulis)forests in Wind River and Absaroka Mountains, Wyoming,USA
2015-06-05JenniferKlutschBetsyGoodrichWilliamJacobi
Jennifer G.Klutsch•Betsy A.Goodrich•William R.Jacobi
Post-fire regeneration dynamics in whitebark pine(Pinus albicaulis)forests in Wind River and Absaroka Mountains, Wyoming,USA
Jennifer G.Klutsch1,2•Betsy A.Goodrich1,3•William R.Jacobi1
Whitebark pine(Pinus albicaulis)populations are in decline across the species’range due to historic wildfire exclusion,mountain pine beetle(Dendroctonus ponderosae,MPB)outbreaks,and an invasive fungal pathogen causing the disease white pine blister rust (Cronartium ribicola,WPBR).Despite reliance on standreplacing fires,information on whitebark pine regeneration occurrence is limited and the trajectory of future forests is largely unknown in some areas of the range.Regeneration densities were assessed in burned and adjacent non-burned areas atsix high elevation locations in northwest Wyoming where stand-replacing fires occurred 8–32 years before thesurveys.In these locations at the eastern extent of the species range,we assessed what site factors were associated with regeneration success.Whitebark pine regeneration density was greater and seedlings were older in non-burned compared to burned areas.Within burns,north aspects had more regeneration than south aspects.Potential seed source densities and other species’regeneration were positively related to whitebark pine regeneration densities in burned areas.South facing slopes or grass covered areas may have either delayed or no regeneration of whitebark pine without the help of artificial planting.
Recruitment·Age structure·White pine blister rust·Mountain pine beetle·Restoration
Introduction
Whitebark pine(Pinus albicaulis Engelm.)is an important component of high-elevation forests in western North America,though it has been in recent decline due to a number of factors,such as the introduced pathogen (Cronartium ribicola J.C.Fisch.,WPBR)causing the disease white pine blister rust(Geils et al.2010),mountain pine beetle(Dendroctonus ponderosae Hopkins,MPB) (Gibson et al.2008),and global climate change(Keane etal.1994;Logan and Powell2001;Tomback etal.2001b; Furniss and Renkin 2003;Hamann and Wang 2006;Logan etal.2010).In these vulnerable high-elevation ecosystems, managementand restoration efforts have occurred to buffer the effects of the current tree decline(Keane and Parsons 2010;Waring and Goodrich 2012).Facilitating natural regeneration and planting of whitebark pine are recommended as restoration activities(Scott and McCaughey 2006;Schoettle and Sniezko 2007;Keane and Parsons2010)as regeneration willbe a key factorin sustaining this keystone species.We conducted a study in the southeastern extent of whitebark pine to determine patterns of post-fire whitebark pine regeneration and the site and stand characteristics associated with regeneration.
The maintenance of whitebark pine in these highelevation alpine and subalpine species is importantbecause it is a keystone species that facilitates biodiversity,influences watershed hydrology and performs other ecosystem services(Tomback et al.2001b;Tomback and Achuff 2010).Whitebark pine produce most seed during mast years(Morgan and Bunting 1992;Crone et al.2011)and severalanimalspecies rely on the large seeds as major food sources,such as squirrels(Tamiasciurus spp.),grizzly bears(Ursus arctos horribilis)and birds including Clark’s nutcrackers(Nucifraga columbiana Wilson)(Hutchins and Lanner1982;Tomback 1982;Mattson etal.1992;Mattson and Reinhart 1997).The main form of seed dispersal for whitebark pine is through a dependentrelationship with the Clark’s nutcracker(Tomback 2001).Seed caching sites that result in the establishment of trees usually occurs in open areas,such as those created by recent fires(Hutchins and Lanner 1982;Tomback 1982).The decline of whitebark pine,even in localized mountain ranges,could have cascading ecosystem effects(Ellison et al.2005).
The regeneration ofwhitebark pine in many locations of the upper subalpine zone ofthe northern Rocky Mountains is assumed to be fire-dependent.As an early arrival to newly opened areas,whitebark pine is eventually outcompeted by more shade tolerant tree species without the maintenance of fire(Pfister et al.1977;Arno and Hoff 1990).Historically fire retun intervals in the upper subalpine zone are long(30–300 years)with fire regimes that vary from stand replacing to mixed-severity;fire exclusion is generally nota factor in contributing to the reduction in whitebark pine regeneration in sites with the longest fire return intervals(Romme 1982;Arno 2001;Walsh 2005; Larson et al.2009).Many whitebark pine forests in the Northern Rockies have originated following stand-replacing fires due to the species ability to survive and regenerate in higher proportions than co-existing shade-tolerant species and its dispersal ecology into areas cleared by fire (Arno and Hoff 1990;Keane and Parsons 2010),but regionally specific guidelines on minimum whitebark pine regeneration densities or the site factors that increase densities is limited in the southern portion of the range.
Recommendations for restoration of whitebark pine have emphasized the use of fire to manage competing vegetation and create seed caching sites for Clark’s nutcrackers and increase age heterogeneity(Keane and Arno 1996,2001;Schwandt2006)while otherstudies have noted adequate regeneration in existing stands(Larson and Kipfmueller2010).Mortality of overstory trees from MPB or WPBR may allow the release of existing whitebark regeneration(Larson and Kipfmueller 2010).The aim of burning is to also promote whitebark pine populations that have the age,stand structure and genetic diversity to be resilientin a changing disease,insect,and climate complex (Schwandt2006;Schoettle and Sniezko 2007).
As important as fires are to whitebark pine and species restoration,limited studies have quantified natural whitebark pine regeneration following stand-replacing fires(but see Tomback et al.1993,1995,2001a).Along the southeastern extent of whitebark pine distribution in the central Rocky Mountains specifically on the Wind River and Absaroka Mountain Ranges of Wyoming,there is little knowledge regarding natural regeneration success following stand replacing fires and in intact non-burned forests. These areas are importantas they are the mostsoutheastern populations of whitebark pine and occur in areas where the species co-occurs with limber pine(Pinus flexilis[James]), another member of the five-needle pine group.Almost all projected species distribution models predict a loss of whitebark pine suitable habitat in these southeastern range edges,therefore itis essentialto provide regionally specific managementguidelines outlining the regeneration niche of whitebark pine here(Crookston 2012).Information on regeneration densities post-fire along with site and stand characteristics associated with whitebark pine regeneration can be valuable information that can be utilized in restoration efforts with regional specificity across the species range,as whitebark pine has a wide distribution and occurs across a range of environments(Arno and Hoff 1990;Larson and Kipfmueller 2012).Research questions related to whitebark regeneration and the influence of fire in this area of its range were:(1)Do whitebark pine regeneration densities by height classes(i.e.,proxy for age) vary between burned and non-burned areas and are there years of greater recruitment;(2)Do site and stand factors correlate with whitebark pine regeneration densities and burn status;and(3)What is the incidence and severity of damage agents including WPBR and MPB on whitebark pine regeneration and mature seed source trees in burned and non-burned areas as this may affect the potential for continuing regeneration cycles.
Materials and methods
Study areas
We chose study areas in the southeastern edge of the natural range of whitebark pine in the Central Rocky Mountains in northwestern Wyoming,USA.We used three wildfire areas on the Wind River Mountain Range that extends along a northwest to southeast axis startingnear Jackson,WY and ending in the southeast,near Lander,WY.Three other fires were used on the southern end of the Absaroka Range that starts at the northern terminus of the Wind River Range and continues north into Montana.Regeneration monitoring plots were established in burned and non-burned areas associated with six stand replacing fires that burned 8–32 years previous to sampling in the summer of 2007(Fig.1; Table 1).Fires ranged in elevation from 2670 to 2890 m and their sizes varied(12 to>2000 ha)with three fires encompassing large areas over 1000 ha each(Table 1). Sampled fires were in areas of potential whitebark pine habitat as defined by elevation and surrounding vegetation,and within an hour hike from a road in the Shoshone National Forest,Bridger-Teton National Forest,and Wind River Reservation(Fig.1).
