Diversity,abundance,and structure of tree communities in the Uluguru forests in the Morogoro region,Tanzania
2015-06-05DavidSylvesterKacholiAnthonyMichaelWhitbreadMartinWorbes
David Sylvester Kacholi•Anthony Michael Whitbread•Martin Worbes
Diversity,abundance,and structure of tree communities in the Uluguru forests in the Morogoro region,Tanzania
David Sylvester Kacholi1,2•Anthony Michael Whitbread1,3•Martin Worbes1
Uluguru forests are globally recognized as importantbiodiversity hotspots,butanthropogenic pressure threatens their value.This study examined species diversity,abundance,and structure of trees in the Uluguru forests.Alltrees ofdiameter atbreastheight(DBH)≥10 cm were inventoried in seven forests ranging from 3 to 995 ha in area.A totalof 900 stems,101 species and 34 families were inventoried.Fabaceae was the most speciose family. Ehretia amoena Klotzsch was the most abundant species with relative abundance of 9.22%.The forests differed significantly in species richness(26–93 species ha-1),tree density(85–390 stems ha-1),basal area(3–24 m2ha-1) and Shannon-Wienerdiversity(2.50–4.02).Forestarea was significantly and positively correlated with species richness (r=0.92)and species diversity(r=0.95).Tree density showed significant positive correlation with speciesrichness(r=0.80)and basal area(r=0.85).Milawilila and Nemele forests had highest floristic similarity(0.55) followed by Kimboza and Kilengwe(0.54)while the rest had similarity coefficients of less than 0.50.Despite legislative protection,many forests remain at risk and therefore the possibility to conserve highly valuable tree species via enhanced protection or cultivation mustbe considered.
Eastern arc·Biodiversity·Disturbance· Hotspots·Similarity
Introduction
Tree species distribution differs greatly from one place to the other in mosttropicalforests,mainly due to variations in biogeography,habitat and disturbances(Whitmore 1989).The variety of tree species richness has been reported in the neo-tropical forests,tropical forests,and Southeast Asia forests(Gentry 1988;Whitmore 1989; Valencia etal.1994).In African forests,a maximum of60 species ha-1was reported,with a number of other studies (Hill and Curran 2001;Mwavu 2007;Kacholi 2013)reporting much higher species richness than this figure for trees of DBH≥10 cm.Though tropicalforests are known to be speciose,they are vulnerable to deforestation and degradation(Madoffe et al.2006),which ultimately leads to fragmentation and loss of habitats.In order to guide nature conservation efforts worldwide,Myers et al.(2000) focused on the concept of biodiversity hotspots,which considers regions with exceptional concentrations of endemic species and which experience high rates of habitat loss due to natural and anthropogenic degradation.The authors proposed thatprotection and conservation activities should be focused on these hotspots.The Eastern Arc andother coastal forests(Uluguru forests inclusive)of Tanzania are one of the identified 25 global hotspots and are estimated to host 4000 plant species of which 38%are endemic.
Uluguru forests are facing the threat of losing species due to increased anthropogenic activities and fragmentation(Newmark 1998).Increased forest fragmentation has been described as the greatest threat to much of tropical forestbiodiversity(Hill and Curran 2001).A rapid human population increase in proximity to many tropicalforests is putting more pressure on these ecosystems via the demand for timber for building purposes,firewood/charcoaling,the provision of food and medicine or increased demand for farmland.Increased wildfire risk from accidentaland nonaccidentallightings is a furtherthreat(Burgess etal.2002). Such threats,which should not be underrated,are also occurring in other biodiversity hotspots within the country (Madoffe et al.2006).More attention is needed for research and conservation in these ecosystems.Studies of floristic composition and structure in forests have become an essentialinstrumentin assessing the sustainability of the forests and the role they play in the conservation of species and management of forest ecosystems(Malimbwi et al. 2005).The presentstudy aimed to:(1)provide information on the status offloristic diversity,abundance,and structure of trees of DBH≥10 cm in the selected Uluguru forests; (2)compare the present findings with other forest inventories done in Tanzania and elsewhere,and(3)determine the relationship between forest area and numbers of tree species,tree density,basal area,and several common measures of diversity.We aimed to improve knowledge of the status of tropical rainforests and contribute to biodiversity managementand conservation.
Materials and methods
Description of the study area
Uluguru Mountains are located about 200 km west of Dar es Salaam City,and south of Morogoro town.The range is one components of the Eastern Arc Mountains,stretching from the Taita Hills in southern Kenya to Udzungwa Mountain in south-central Tanzania.The range covers an area of 1500 km2and elevation ranges from 150 m on the southeastern margin to a peak of 2630 m above mean sea level.The climate is oceanic due to proximity to the Indian Ocean with a bimodal rainfall regime,the long rainy season lasts from March to May,peaking in April and the short rainy season lasts from October to December.Mean annual rainfall and temperature are 740 mm and 25.1°C, respectively.Agriculture is the main socioeconomic activity for most people living in villages surrounding the studied forests.Seven forests were selected to represent lowland forest(i.e.<800 m above mean sea level)with minimum anthropological disturbance,and to represent a range of forest areas(Fig.1).
Data collection
Alltrees with a diameter atbreastheight(DBH)≥10 cm measured at1.3 m above the ground were sampled from a total of 114 plots of 20×20 m(0.04 ha)each.Eighteen plots were sampled in each forest at Kimboza,Kisego, Kilengwe,Milawilila and Nemele while 12 plots of the same size were sampled at Ngambaula and Gunauye.The plots were placed in the forests from the edge towards the interior.Trees were counted,identified and DBH were measured.Trees with multiple stems at1.3 m heightwere treated as a single individual whereby the diameters of all stems were taken and averaged.If a tree had buttress and abnormality at 1.3 m height,the diameter was measured just above the buttress where the stem assumed near cylindrical shape.These data were collected from June 2010 to February 2011.