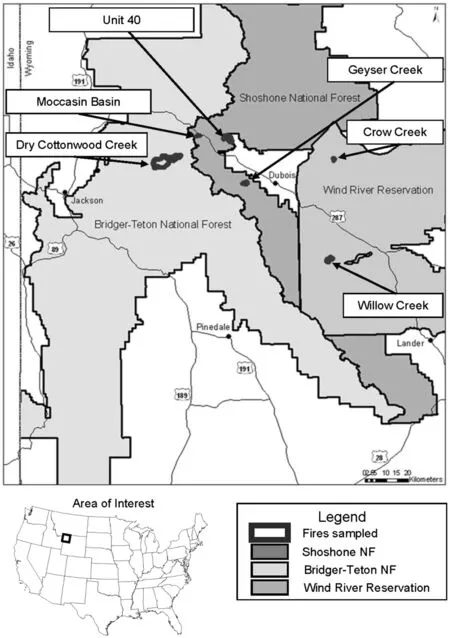
Fig.1 Burned areas sampled in 2007 in the northern Wind River and southern Absaroka Mountain Ranges,WY

Table 1 Study sites sampled in 2007 and condition of non-burned forests around burned areas in the northern Wind River and southern Absaroka Mountain Ranges,WY
Stands in both burned and non-burned areas were mixtures of whitebark pine,Engelmann spruce(Picea engelmannii Parry ex Engelm.),subalpine fir(Abies lasiocarpa [Hook.]Nutt.)and lodgepole pine(Pinus contorta Douglas ex Louden),with proportions of each species differing by site.Limber pine was found at lower elevations than whitebark pine in the study area and was avoided when encountered in two fires by not surveying these lower elevations.On three burned areas the local National Forests had planted either lodgepole pine(Willow Creek,Dry Cottonwood)or whitebark pine(Geyser Creek)with one burned area(Unit40)being planted with both species.We were able to distinguish between human and naturally planted whitebark pine by aging the seedlings by counting whorls.Trees whose age corresponded to dates of planting (based on agency records)were excluded from most analyses.
Sampling strategy
In each burn area,sites with similar aspects,elevation and foresttypes were delimited using GIS.The number of sites within a burned area depended on the uniformity of the burn area in regard to aspect,elevation,foresttype and size of the fire(modified methods from Coop and Schoettle 2009).Sample sites in the non-burned areas were selected from the surrounding forest to match the aspect,elevation and foresttype of the burned sites.In large burn areas up to four sites were identified for sampling to ensure an adequate representation of the conditions within the burn area. Sample sites ranged in size from 12 to 20 ha(Table 1)and the shape varied from elongated rectangles to circular areas.
On each sample site,six potential transects were systematically located with spacing between transects varyingfrom 30 m on smaller sites to 100 m on large sites.Transect lengths were equal to the distance across the fire following the elevational contours and ranged from 200 to 520 m.The six transects defined the sampling frame from which three transects were randomly selected for sampling, without replacement.Two transects were randomly selected from the six transects systematically placed in nonburned areas associated with each sample site.Two transects were used in non-burned areas because these forested areas were uniform and regeneration was not a rare event as in the burned areas.
In designing the survey,transects were treated as clusters and the size of the clusters(the number of plots per transect)was allowed to vary from five to seven depending on transect length.Sample plots were systematically located along each transect.The ends of transects were≥10 m from the nearest non-burned area in burned portions of a sample site and≥10 m from the nearest burned area in non-burned forests.Transects in burned and nonburned areas were monumented at each end with a tagged and painted rebar and a tagged and painted log,tree or stump.
Plot level data
We established 256 plots(each 2 m×40 m)in burned areas and 140 in non-burned forests adjacent to burns (Table 1).On each plot we recorded the aspect,later categorized as north(≥270°and<90°)and south(≥90° and<270°),percent slope,disturbance types(fire,salvage logging,selective cuts,domestic livestock grazing,burn piles,planting ofseedlings),percentage oftop three species of ground cover,percentage of bare ground,litter,and surface area disturbed by pocket gophers(Thomomys talpoides).Plots were categorized as either mesic or xeric; mesic site conditions were based on the presence of Vaccinium scoparium as the majorspecies associated with cool and moist conditions,and xeric conditions were associated with Carex spp.,Juncus spp.,Arnica cordifolia,and bunch grasses(Poaceae family)(Arno 2001).
Tree leveldata
All individual trees were tallied by species in each plot within five height classes(<20,20–99.9,100–199.9, 200–300,≥300 cm).For whitebark pine only,crown class (dominant,codominant,intermediate,overtopped,open grown),seedlings microsite conditions,living or dead status,number of branch whorls(proxy for age)on seedlings<100 cm tall,and abiotic/biotic damage incidence and damage severity(0–9 with a rating of crown or stem killed 0=0–9%affected,1=10–19%affected… 9=90–100%affected)were collected(Kearns and Jacobi 2007;USDA Forest Service 2007).On all whitebark pine in all plots,damage agents were recorded,including both abiotic(frost,sunscald,etc.)and biotic(insect defoliation, diseases including white pine blister rust,foliar fungi,and animal damages including browsing and root damages). Time since mortality by MPB was estimated by bark and branch degradation and whether dead trees were burned in the fire.To determine how many seedlings were in bird seed catches,we recorded how many seedlings<100 cm tall were growing in a cluster(Tomback et al.2001a). Microsites within 20 cm around whitebark pine regeneration were classified as:open,no protection from wind or solar;nearthe base ofa tree orunderlow tree canopy;near downed woody debris,logs,or roots;rocks;and in cracks or holes in trees.
Since tree diameters were notrecorded within plots,the basal area of all tree species along with health class, damage agent incidence,damage severity,whether cones were present and diameter class were assessed in variable radius plots at both ends of each plot using a basal area angle gauge(1.1 metric basal area factor(5 English BAF) in burned areas and 2.3 BAF(10 English BAF)in nonburned areas).The stand basal area and related data were used to assess whether there were relationships between stand characteristics and regeneration.
Seed sources
The non-burned forest outside the fire was sampled to assess whitebark pine seed source at the perimeter of the fire.Sampling was conducted with variable radius plots (2.3 BAF)spaced 80 m apart in non-burned forest 10 m from the perimeter of the fire.The basal area of all tree species along with the health class,damage agent,damage incidence and severity,diameter class and whether whitebark pine cones were present were recorded in each plot.
Seedling age versus height and whorls
A subsample of whitebark pine seedlings were collected in burned and non-burned areas of the Dry Cottonwood fire. Twelve trees under closed canopy and 52 from an open area within the burn were destructively sampled.We determined the pith age of each seedling at ground line by counting rings under 40×magnification.Tree heights and whorls were recorded and a linear regression was used to determine the relationships between tree ages and height, whorl counts(proxy for age),and overstory influences (Bu¨sgen and Munch 1929).
Data analysis
To determine differences in tree densities and vegetation covers between burned and non-burned areas and north and south facing slopes,a mixed model analysis of variance was applied with aspect and burn status as fixed effects, and polygon,transect,and number of plots as random effects(SAS/STAT version 9.1,MIXED procedure).Fires were assumed to be independentreplicates.Firstwe tested for spatial auto-and cross-correlation between plots within fires(p<0.05)by performing spatial proximity weights equal to the inverse of the distance between plots,defining a neighborhood for each fire thatranged from 10 to 45 m. Analysis of spatial correlation was carried out using R 2.6.1(R Development Core Team 2008)with additional functions to calculate and test Moran’s I(Robin Reich, Colorado State University,personalcommunication).Since there was spatial auto-correlation the coordinates associated with each plot were used to fit a spatial power model forthe residuals in the mixed model.In allmodelspredicting whitebark pine density,the following site and stand characteristicsfrom each plotwere tested aspotentialcovariates: elevation,time since fire,percent cover of understory vegetation(total,shrubs,grasses,and V.scoparium),density of non-whitebark pine trees in all size classes(total and individual species),and distance and basal area of nearest whitebark pine potential seed source.Covariates that produced the lowest mode Akaike’s Information Criterion (AIC)score were kept in models.The density of trees and vegetation cover were positively skewed and to meet the assumption ofnormality foranalysisthe tree data were log10transformed and the vegetation data square-root transformed.The back-transformed data are presented which correspond more to the median than to the mean(James zumBrunnen,Colorado State University Franklin A.GreybillStatisticalLaboratory,personalcommunication).Linear regressions and paired t-tests were performed to identify differences in height,whorl count,and tree ring count values,which were not transformed because they met the assumptions of normality and homoscedasticity.Also,the frequency distributions of annualrecruitmentof whitebark pine≤100 cm in height in burned and non-burned within and between fires were tested using a Chi square test.To meetthe assumptions ofa Chisquare testwith greater than five observations per recruitment year,groups of 2 years were used to testsimilarities in recruitmentdistribution over time.To visualize understory vegetation community compositions of burned and non-burned areas,a nonmetric multidimensional scaling(NMDS)was performed using the Bray-Curtis dissimilarity distance measure in R 2.6.1 (R DevelopmentCore Team 2008).
Results
Whitebark pine regeneration density and associated site factors
All analyses were conducted on live whitebark pine regeneration because regeneration(≤200 cm height)was generally healthy(mean±SEM=97±1%).Regeneration densities in height classes>20 cm were significantly lower in burned areas than in non-burned areas (Fig.2).This trend was consistent even with the oldestfire (Geyser Creek),where densities of regeneration in areas burned 32 years prior were approximately half of densities in adjacent non-burned forest(Table 2).The only fire that had greater density of regeneration in burned than nonburned was Willow Creek.
Whitebark pine regeneration densities and recruitment years were quite variable in our study area.More than 25%of the plots lacked whitebark pine presence.Furthermore,yearly recruitmentwas notsynchronous between burned and non-burned areas.The frequency distributions of recruitment per year in burned and non-burned areas were significantly different across fires sampled (χ2=185.0,df=8,p<0.001),except for Moccasin Basin which was not included because of very low density of whitebark pine regeneration(Fig.3).Aspect and burn status were significant predictors for densities of the shortest height classes of whitebark pine(≤200 cm in height,representing younger seedlings)(F-value=4.84, p=0.064).South facing burned slopes had six times lower mean whitebark pine(≤200 cm tall)densities compared to north facing burned slopes(mean±SEM=200±43 stems ha-1for south facing burned areas and 1143±119 stems ha-1for north facing burned areas).Similarly,there were eightand four times less whitebark pine≤200 cm tall on south facing burned slopes compared to north nonburned slopes(mean±SEM=1578±205 stems ha-1) and south facing non-burned slopes(773±122 stems ha-1),respectively.The percentcoverof V.scoparium was a positive covariate for this model.
Similarly,the density of whitebark pine in only burned areas was greater on north aspects compared to south aspects for the two shortest height classes(whitebark pine≤20 and>20–100 cm)(Fig.4).Positive covariates in the models for the height classes≤100 cm in height in burned areas were:basal area of nearest whitebark pine potential seed source,percentcover of shrubs,number of years since the fire,and density oflodgepole pine(seedlings to mature trees).Percent cover of grass and minimum distance to potential whitebark pine seed source were negative covariates.
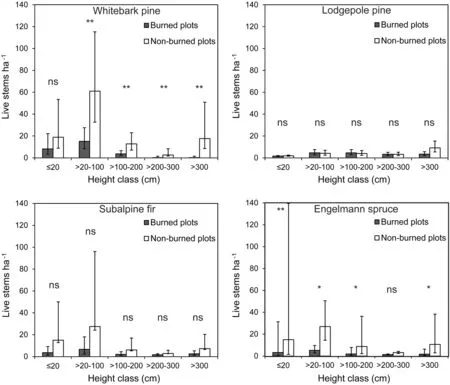
Fig.2 Density of whitebark pine,lodgepole pine,subalpine fir,and Engelmann spruce(back-transformed from log10)live trees by height class in burned and non-burned areas in the northern Wind River and southern Absaroka Mountain Ranges,WY.Values are adjusted for aspect.Error bars are±half LSD.Within height class,ns denotes not significant,*denotes p<0.10,and**denotes p<0.05
With site and stand characteristics such as the cover of V.scoparium and measures of overstory whitebark pine stand characteristics being associated with whitebark pine regeneration,many other site and stand characteristics that were measured did not help explain the variability in whitebark pine densities.For example,the cover of bare ground and litter,and the presence of harvesting,domestic grazing,salvage logging,and pocketgophertunnels did not correlate with regeneration.In fires planted with lodgepole pine,the density of whitebark pine≤200 cm tall was not different in burned areas planted with lodgepole pine compared to burned areas not planted with lodgepole pine (mean±SEM=977±118 stems ha-1).
High percentages of whitebark pine regeneration in burned areas(81%)and in non-burned areas(48%)were found in microsites created by downed woody debris,logs, and roots(Table 3).In the burned areas,there were lower proportions of whitebark pine regeneration under the protection of tree bases or under low tree canopies compared to these same microsites in non-burned areas.Microsites categorized as under protection of rocks and no protection had equaldensities of whitebark pine regeneration between burned and non-burned areas.
Whitebark pine regeneration ages and clusters
With increasing height,there was a greaterage discrepancy of whitebark pine regeneration between burned and nonburned areas.Regeneration below 20 cm talldid notdiffer in age between burned and non-burned areas(mean±-SEM=4.9±0.2 years and 6.9±0.4 years,respectively) (n=177 plots,p=0.177)based on counts of branch whorls.However,regeneration>20–100 cm tall was younger in burned areas(mean±SEM=10.2±0.3-years)compared to non-burned areas(18.8±0.8 years) (n=215 plots,p=0.004).Therefore,heightclass did not accurately represent age class for whitebark pine regeneration when comparing burned and non-burned areas.Although there was a strong positive linear relationship between whorl and tree ring counts for sub-sampled whitebark pine regeneration,whorl counts underestimated tree ring count age by half(Fig.5).Whorl counting generally underestimated the age compared to rings with an average difference of 13.8±1.2 years(t-value=11.74, p<0.001).The heightof sub-sampled regeneration ranged from 7 to 90 cm and averaged 42 cm(SEM=3).Finally, whitebark pine were found growing in clusters of 2–10 trees,with an average of 3.1 stems per cluster (SEM=0.1).