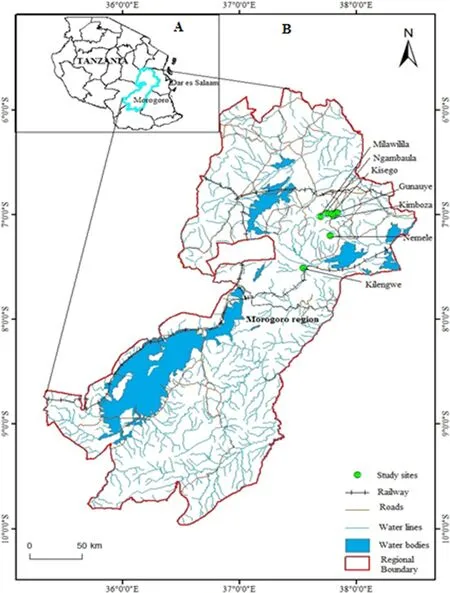
Fig.1 The figure shows two maps,the map of Tanzania(map A,to the top left corner)showing the setting of Morogoro region in the country and the map of Morogoro region(map B)that shows the location of the seven studied forests within the region
Data analysis
Species diversity was calculated using the Shannon-Wiener diversity index while the structure of the forests was described by stem density(stems ha-1),basalarea(m2ha-1) and size class distributions(SCDs).A totalof 7 size class distributions arranged in 10 cm intervals was formed based on recorded trees diameters in all forests.Differences of the structural characteristics(i.e.stand density and basal area)between forests were tested using one-way analysis of variance(ANOVA)followed by the posthoc Tukey’s HSD multiple comparison test.The number of recorded species in each forest and the first order jackknife richness estimator were used as measure ofspecies richness(Magurran 2004).Species richness was calculated and species accumulation curves were constructed using Species Richness and Diversity IV(SDR IV)Software(Seaby and Henderson 2006a).Sørensen coefficients of similarities were calculated between the studied forest pairs and Pearson correlation coefficients were calculated to determine the relationship between floristic similarity,forestarea,species diversity,tree density and basalarea.The coefficients were calculated using the Community Analysis Package version 4 software(CAP IV)(Seaby and Henderson 2006b).
Results
Floristic description
A totalof 900 trees(1335 stems ha-1)with DBH≥10 cm representing 101 species,73 genera and 34 families were recorded in the seven forests(Table 3 in Appendix).The most speciose family was Fabaceae(with 33 species), followed by Moraceae(6 species)and Sterculiaceae(5 species).Of the 34 recorded families,38%were represented by one species only while out of the 101 recorded species,17%were represented by one individual only. Ehretia amoena was the most abundant species representing 9.2%of the total stems,followed by Sorindeia madagascariensis(6.1%)and Khaya anthotheca(4.7%). E.amoena and Diospyros squarrosa were the most frequent species occurring in six of the surveyed forests. About 41.6%of the total species occurred in one forest while 15.0%of species occurred in one plot out of 114 surveyed plots.Observed species richness(Table 1)varied widely from 17 species(at Ngambaula)to 67 species(at Kilengwe).Of all recorded species in all forests,13 are considered threatened on the 2014 IUCN plant redlist,of which one is Endangered(Cynometra uluguruensis),five are Vulnerable(K.anthotheca,Ophrypetalum odoratum, Millettia sacleuxii,Allanblackia uluguruensis,Allanblackia stuhlmannii),five are Near Threatened/Lower Risk (Milicia excelsa,Pterocarpus angolensis,Pandanus rabaiensis,D.melanoxylon,Pouteria altissima),and two are Least Concern(Cussonia zimmermannii and Holarrhena pubescens).Among the threatened species,two were endemic(A.uluguruensis,C.uluguruensis)and three were nearendemic(Scorodophloeus fischeri,A.stuhlmannii,and O.odoratum).
In Kilengwe,a total of 199 trees(276 stems ha-1)belonging to 67 species(93 species ha-1),26 families,and 54 genera were recorded.Brachystegia speciformis was most abundantwith 8 individuals while 28.4%of the species in the studied forest area were rare,being represented by only one individual.In Kimboza,a total of 281 trees (390 stems ha-1)belonging to 52 species (72 species ha-1),22 families,and 39 genera were recorded.K.anthotheca wasmostabundant(37 individuals) followed by S.madagascariensis(27)and E.amoena(25). Of the Kimboza species,23.1%were rare,being represented by only one individual.In Milawilila,a totalof 124 trees(172 stems ha-1)of 20 species(28 species ha-1),15 families,and 18 genera were recorded.Xylopia parviflora was dominant with 26 individuals,followed by E.amoena with 21 individuals.Only one species was represented by a single individualin Milawilila forest.
In Kisego,101 trees(140 stems ha-1)representing 21 species(29 species ha-1),12 families,and 19 genera were recorded.The most abundant species were E.amoena(23 individuals)and Albizia versicolor(14 individuals),while 33.3%of the species were represented by only one individual.In Nemele,70 trees(97 stems ha-1)of 19 species (26 species ha-1),13 families,and 16 genera were recorded.The most abundant species were E.amoena(8 individual)followed by Voacanga Africana(7 individuals), and 31.6%of the species were represented by one individual.In Ngambaula,a totalof41 trees(86 stems ha-1)of 17 species(35 species ha-1),9 families,and 15 genera were recorded.In Gunauye,a totalof 84 trees(175 stems ha-1)of 22 species(46 species ha-1),14 families,and 19 genera were recorded.Brachystegia boehmii and D. squarrosa were the most dominant species at Ngambaula while S.madagascariensis was mostabundantatGunauye. Of the recorded species,41.7 and 22.7%were represented by one individual at Ngambaula and Gunauye, respectively.