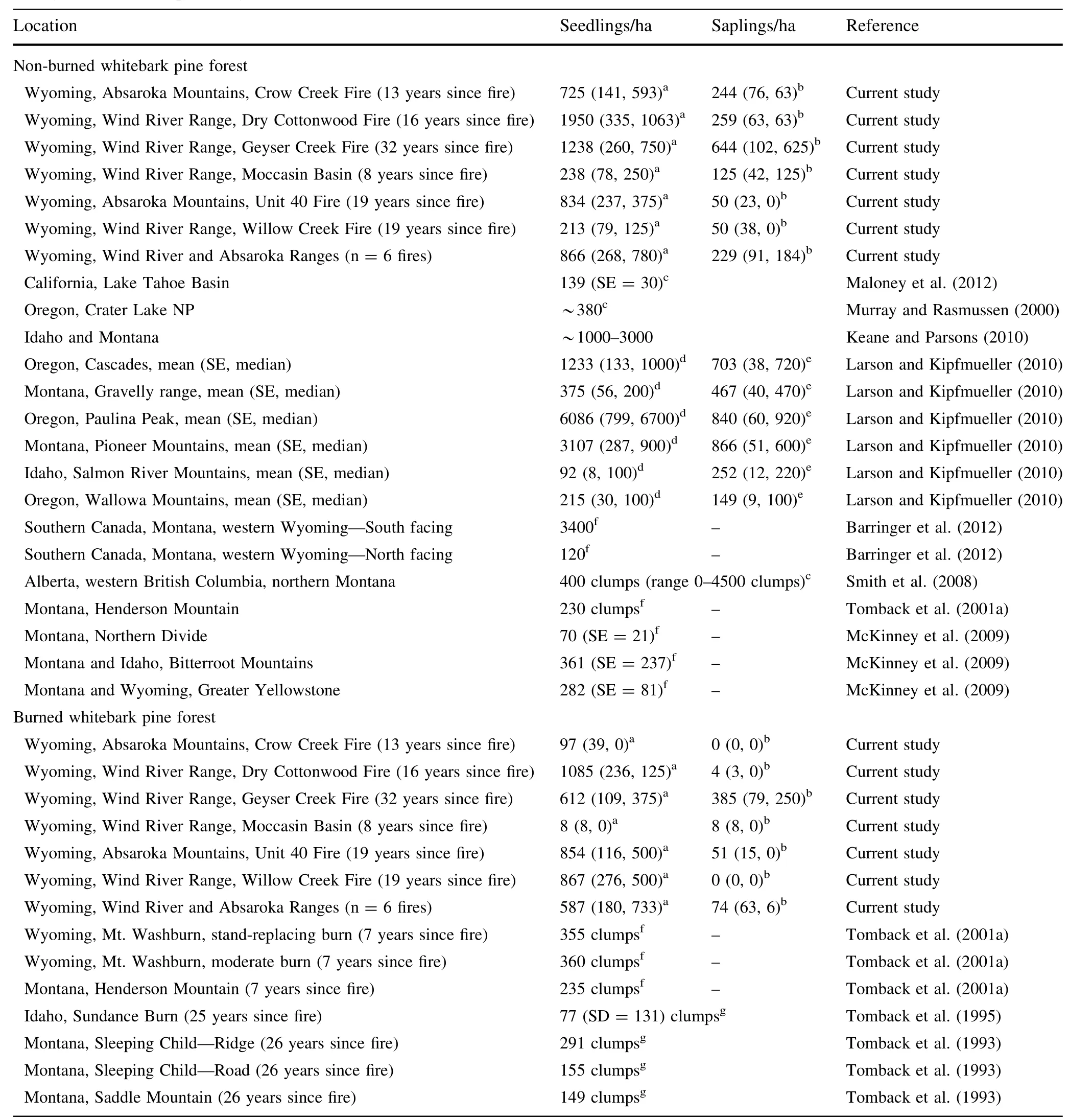
Table 2 Published densities of whitebark pine seedlings and saplings[mean(standard error,median)]in non-burned areas and burned areas throughout whitebark pine range
Disturbance and damage agents on whitebark pine
Animal damage,WPBR infections and MPB mortality were the most common damages to whitebark pine regeneration and overstory trees(Tables 1,4).Browsing or rubbing from animals thatcaused the death of branches or tops of trees were the most common damages on regeneration in both burned(7%)and non-burned areas(12%) (Table 4).Cankers from WPBR occurred on seedlings in allheightclasses exceptforregeneration<20 cm in height and was more prevalent in taller regeneration,but incidence was low(<1%of total density).There was no WPBR or MPB activity on overstory trees in the burned areas.In non-burned areas,the incidence of live whitebark pine(with a measurable DBH)infected with WPBR averaged 3%(range 0–7%),with low average severity (16%),exceptfor the Unit40 area where 30%of crowns or stems on infected trees were infected(Table 1). Mountain pine beetle had caused mortality in the whitebark pine before and after the fires at every burn except Moccasin Basin.Mortality from MPB that occurred before 2000 ranged from 0 to 27%of the susceptible(DBH>20 cm)whitebark pine.Extensive recent MPB-caused mortality of the whitebark pine ranging from 1 to 75%of the stems has occurred since the fires.
Other regenerating species and vegetation
In addition to whitebark pine,other regenerating tree species were subalpine fir,Engelmann spruce,lodgepole pine and Douglas-fir(Pseudotsuga menziesii[Mirb.]Franco.). Densities were generally higher in non-burned areas compared to burned areas for Engelmann spruce butwere equal between burn classes for subalpine fir and lodgepole pine (Fig.2).

Fig.4 Density of(back-transformed log10)live whitebark pine regeneration in only burned areas on north and south aspects in the northern Wind River and southern Absaroka Mountain Ranges,WY. Trees potentially planted in the Geyser Creek and Unit40 fires are not included.Error bars are±half LSD.Within heightclass,ns denotes not significant,*denotes p<0.10,and**denotes p<0.05.aCovariates:trees ha-1of allspecies(seedlings to mature trees)other than whitebark pine,basal area of nearest whitebark pine potential seed source,and percent cover of shrubs.bCovariates:years since fire,density of lodgepole pine(seedlings to mature trees),percent cover of grass,and minimum distance to potential whitebark pine seed source.cCovariate:years since fire
Along with tree regeneration composition and density differencesbetween burned and non-burned areas,there was a clear separation of the understory vegetation community by burn status(Fig.6).Outofthe 47 plantspecies orgroups ofspecies(e.g.,totalgrasses[Poaceae family])recorded,the most influential vegetation types were the cover of total grasses and lupine species(Lupinus spp.),which were associated with burned areas,and grouse whortleberry(V. scoparium)and heartleaf arnica(Arnica cordifolia),which were associated with non-burned areas.From the NMDS ordination analysis,the cover of grass had the maximum correlation with the configuration ofburned and non-burned plots(r=0.79,p<0.05).The coverofgrasswasfive times greater in burned areas than non-burned areas(back-transformed mean±SEM=26.2±0.1%in burned and 5.5±0.1%in non-burned,p=0.001).In the ordination, grass cover was associated with plots with low densities of whitebark pine seedlings.The vegetation type associated with high densities of whitebark pine regeneration was the percent cover of V.scoparium.The percent cover of V. scoparium was 28 times greater in non-burned than burned areas(back-transformed mean±SEM=0.1±0.2%in burned vs.2.8±0.2%in non-burned,p=0.020).Furthermore,burned and non-burned areashad differentamount offorestfloorthatwas bare,disturbed by pocketgophersand covered by litter.There was two times more cover of bare ground in burned areas than non-burned areas(back-transformed mean±SEM=22.4±0.2%in burned and 11.7±0.2%in non-burned,p=0.040).Conversely,there was greater cover of litter in non-burned areas compared to burned(back-transformed mean±SEM=0.2±0.2%in burned and 17.9±0.2%in non-burned,p=0.001).Finally,percentcoverofpocketgopherdisturbance was seven timesgreaterin burned areascompared to non-burned(backtransformed mean±SEM=2.8±0.3%in burned and 0.4±0.3%in non-burned,p=0.021).
Discussion
Whitebark pines have a wide geographic distribution under a diverse range of ecological conditions.Due to the variability in habitats in which whitebark pine occur,the particular conditions associated with whitebark pine regeneration may also vary across its range(Larson and Kipfmueller 2012).Our data contribute to the understanding of regionally specific regeneration dynamics on the southeastern extent of whitebark pine distribution and illustrates the variability and delay in regeneration recruitment after fire.The stand-replacing fires that we sampled were variable in the promotion of successful whitebark pine regeneration in this area of the species range,at least in the post-fire timeframe thatwe studied.
Whitebark pine regeneration density varies between burned and non-burned stands
In the stands studied in the high-elevation forests in northwest Wyoming,fire has altered densities of whitebark pine regeneration as post-fire densities were stilllowerthan adjacent non-burned areas.The fires we assessed that had occurred 8–32 years previous were around the range recommended to adequately assess whitebark pine recruitment after disturbance(10–20 years post-treatment)(Keane and Parsons 2010).However,a lag time of up to 40 years in upper subalpine areas depending on site and disturbance severities has even been suggested for whitebark pine regeneration establishment(Arno and Hoff 1990).Our data showed delayed regeneration establishment with substantial numbers of sapling sized trees(100–300 cm)only occurring in the oldestfire we sampled(32 years since fire; Geyser Creek)(Table 2).The range of densities in the currentstudy extend beyond ranges provided in previously published research,potentially due to a wide range of time since fire(Table 2 and references therein).
To identify whether we could detect years of increased whitebark pine recruitment due to factors such as mast years of cone production or favorable periods of moisture (Morgan and Bunting 1992;Crone et al.2011),we compared annual recruitment in burned and adjacent nonburned areas for each fire(Fig.3).There was a lack ofwhitebark pine recruitment synchrony(based on whorl counts as a proxy for age)between burned and non-burned areas.A combination of factors,such as fluctuations in predation pressure,seedling survival,length of seed dormancy,amountand timing ofprecipitation,and population of Clark’s nutcrackers along with the sampling error for age due to counting whorls may affect yearly seedling recruitment estimates(Tomback et al.2001a;Lorenz et al. 2008).

Table 3 Percentoflive whitebark pine regeneration≤100 cm in heightby microsite location in the northern Wind River and southern Absaroka Mountain Ranges,WY
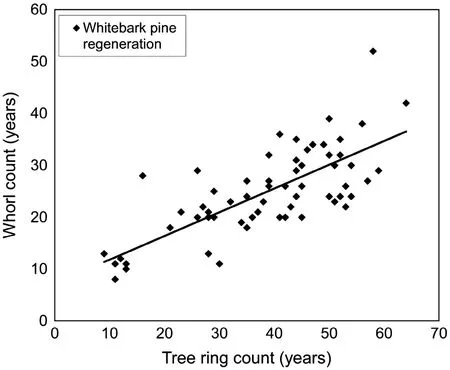
Fig.5 Relationships between whorl counts and tree rings(assessed by dendrochronology)of whitebark pine live regeneration subsamples(n=68)collected at the Dry Cottonwood fire in Wyoming. Regression equation:(Whorl)=(0.4573*Tree ring)+7.2403;F value=72.08,p value<0.0001,R2=0.537
Site and stand factors associated with whitebark pine regeneration
In our study area,densities of whitebark pine were not previously quantified in either burned or non-burned areas. In addition,site and stand conditions related to successfulregeneration were notknown.Densities of regeneration in the burned area varied by fire and were related to specific site and stand characteristics,such as aspect,non-burned tree species composition,vegetation type,and basalarea of potential seed sources.Southerly aspects of these standreplacing fires had the lowest density of whitebark regeneration which would be expected in these mountain ranges where summer precipitation is limited.The relatively cooler and wetter site conditions on north facing slopes along with the success of non-whitebark pine regeneration were positively related to the density of whitebark pine regeneration in burned areas.In western Montana,Tomback and others(1993,2001a,b)also found greater postfire whitebark pine regeneration densities on north-facing aspects and mesic sites.In contrast,Scott and McCaughey (2006)list the warmer aspects exposed to more sun and wind as those that will be more historically dominated by whitebark pine due to less competition with other shadetolerant conifers.We found that the success of other tree species regeneration also correlate with whitebark pine regeneration success.To improve success of artificially planted whitebark pine in burned areas,management recommendations for restoration include the need to remove competing vegetation around trees(HSJ Kearns pers. comm.).There was an association of whitebark pine regeneration with the cover of V.scoparium,which was also shown by Tomback and others(1993).This association may be due to a facultative nurse plantrelationship and the involvement of the potential of shared beneficial mycorrhizal fungi(Perkins 2004).Furthermore,V.scoparium, along with grasses,A.cordifolia,and Lupinus spp.were the most influential vegetation types that defined post-fire community differences in burned and non-burned areas with more whitebark pine regeneration associated with non-burned areas,which were also defined by greater cover of V.scoparium and less cover of grasses.
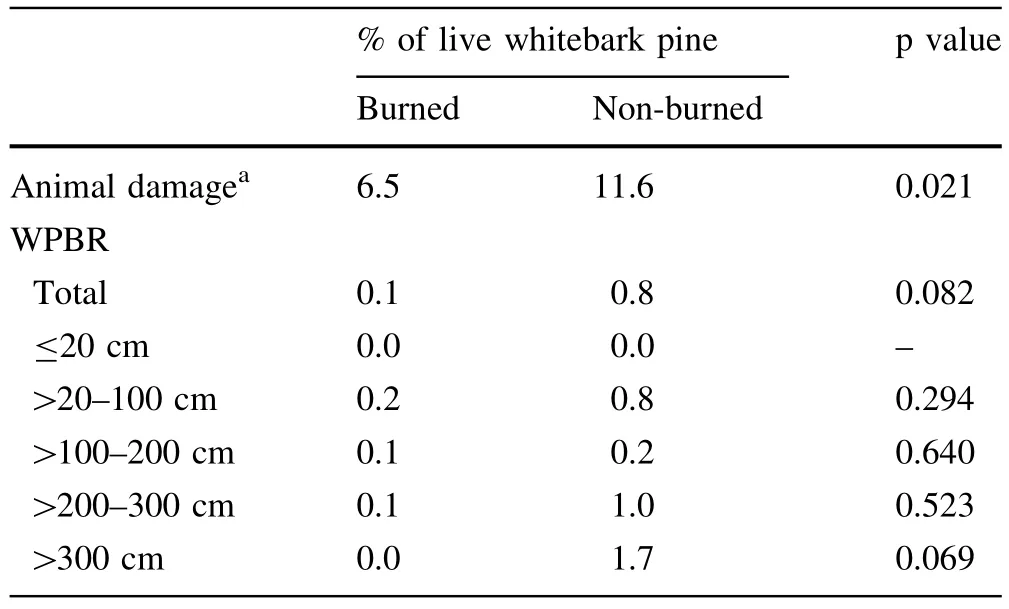
Table 4 Percent affected of whitebark pine regeneration with damages in burned and non-burned areas on whitebark pine in the northern Wind River and southern Absaroka Mountain Ranges,WY

Fig.6 Relationship between understory vegetation community, whitebark pine seedling density,and burn status of plot.Percent cover of all vegetation species were used in the non-metric multidimensional scaling with Bray-Curtis distance ordination analysis (stress=0.26).Point size was scaled to density of whitebark pine seeding density(height<100 cm),which ranged from 0 to 9500 stems ha-1.Only the top four mostinfluentialvariables are displayed as vectors.Vegetation common and scientific names are as follows: grouse whortleberry(Vaccinium scoparium),heartleaf arnica(Arnica cordifolia),allgrasses(Poaceae family),and lupine species(Lupinus spp.)