Species diversity and species accumulation curves
Kilengwe and Kimboza were the most diverse forests, having highest diversity indices of 4.02 and 3.40,respectively(Table 1).The least diverse forest was Kisego with 2.50 diversity value.When all data are pooled,an overall Shannon-Wienerdiversity index of4.03 was obtained.The species accumulation curves for Kimboza and Kilengwe(Fig.2)showed an increasing trend as the number of plots increased while the curves for Milawilila,Kisego,Ngambaula,Nemele and Gunauye rapidly approached an asymptote.The first order jackknife species richness estimatorcalculated higherspecies richness than was recorded in the field(Table 1).
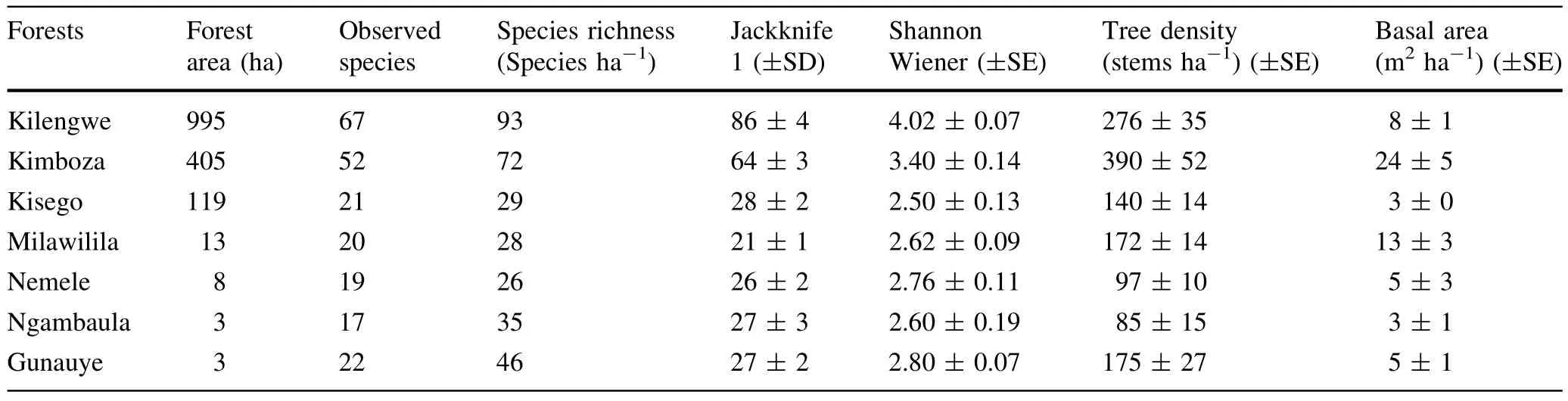
Table 1 Forest size,tree species richness,diversity,density and basal areas in the studied forests

Fig.2 Species accumulation curves of tree species based on the cumulative plot samples in each of the seven studied forest areas
Correlation between forest area,structure,species richness and diversity
Forest area was significantly positively correlated with species richness and species diversity(Table 2).Tree density was positively and significantly correlated with species richness and basal area density.Although tree density and basal area were positively correlated with forest area,the association was not statistically significant.
Species composition similarity
Species composition similarity between the studied forests revealed higher similarity coefficientbetween Nemele and Milawilila(0.55),followed by Kimboza and Kilengwe (0.54).The remaining forest pairs had similarity coefficients of less than 0.50.The lowest similarity coefficient was 0.20 between Nemele and Kisego(Fig.3).
Structuralcomposition of the forests
Tree density varied significantly between forests(F(6,107)= 14.37,p<0.001),ranging from 85 to 390 stems ha-1(Table 1).Kimboza supported considerably greater density than the otherforests with the exception of Kilengwe,which did not differ considerably.Tree density at Kilengwe was considerably higherthan atNemele,Ngambaula and Kisego butdid notdiffersignificantly from density atMilawilila and Gunauye.Other forest pairs did not differ appreciably in terms of tree density.Basal area density differed greatly between forests(F(6,107)=9.92,p<0.0001),ranging from 3 to 24 m2ha-1(Table 1).Kimboza had significantly higher basal area followed by Milawilila and Kilengwe while Kisego had the lowestbasalarea density.
The size class distribution of trees(Fig.4)in the forests exhibited the‘‘negative exponential’’or‘‘inverse J-shape’’. About69.1 and 18.2%ofrecorded trees in allforests were represented in 10–19.9 and 20–29.9 cm DBH size classes, respectively.Kisego and Gunauye had no individualin the size classes’≥50 cm DBH.Though Nemele and Ngambaula possessed individuals in size class>70 cm DBH,the forests had no individuals in size classes between 40 and<70 cm DBH.The maximum DBH value in the present study was 126 cm recorded for K.anthotheca at Kimboza forest,followed by Brachystegia bussei(Nemele)and Synsepalum cerasiferum(Milawilila)at 122 and 106 cm, respectively.