Mature whitebark pine density and cone production have been shown to be positively associated with the probability of seed dispersal(McKinney et al.2009).In unburned forest in our study area,we would expect successfulseed dispersalas the basalarea of mature whitebark pine(3–7 m2ha-1)was within and above the density range found by Barringer et al.(2012)to have observed Clark’s nutcrackers visitations(0.04–3.23 m2ha-1).Although we found greater densities of whitebark pine regeneration nearerto potentialseed sources,the long distance dispersal of seeds by Clark’s nutcrackers can range from a few meters to severalkilometers(Vander Walland Balda 1977; Tomback 1978;Hutchins and Lanner 1982;Richardson et al.2002;Lorenz and Sullivan 2009).
The microsite conditions of cached whitebark pine seeds in burned and non-burned areas were often associated logs and bases of trees,which act to alter the moisture and temperature of a site to favor regeneration growth.The prevalence of these potential microsites could differ between burned and non-burned areas,for example,tree canopy cover may be lower and amountof downed woody material may be greater in burned areas.Furthermore,the ability to relocate seed caches by Clark’s nutcrackers from these microsites may also differ between burned and nonburned areas,which could also result in differences in regeneration density.Tomback et al.(1993)found the greatest occurrence of whitebark pine regeneration in burned areas were near smallwood pieces,fallen trees and branches.Planting whitebark seedlings atvarying densities in order to use local,naturalmicrosites has been noted as a common practice in restoration planting(Scott and McCaughey 2006;Waring and Goodrich 2012).
Along with altering densities,the stand-replacing fires we surveyed also altered the age structure of regenerating whitebark pine,with regeneration in burned areas growing more rapidly than in the adjacentnon-burned areas.In this study,ages of whitebark pine regeneration in non-burned areas were up to 59%older than those in burns.Furthermore,the growth rate for whitebark pine regeneration>20–100 cm was 1.6 times greater in burned compared to non-burned areas.Because of the paired design comparing similar site characteristics in burned versus adjacent nonburned areas,plausible explanations for differences in growth are changes in overstory stand structure due to fire and the effectthese changes have on abiotic factors such as snow retention and moisture regimes.Due to slow growth rates of older trees and relative shade intolerance,suppressed whitebark pine regeneration in the non-burned areas may have a lower potential of becoming mature overstory trees than the younger and faster growing trees in the burned areas(Pfister et al.1977;Arno and Hoff 1990; Greene et al.1999;Niinemets 2006),so the density disparity between burn status may lower by the time trees reach reproductive ages.However,Keane and others (2007)found thatolderand largerwhitebark pine advancedregeneration had the largestgrowth response after removal of competing vegetation.There is evidence that suggests advanced regeneration may release following canopy gap creations from MPB or WPBR-induced overstory mortality in areas north and west of our research sites(Larson and Kipfmueller2010).Although initialgrowth and survivalof whitebark and limber pine seedlings are greater in areas under some canopy cover compared to open conditions (Maher etal.2005;Casper 2012),the release of these trees is uncertain and more research is needed on the effect of ontogeny and site characteristics on release potential.
Damage agents on whitebark pine regeneration and seed source trees
The incidence of animal damage on seedlings in nonburned(9%)and burned(2%)areas was low and there has been some success with techniques for protecting planted seedlings from herbivores(Smith et al.2011), which may be usefulfor natural regeneration.After animal damage,the damage agent that had the second highest incidence on regeneration was WPBR.The incidence of WPBR on regeneration within burns was low(<1%)butit is notknown whether itmay threaten regeneration density in burned areas in the future.Incidence of rust on taller regeneration in the non-burned stands was high enough (2%)that it may impact whether there is sufficient regeneration surviving to maturity should the overstory trees be removed.The disease pressure on adult whitebark pine is low<3%and seems to be lower(<1%)than other regions on regeneration,as incidence on regeneration ranged from 25 to 29%in southern Canadian Rocky Mountains and northern Idaho(Tomback etal.1995;Smith et al.2008)to<1%in six mountains ranges in Montana, Idaho and Oregon(Larson and Kipfmueller 2010).Recently,Cleaver(2014)surveyed limber pine in the same region as the current study and found the incidence of WPBR on limber pine much greater than on whitebark pine in the currentstudy(23 vs.<3%,respectively).White pine blister rustmay stay atthe presentlow incidence in mature and regenerating whitebark pine,or increase in incidence as ithas done in stands directly to the southeast,also on the Wind River Mountain Range(Kearns and Jacobi 2007). Anotherhindrance to successfulnaturalregeneration in our study area was the ongoing MPB outbreak(Gibson et al. 2008).The mortality of cone-bearing whitebark pine could hinder the successful establishment of natural regeneration in more recentwildfires,such as the Purdy(2006)and Hard Scrabble(2007)fires on the Bridger-Teton and Shoshone National Forests.An increase in WPBR incidence on mature trees and the current MPB outbreak may together reduce seed source density,lower reproductive efforts and interupt seed dispersal to burned areas(Tomback et al. 1995;Tomback and Kendall 2001;McKinney and Tomback 2007;McKinney et al.2009;Barringer et al.2012; Maloney etal.2012).Also,the genetic diversity,hence the resource for potential resistance against WPBR,may be greatly reduced due to the extensive overstory mortality caused by MPB(Schoettle and Sniezko 2007).
Management implications
The Unit40,Willow Creek,and Dry Cottonwood fires were planted at least 6 years prior to the survey with lodgepole pine,though the success ofthese plantings were differentfor each fire and within each fire(E Jungck,pers.comm.).The planting of lodgepole pine in three of the six fires has the potentialto negatively affectthe density of whitebark pine regeneration by eitherdeterring caching ofseedsby birds,by competition,or even by the complex interaction with ectomycorrhizae(Mohatt et al.2008;McCaughey et al.2009). However,there was not a difference in natural whitebark pine regeneration in burned areas notplanted orplanted with lodgepole pine.In the fires interplanted with whitebark pine (Geyser Creek and Unit 40),interplanting in burned areas appears to have been initially successfulas we found lower densities of whitebark pine in areas not interplanted.Furthermore,Izlar(2007)found that the first year survival of planted whitebark pine seedlingsto be higherin burned areas than non-burned areas and positively associated with beneficial ectomycorrhizae,which were found in areas of natural whitebark pine forest type.From these data,the planting of trees after a fire may not deter the natural whitebark pine regeneration and planted and naturalwhitebark pine seedlings may actually benefit from each other through mechanisms such as interactions with beneficial ectomychorrizae.It is generally recommended to plant whitebark pine away from lodgepole pine as lodgepole pine has a faster growth rate and produces seeds when younger than whitebark pine(McCaughey etal.2009).The successof naturalregeneration and artificialinterplanting ofwhitebark pine in areas previously planted with otherspecies should be further researched,as the shade produced by lodgepole pine may help ameliorate dry conditions and promote initial establishment of whitebark pine on south-facing slopes (Arno and Hoff1990).
Recommendations for restoration of seral communities of whitebark pine in dry and exposed sites in previously burned areas,such as sites found in the Wind River Mountain region of Wyoming,include planting in areas that have V.scoparium and do not have extensive grasses (McCaughey et al.2009).Furthermore,planting is recommended in burned areas nextto microsites and in areas that are north facing where there are relatively wet,cool, and sheltered conditions.Promoting and maintaining mature whitebark pine over time in these seral communitiesmay also require removing competing tree species and grasses(Keane and Arno 1996;McCaughey et al.2009). Interplanting whitebark with competing advanced regeneration may be useful to initial establishment,but co-occuring species may also have to be removed.Our research provides regionally specific density estimates in burned and adjacentnon-burned areas and contributes information on whitebark pine regeneration ecology in the southeastern extent of its range.This information can be used by managers as indications of where planting of whitebark pine may be required to restore whitebark pine communities.