Discussion
Floristic composition,species richness and diversity
The dominant family in our research area is Fabaceae with 33.7%of the individualtrees.This resultconcurs with thefindings of Burgess and Muir(1994)and Burgess and Clarke(2000)who reported the same family to dominate the coastal forests of Tanzania by 25%–50%of trees. Mrema(2006)and Rwamugira(2008)reported the family Fabaceae to be dominant in Dindili and Ruvu forests in Tanzania,respectively.Moreover,the finding confirms the verdict of different authors(e.g.Gentry 1988;Valencia et al.1994;Mwavu 2007;Addo-Fordjour etal.2009)that the Fabaceae family is the most speciose tree family in many tropical lowland forests.

Table 2 Correlations between forest size,species richness,diversity,density and basal area
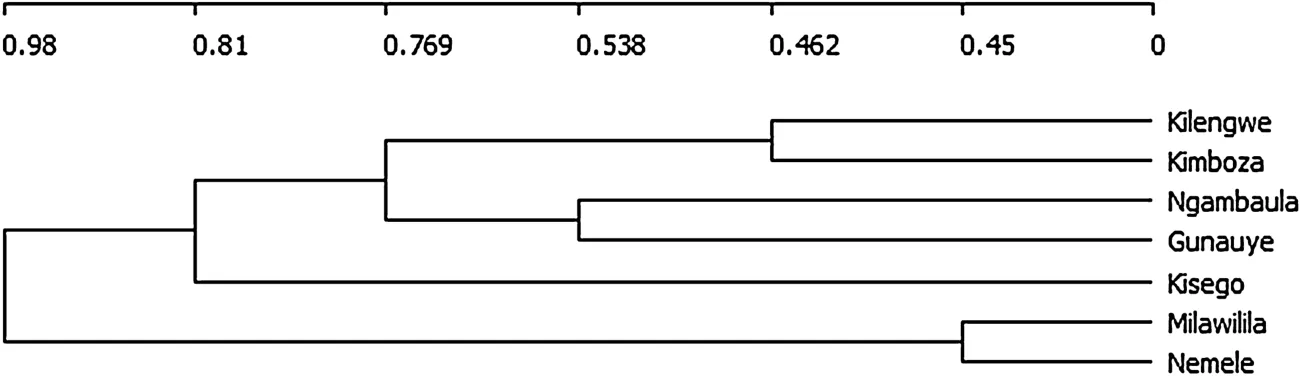
Fig.3 The dendrogram constructed using Ward group linkage and Sørensen coefficients of similarity shows dissimilarity coefficients among the seven studied forest pairs in Uluguru.The dissimilarity coefficients increase as the scale increases while similarity increases in an opposite direction of the dendogram scale
The range of species richness(26–93 species ha-1)in this study overlaps thatreported by Malimbwietal.(2005) of 8–66 species ha-1in different lowland forests of Mvomero district in Morogoro region.However,the range is within the range reported by Mwavu(2007)of 24–112 species ha-1in Budongo forests in Uganda.In contrast,the recorded species richness appears to be lower when compared to Amazonian forests where species richness of 275–283 species ha-1was reported for upper Amazonia (Gentry 1988).The comparisons involving differentstudies are complicated due to fact that different plot sizes,sampling protocols,total used sampled area and sometimes subjectivity used to arrive at a range of values in other studies is unclear.The occurrence of D.squarrosa and E. amoena in the six forests and S.madagascariensis,S.fischeri,Oxyanthus goetzei,and A.versicolor in five forests indicate that they thrive across a wide range of habitats.
According to Kent and Coker(1992),a forest community is said to be rich if ithas a Shannon-Wiener diversity index value of≥3.5.With the exception of Kilengwe forest,the resthad Shannon-Wienerdiversity values below 3.5 making them relatively poor in diversity.The overall diversity index of 4.03 for all seven forests signifies that Uluguru forests as a whole supporthigh diversity.The high diversity in Kilengwe and Kimboza could be due to fact thatthese forests are relatively undisturbed as compared to other studied forests where illegal logging,encroachment, hunting,removal of tree bark for medicinal use,fire,and collection of forest products were observed to be the main activities of the localpopulation.A smallscale gold mining was seen near Kimboza forest and this could account for the low recorded tree diversity.
The presence ofthreatened species in the studied forests could be because this area is within the Eastern Arc of coastal forests of Tanzania,which are known to be biodiversity hotspots and centers ofendemism forboth flora and fauna(Myers et al.2000).Temu and Andrew(2008)also found that the Uluguru forests contain several endemic plant species while the study area is among the recognized importantecoregions and an endemic bird area in Tanzania (Mittermeier et al.1998;Stattersfield et al.1998;Burgess etal.2007).The floristic composition,overalldiversity and threatened species listed in the IUCN categories show that the Uluguru forests are qualitatively diverse.Additionally, among the threatened species,P.angolensis,D.melanoxylon,K.anthotheca,and M.excelsa have also been reported by several authors(e.g.Ahrends 2005;Modest et al.2010)to be severely exploited for timber in the coastal forests of Tanzania and some logged below the minimum harvestable diameter.This could be the reason why some of these species occurred at very low frequencies in the sampled forests.

Fig.4 Size class distributions(SDC’s)of the trees in the studied Uluguru forests
Species accumulation curves
The species accumulation curves for Kilengwe and Kimboza forests(Fig.2)illustrated an escalating trend as the number of plots increased.This observation concurs with the results shown in Table 1 where the species richness estimator predicted more species in the forests than were recorded.The species accumulation curves for Milawilila,Nemele,Ngambaula and Gunauye approached an asymptote signifying thatmostof the species in these forests were recorded(Magurran 2004).This is also supported by the species richness estimator,which provides estimates that did notdiffer markedly from the recorded species richness. On the contrary,the curves for Kilengwe and Kimboza did notshow asymptotic behavior due to the presence ofmany rare species and species with narrow habitatranges(Gotelli and Colwell2011).The high species richness in Kilengwe and Kimboza could also be attributed to their large areas and high environmental heterogeneity(Tuomisto and Ruokolainen 2005).The increasing trend in the number of species with increasing forest size implies that a reduction in forest size will lead to species loss as supported by the correlation coefficients between forest size and species richness in Table 2.
Association between forest size,structure,species richness,and diversity
The significantassociations between forestarea and species richness/diversity(Table 2),support the hypothesis that large forestscontain greaternumbersoftree species.Also,it suggeststhatincreased forestfragmentation which normally resultsin area reduction would cause the lossoftree species. The smallerthe population,the more vulnerable itis to extinction when fluctuations in microclimate,resources and otherfactors occur(Hilland Curran 2001).Thus,large forests are needed by many species in order to maintain viable populations,though it is important also to recognize the complementary value of smallforestfragments.Forestsize and stand were positively correlated,suggesting that the density of trees is greater in larger forest fragments than smaller ones.Thus,in case of forest fragmentation,stand density isexpected to decline too.The significantcorrelation between basalarea density and tree density signifies thata decline in stand density would lead to decreased basalarea density.Species richness,diversity,stand density and basal area density are reported to decline in response to increased intensity of anthropogenic disturbance in tropical forests (Top etal.2009;Kacholi2013).
Similarity among the studied forests
The species composition similarity coefficients among the studied forestsranged from 0.20 to 0.55(Fig.3).The highest similarity between Milawilila and Nemele,and thatof Kimboza and Kilengwe could be attributed to similar environmentalconditionsamong the pairsascompared to Kisego and Nemele which had leastsimilarity value.With exception of the two forestpairs thatshowed high floristic similarity,the remaining pairs had similarity coefficients below 0.50,signifying that each forest has its own unique species composition.Thus,allthe forestsare importantin termsofthe floristic diversity and sensitive from a conservation pointof view.
Structural composition of the forests
The observed basalarea densities ofthe studied forests are within the range(1.7–32 m2ha-1)reported by Malimbwi etal.(2005)in theirinventories of the forests of Morogoro region in Tanzania.The considerably higher basal area density in Kimboza and Milawilila was contributed by high stem density in the higher DBH classes as compared to other forests.Kimboza revealed a 54%increase in basal area density from the value(15.8 m2ha-1)observed by Malimbwietal.(2005),which signifies thatthe foresthas notbeen greatly affected by human disturbance during the intervalfrom 2005 to 2010.In contrast,the observed basal area density atKilengwe was lowerby 29%than the value (11.2 m2ha-1)observed by Malimbwi et al.(2005),implying that the forest was impacted by anthropogenic disturbances.The significantly lower basal areas at Kilengwe, Kisego,Nemele,Ngambaula and Gunauye suggest that these forests are overexploited and in reality no big trees were recorded in these forests(Fig.4).Rwamugira(2008) reported average basal area of 4.7 m2ha-1(for trees with DBH≥10 cm)at a disturbed stand in the Ruvu forest in the Morogoro region,while Malimbwi et al.(2005) recorded very low basal area of 1.7 m2ha-1at Mindu forestin the same region,and concluded thatthe forestwas disturbed.Illegal logging is the main cause for low basal areas in many forests in Morogoro(Malimbwi et al.2005) and itis reported to be done by wellcoordinated syndicates involving traders,irresponsible local government leaders and unfaithful villagers(Kacholi 2013).
The size class distributions of Kilengwe,Kisego,Nemele, Ngambaula and Gunauye indicate that the tree species are recruiting and there are signs ofrecovery from the effects of previous and on-going disturbances.Illegallogging was also observed in these forests,indicating that the forests are still under anthropogenic pressure.The anthropogenic pressure could have resulted to the absence ofindividualsin the higher sizeclassesin theforests(Fig.4)becausebig treesare illegally logged by locals for various purposes.The significantdifferences in the observed structuralfeatures between the studied forests are mainly due to anthropogenic exploitation,which targetstreesofhigh size classesfortimberand building poles. Other factors such as soils,habitat preferences/adaptation ability between species,and presence offavorable conditions for regeneration have been reported to affect forest structure (Richards 1952;Zegeye et al.2006).Our results show that forests ofsmallerarea had lowerstand density and basalarea (Table 2).The reason could be thatthe smallfragments are highly vulnerable to human disturbances because they areeasily accessibleforlogging and clearanceactivitiesthataffect the foreststructure as reported by Echeverria etal.(2007).In Madagascar,the spatial pattern analysis of forest structure revealed thatlevels of basalarea were associated with accessibility to the fragments(Ingram etal.2005).
Conclusion and recommendations
Understanding foresttreediversity,abundance,and diversity are very importantin managementof the ecosystem for environmentaland conservation value.This study has revealed that family Fabaceae was the most speciose family in the forests while D.squarrosa and E.amoena were the most common species occurring in six forests.Species richness, diversity and tree density were positively correlated with forest size.Though the smaller forests had lower species richness,they mustbegiven priority in conservation to avoid lossofspeciesespecially endemic and nearendemic species. The structuralparameters differed significantly between the studied stands.The floristic similarity revealed low similarity coefficients among many forest pairs.Despite legislative protection,many forestfragmentsin Uluguru remain at risk and therefore the possibility to conserve highly
valuable tree species via enhanced protection or cultivation must be considered.The study recommends the following; (1)more conservation and managementeffortsshould be put to rare and threatened species,(2)research is needed to investigate major causes,types and level of anthropogenic disturbances to forests,(3)study of soils in Uluguru forest fragments is needed to understand their influence on tree composition and distribution,(4)research on the effects of climate change and land use/coverchange is needed to understand changes over time within and around the forests, and(5)protection of forests should be emphasized so that future generation can enjoy nature and meettheir needs.