AcknowledgmentsWe greatly appreciate the help and invaluable knowledge of our cooperators,E.Jungck,Shoshone NF,E.Davy, Bridger-Teton NF,and E.Rhodenbaugh,Wind River Reservation. We thank J.zumBrunnen and R.Reich for statistical assistance;C. Chavis,K.Houston,E.Winthers,D.West,and J.Ellenwood for data assistance;and field assistance from J.Morrow,J.Egan,and T.Graus. We appreciate the review of this manuscriptby J.Coop,H.Kearns,E. Jungck,and J.Schwandt.We appreciate the anonymous reviewers who helped to improve this manuscript.
Arno SF(2001)Community types and naturaldisturbance processes. In:Tomback DF,Arno SF,Keane RE(eds)Whitebark pine communities:ecology and restoration.Island Press,Washington, pp 76–88
Arno SF,Hoff R(1990)Pinus albicaulis Engelm.Whitebark pine.In: R.M.Burns,B.H.Honkala,(tech.coords.),Silvics of North America.USDA Forest Service,Agricultural Handbook 654, Washington,pp.268–279
Barringer LE,Tomback DF,Wunder MB,McKinner ST(2012) Whitebark pine stand condition,tree abundance,and cone production as predictors of visitation by Clark’s nutcracker. PLoS One 7:e37663.doi:10.1371/journal.pone.0037663
Bu¨sgen M,Munch E(1929)Life and structure of forest trees(trans: Thomson T)3rd edn.Wiley,New York,p 436
Casper AM(2012)Restoration planting options for Pinus flexilis James in the southern Rocky Mountains.MS Thesis,Colorado State University,Fort Collins
Cleaver CM(2014)Limber pine health in the southern and central Rocky Mountains.MS Thesis,Colorado State University,Fort Collins
Coop JD,Schoettle AW(2009)Regeneration of Rocky Mountain bristlecone pine(Pinus aristata)and limber pine(Pinus flexilis) three decades after stand-replacing fires.For Ecol Manag 157:893–903
Crone EE,McIntire EJB,Brodie J(2011)Whatdefines mastseeding? Spatio-temporal patterns of cone production by whitebark pine. J Ecol 99:438–444
Crookston NL(2012)Research on forest climate change:potential effects of global warming on forests and plant climate relationships in Western North America and Mexico.In:Climate data and predictions.USDA Forest Service,Rocky Mountain Research Station,Moscow Forestry Sciences Laboratory,Moscow.<http:// forest.moscowfsl.wsu.edu/climate/>.Accessed 4 April2014
Ellison AM,Bank MS,Clinton BD,Colburn EA,Elliott K,Ford CR, Foster DR,Kloeppe BD,Knoepp JD,Lovett GM,Orwig DA, Rodenhouse NL,Sobczak WV,Stinson KA,Stone JK,Swan CM,Thompson J,Von Holle B,Webster JR(2005)Loss of foundation species:consequences forthe structure and dynamics of forested ecosystems.Front Ecol Environ 3(9):479–486
Furniss MM,Renkin R(2003)Forest entomology in Yellowstone National Park,1923–1957:a time of discovery and learning to let live.Am Entomol 49(4):198–209
Geils BW,Hummer KE,Hunt RS(2010)White pines,Ribes,and blister rust:a review and synthesis.For Pathol 40:147–185
Gibson K,Skov K,Kegley S,Jorgensen C,Smith S,Witcosky J (2008)Mountain pine beetle impacts in high elevation fiveneedle pines:current trends and challenges.R1-08-020.USDA Forest Service,Forest Health Protection,Missoula
Greene DF,Zasada JC,Sirois L,Kneeshaw D,Morin H,Charron I, Simard M-J(1999)A review of the regeneration dynamics of North American boreal forest tree species.Can J For Res 29:824–839
Hamann A,Wang T(2006)Potential effects of climate change on ecosystem and tree species distribution in British Columbia. Ecology 87(11):2773–2786
Hutchins HE,Lanner RM(1982)The central role of Clark’s nutcracker in the dispersal and establishment of whitebark pine. Oecologia 55:192–201
Izlar DK(2007)Assessment of whitebark pine seedling survival for Rocky Mountain plantings.M.S.thesis,College of Forestry and Conservation,University of Montana,Missoula,76 p
Keane RE,Arno SF(1996)Whitebark pine(Pinus albicaulis)ecosystem restoration in western Montana.In:Arno SF,Hardy CC(eds),The use of fire in forest restoration,Society of Ecosystem Restoration. USDA Forest Service,Intermountain Research Station,General Technical Report INT-GTR-341,Ogden,pp 51–54
Keane RE,Arno SF(2001)Restoration concepts and techniques.In: Tomback DF,Arno SF,Keane RE(eds)Whitebark pine communities:ecology and restoration.Island Press,Washington, pp 367–400
Keane RE,Parsons RA(2010)Restoring whitebark pine forests ofthe northern Rocky Mountains,USA.Ecol Restor 28:56–70
Keane RE,Morgan P,Menakis JP(1994)Landscape assessment of the decline of whitebark pine(Pinus albicaulis)in the Bob Marshall Wilderness Complex,Montana,USA.Northwest Sci 68:213–229
Keane RE,Gray KL,Dickinson LJ(2007)Whitebark pine diameter growth response to removal of competition.USDA Forest Service Research Note RMRS-RN-32
Kearns HSJ,Jacobi WR(2007)The distribution and incidence of white pine blister rust in centraland southeastern Wyoming and northern Colorado.Can J For Res 37:1–11
Larson ER,Kipfmueller KF(2010)Patterns in whitebark pine regeneration and their relationships to biophysical site characteristics in southwestern Montana,central Idaho,and Oregon, USA.Can J For Res 40:476–487
Larson ER,Kipfmueller KF(2012)Ecological disaster or the limits of observation?Reconciling modern declines with the long-term dynamics of whitebark pine communities.Geogr Compass 6:189–214
Larson ER,Van de Gevel SL,Grissino-Mayer HD(2009)Variability in fire regimes of high-elevation whitebark pine communities, western Montana,USA.E´coscience 16:282–298
Logan JA,Powell JA(2001)Ghost forests,global warming,and the mountain pine beetle(Coleoptera:Scolytidae).Am Entomol 47(3):160–173
Logan JA,Macfarlane WW,Wilcox L(2010)Whitebark pine vulnerability to climate-driven mountain pine beetle disturbance in the Greater Yellowstone Ecosystem.EcolAppl20(4):895–902
Lorenz JT,Sullivan KA(2009)Season differences in space use by Clark’s nutcracker in the Cascade Range.Condor 111:326–340
Lorenz TJ,Aubry C,Shoal R(2008)A review of the literature on seed fate in whitebark pine and the life history traits of Clark’snutcracker and pine squirrels.USDA Forest Service Pacific Northwest Research Station,General Technical Report,PNWGTR-742,p 62
Maher EL,Germino MJ,Hasselquist NJ(2005)Interactive effects of tree and herb cover on survivorship,physiology,and microclimate of conifer seedlings at the alpine tree-line ecotone.Can J For Res 35:567–574
Maloney PE,Vogler DR,Jensen CE,Delfino Mix A(2012)Ecology of whitebark pine populations in relation to white pine blister rustinfection in subalpine forests ofthe Lake Tahoe Basin,USA: implications for restoration.For Ecol Manag 280:166–175
Mattson DJ,Reinhart DP(1997)Excavation of red squirrel middens by grizzly bears in the whitebark pine zone.J Appl Ecol 34:926–940
Mattson DJ,Blanchard BM,Knight RR(1992)Yellowstone grizzly bear mortality,human habituation,and whitebark pine seed crops.J Wildl Manag 56:432–442
McCaughey W,Scott GL,Izlar KL(2009)Whitebark pine planting guide.West J Appl For 24(3):163–166
McKinney ST,Tomback DF(2007)The influence of white pine blister rust on seed dispersal in whitebark pine.Can J For Res 37:1044–1057
McKinney ST,Fielder CE,Tomback DF(2009)Invasive pathogen threatens bird-pine mutualism:implications for sustaining a high-elevation ecosystem.Ecol Appl 19(3):597–607
Mohatt KR,Cripps CL,Lavin M(2008)Ectomycorrhizal fungi of whitebark pine(a tree in peril)revealed by sporocarps and molecular analysis of mycorrhizae from treeline forests in the Greater Yellowstone Ecosystem.Botany 86:14–25
Morgan P,Bunting SC(1992)Using cone scars to estimate pastcone crops of whitebark pine.West J Appl For 7:71–73
Murray MP,Rasmussen M(2000)Status of whitebark pine in Crater Lake National Park.Unpublished Final Report,Cooperative Cost-Share Agreement No.H9320000035.Copy on file at US Departmentof Interior,Park Service,Crater Lake National Park, Resource Management Division 13 pp
Niinemets U¨(2006)The controversy over traits conferring shadetolerance in trees:ontogenic changes revisited.J Ecol 94:464–470
Perkins JL(2004)Pinus albicaulis seedling regeneration after fire. Ph.D.thesis,Division of Biological Sciences,University of Montana,Missoula,149 p
Pfister RP,Kovalchik BL,Arno SF,Presby R(1977)Forest habitat types of Montana.USDA Forest Service,Intermountain forest and range experimentstation,General Technical Report INT-34, Ogden,p 174
R Development Core Team(2008)R:a language and environmentfor statistical computing.R Foundation for Statistical Computing, Vienna
Richardson BA,Brunsfeld SJ,Klopfenstein NB(2002)DNA from a bird-dispersed seed and wind-disseminated pollen provides insights into postglacial colonization and population genetic structure of whitebark pine(Pinus albicaulis).Mol Ecol 11:215–227
Romme WH(1982)Fire and landscape diversity in subalpine forests of Yellowstone National Park.Ecol Monogr 52:199–221
Schoettle AW,Sniezko RA(2007)Proactive intervention to sustain high-elevation pine ecosystems threatened by white pine blister rust.J For Res 12:327–336
Schwandt JW(2006)Whitebark pine in peril:a case for restoration. USDA Forest Service Report R1-06-28
Scott GL,McCaughey WW(2006)Whitebark pine guidelines for planting prescriptions.In:Riley LE,Dumroese RK,Landis TD, (tech.coords.)National Proceedings:Forest and Conservation Nursery Associations—2005.Proceedings on RMRS-P-43 U.S. Department of Agriculture,Forest Service,Rocky Mountain Research Station,Fort Collins,p 84–90
Smith CM,Wilson B,Rasheed S,Walker RC,Carolin T,Shepherd B (2008)Whitebark pine and white pine blister rust in the Rocky Mountains of Canada and northern Montana.Can J For Res 38:982–995
Smith CM,Poll C,Gillies C,Praymak C,Miranda E,Hill J.2011. Limber pine seed and seedling planting experiment in Waterton Lakes National Park,Canada.In Keane RE,Tomback DF, Murray MP,Smith CM(eds)The future of high-elevation,fiveneedle white pines in western North America.Proceedings ofthe high five symposium;28–30 June 2010;Missoula.Proceedings on RMRS-P-63.U.S.Departmentof Agriculture,Forest Service, Rocky Mountain Research Station,Fort Collins,p 365–374
Tomback DF(1978)Foraging strategies of Clark’s nutcracker.Living Bird 16:123–161
Tomback DF(1982)Dispersal of whitebark pine seeds by Clark’s nutcracker:a mutualism hypothesis.J Anim Ecol 51:451–467
Tomback DF(2001)Clark’s nutcracker:agent of regeneration.In: Tomback DF,Arno SF,Keane RE(eds)Whitebark pine communities:ecology and restoration.Island Press,Washington, pp 89–107
Tomback DF,Achuff P(2010)Blister rust and western forest biodiversity:ecology,values and outlook for white pines.For Pathol 40:186–225
Tomback DF,Kendall KC(2001)Biodiversity losses:the downward spiral.In:Tomback DF,Arno SF,Keane RE(eds)Whitebark pine communities:ecology and restoration.Island Press,Washington,pp 243–262
Tomback DF,Sund SK,Hoffman LA(1993)Post-fire regeneration of Pinus albicaulis:height-age relationships,age structure,and microsite characteristics.Can J For Res 23:113–119
Tomback DF,Clary JK,Koehler J,Hoff RJ,Arno SF(1995)The effects of blister ruston post-fire regeneration of whitebark pine: the Sundance Burn of Northern Idaho(U.S.A.).Conserv Biol 9:654–664
Tomback DF,Anderies AJ,Carsey KS,Powell ML,Mellmann-Brown S(2001a)Delayed seed germination in whitebark pine and regeneration patterns following the Yellowstone fires. Ecology 82(9):2587–2600
Tomback DF,Arno SF,Keane RE(2001b)The compelling case for managementintervention.In:Tomback DF,Arno SF,Keane RE (eds)Whitebark pine communities:ecology and restoration. Island Press,Washington,pp 3–25
USDA Forest Service(2007)Common Stand Exam Users Guide and Appendices,Version 1.7.USDA Forest Service,Natural Resource Information System:Field Sampled Vegetation.Available online at<http://www.fs.fed.us/emc/nris/products/fsveg/ index.shtml>
Vander Wall SB,Balda RP(1977)Coadaptations of the Clark’s nutcracker and the pin˜on pine for efficient seed harvest and dispersal.Ecol Monogr 47:89–111
Walsh MR(2005)Fire history and stand dynamics of whitebark pine (Pinus albicaulis)communities in the Greater Yellowstone Ecosystem.MS Thesis,Colorado State University,Fort Collins, CO,48 p
Waring KM,Goodrich BA(2012)Artificial regeneration of fiveneedled pines of western North America:a survey of current practices and future needs.Tree Plant Notes 55:56–72
7 May 2014/Accepted:15 August 2014/Published online:28 April 2015
©Northeast Forestry University and Springer-Verlag Berlin Heidelberg 2015
Projectfounding:This work was supported by USDA Forest Service, Region 1 and 4,Whitebark Pine Restoration Fund,Agreement No. 07-CA-11010000-009 and the Colorado State University,Agricultural Experiment Station.
The online version is available athttp://www.springerlink.com
Corresponding editor:Chai Ruihai
✉Jennifer G.Klutsch klutsch@ualberta.ca;jklutsch@gmail.com
Betsy A.Goodrich betsy.goodrich@nau.edu
William R.Jacobi william.jacobi@colostate.edu
1Department of Bioagricultural Sciences and Pest Management,Colorado State University,Ft.Collins, CO 80523-1177,USA
2Present Address:Department of Renewable Resources, University of Alberta,442 Earth Science Building, Edmonton,AB T6G 2E3,Canada
3Present Address:School of Forestry,Northern Arizona University,PO Box 15018,Flagstaff,AZ 86011,USA
杂志排行

Journal of Forestry Research的其它文章
- Management of pests and diseases of tropical sericultural plants by using plant-derived products:a review
- Gamma generalized linear model to investigate the effects of climate variables on the area burned by forest fire in northeast China
- Diversity,abundance,and structure of tree communities in the Uluguru forests in the Morogoro region,Tanzania
- Brazilian savanna re-establishment in a monoculture forest: diversity and environmental relations of native regenerating understory in Pinus caribaea Morelet.stands
- Carbon storage and sequestration rate assessment and allometric model development in young teak plantations of tropical moist deciduous forest,India
- Use of infrared thermal imaging to diagnose health of Ammopiptanthus mongolicus in northwestern China