AcknowledgmentsThe authors would like to thank the Dar es Salaam University College of Education(DUCE)and Deutscher Akademischer Austausch Dienst(DAAD)for funding the projectthat led to this article.The authors are also indebted to Prof.Dr.Teja Tscharntke,Prof.Dr.Ralph Mitlo¨hner and other anonymous reviewers for their constructive criticisms and suggestions to improve this study.Ahsanteni Sana.
Appendix
See Table 3.
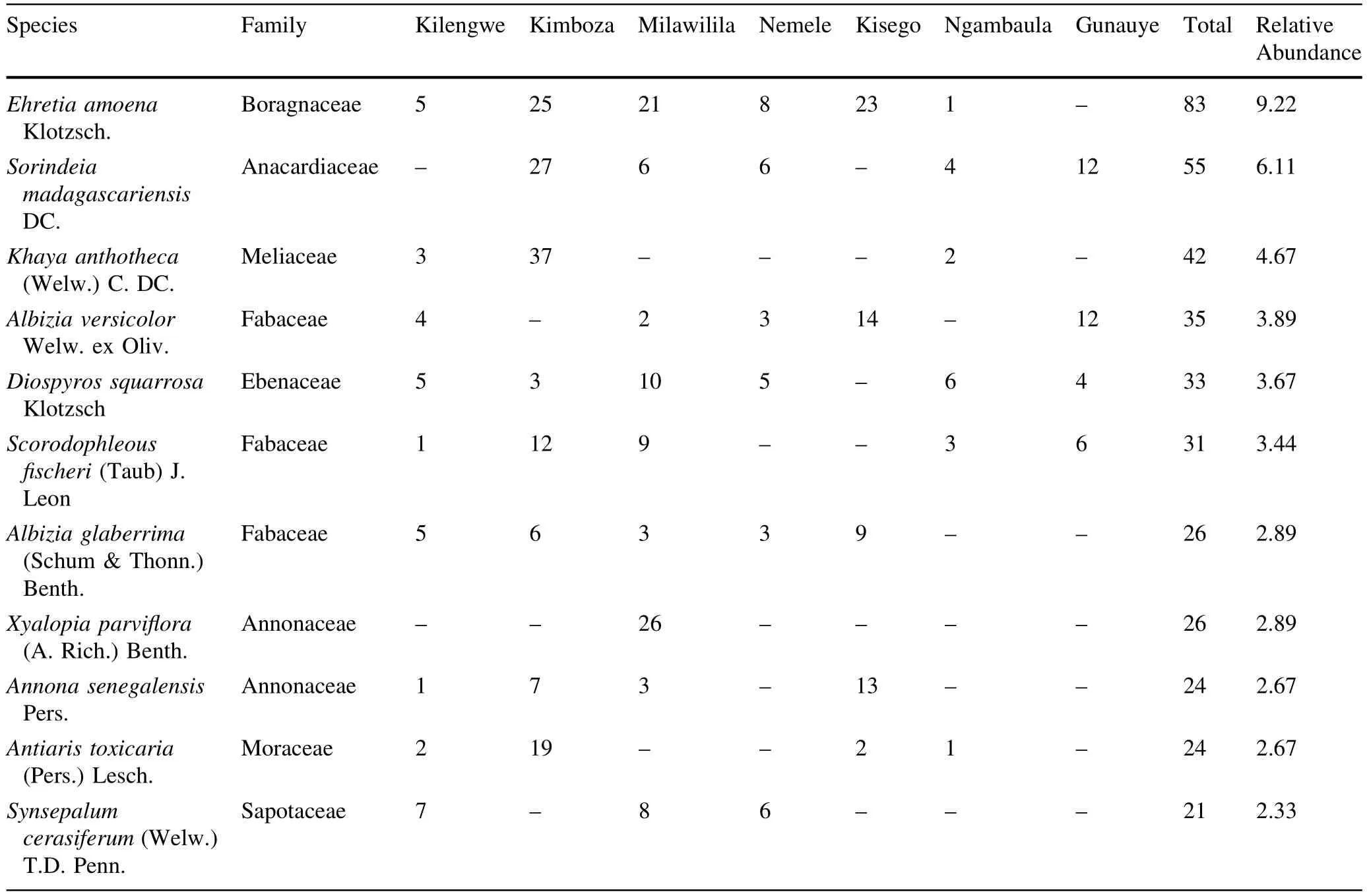
Table 3 Tree species abundance in the surveyed forests
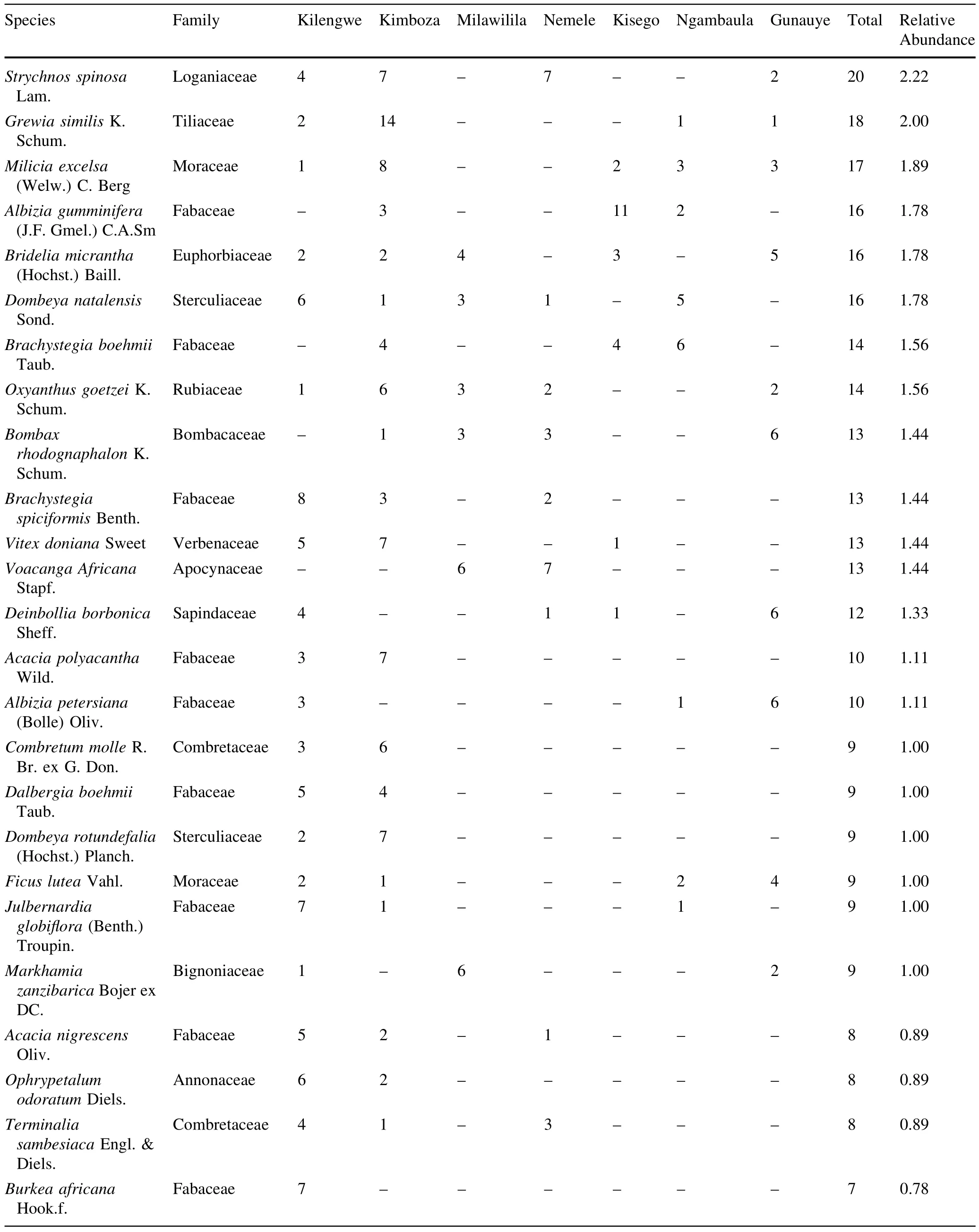
Table 3 continued
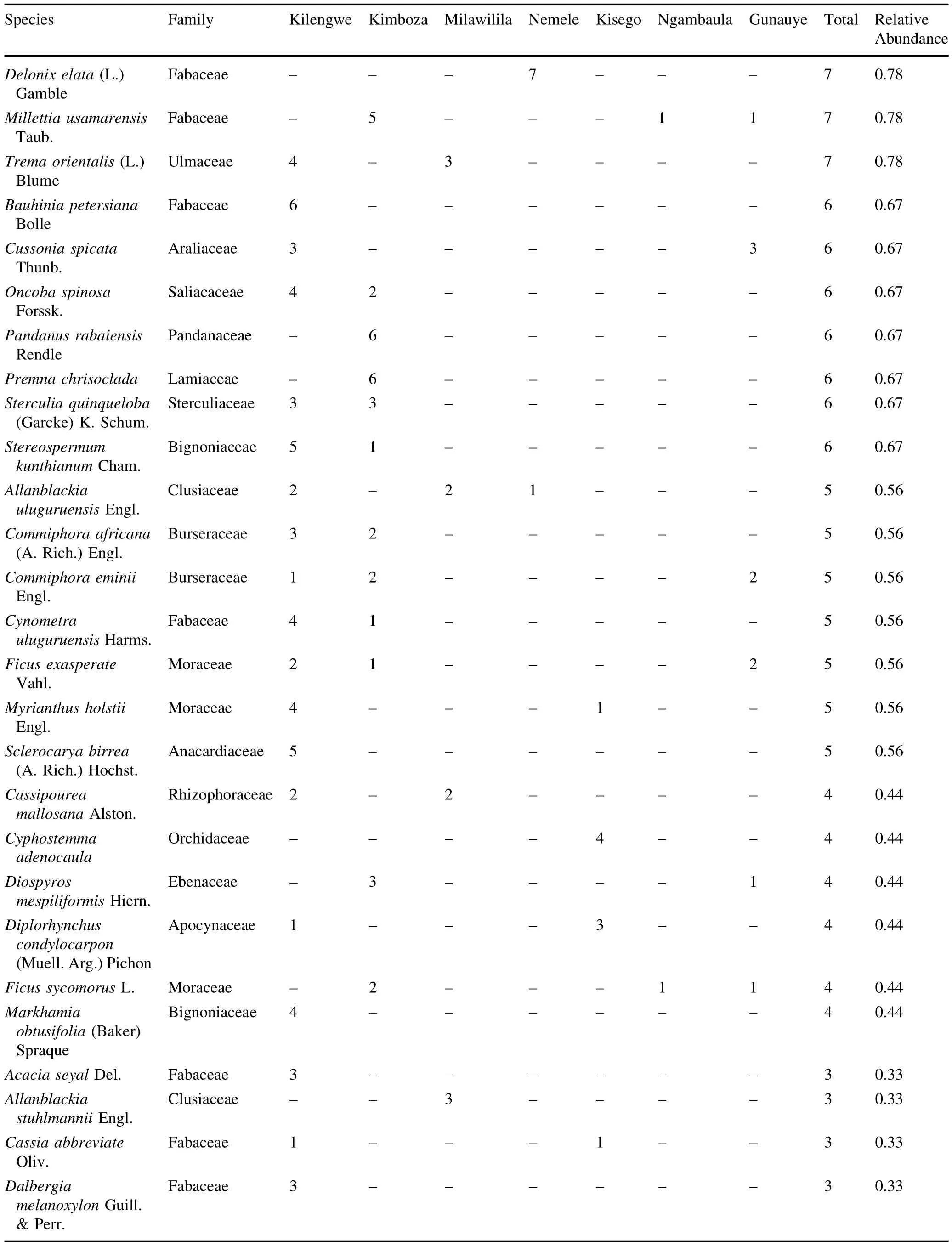
Table 3 continued

Table 3 continued

Table 3 continued
Addo-Fordjour P,Obeng S,Anning K,Add MG(2009)Floristic composition,structure and natural regeneration in a moist-semi deciduous forestfollowing anthropogenic disturbances and plant invasion.Int J Biodivers Conserv 1:021–037
Ahrends A(2005)Patterns of degradation in lowland coastal forests in coastal region Tanzania M.Sc.Dissertation,Germany:Ernst-Moritz-Arndt University of Greifswald,p 150
Burgess ND,Clarke GP(2000)The coastal forests of Eastern Africa. IUCN,Cambridge,p 443
Burgess ND,Muir C(1994)Coastal forests of Eastern Africa: Biodiversity and Conservation.In:Proceedings of a workshop held at the University of Dar es Salaam,August 9–11,1993. London(UK):Society for Environmental Exploration/Royal society for the protection of birds,United Kingdom,p 112
Burgess N,Dorggart N,Lovett JC(2002)The Uluguru Mountains of eastern Tanzania:the effects of forestloss on biodiversity.Oryx 36:140–152
Burgess ND,Butynski TM,Cordeiro NJ,Doggart NH,Fjeldsa J, Howell KM,Kilahama FB,Loader SP,Lovett JC,Mbilinyi B, Menegon M,Moyer DC,Nashanda E,Perkin A,Rovero F, Stanley WT,Stuart SN(2007)The biological importance of the Eastern African Mountains of Tanzania and Kenya.Biol Conserv 134:209–231
Echeverria C,Newton AC,Lara A,Benayas JMR,Coomes DA (2007)Impacts of forest fragmentation on species composition and foreststructure in the temperate landscape ofsouthern Chile. Glob Ecol Biogeogr 16:426–439
Gentry A(1988)Changes in plant community diversity and floristic composition on environmental and geographical gradients.Ann Mo Botan Gard 75:1–34
Gotelli NJ,Colwell RK(2011)Estimating species richness.In: Magurran AE,McGill BJ(eds)Biological diversity:frontiers and measurementand assessment.Oxford University Press,New York,pp 39–54
Hill JL,Curran PJ(2001)Species composition in fragmented forests: conservation implications of changing forest area.Appl Geogr 21:157–174
Ingram JC,Dawson TP,Whittaker RJ(2005)Mapping tropicalforest structure in southern Madagascar using remote sensing and artificial neural networks.Remote Sens Environ 94:491–507
Kacholi DS(2013)Effects of habitatfragmentation on biodiversity of Uluguru Mountain forests in Morogoro region,Tanzania.Ph.DDissertation,Georg-August University,Goettingen,Cuvillier Verlag,Germany,p 150
Kent M,Coker P(1992)Vegetation description and analysis.CRC Belhaven Press,London,p 363
Madoffe S,Hertel GD,Paul R,Connell BO,Killenga R(2006) Monitoring health of selected Eastern Arc Forests in Tanzania. Afr J Ecol44:171–177
Magurran AN(2004)Measuring biological diversity.A Blackwell Publishing Company,Melden
Malimbwi RE,Shemweta DTK,Zahabu E,Kingazi SP,Katani JZ, Silayo DA(2005)Forest inventory for Mvomero district, Morogoro Tanzania:Forconsult Report
Mittermeier RA,Myers N,Thompsen JB,da Fonesca GAB,OlivieriS (1998)Global biodiversity hotspots and major tropical wilderness areas.Conserv Biol 12:516–520
Modest RB,Maganga SLS,Hassan SN,Mariki SB,Muganda M (2010)Population structure and extraction of three commercial trees in Nguru ya Ndege forest reserve,Morogoro-Tanzania. Ethiop J Environ Studies Manag 3:41–48
Mwavu EN(2007)Human impact,plant communities,diversity and regeneration in Budongo forestreserve,North-Western Uganda. Ph.D Thesis,University of Witwatersrand,South Africa,p 197
Myers N,Mittermeier RA,Mittermeier CG,da Fonseca GAB,Kent J (2000)Biodiversity hotspots for conservation priorities.Nature 403:853–858
Newmark WD(1998)Forest area,fragmentation,and loss in the Eastern Arc Mountains:implications for the conservation of biological diversity.J East Afr Natural Hist 87:29–36
Richards PW(1952)The tropical rain forests.Cambridge University Press,Cambridge,p 450
Rwamugira SP(2008)Impact of mining on forest ecosystems and adjacentcommunities of Eastern Arc Mountains.A case study of Ruvu catchment forest reserve.M.Sc.Dissertation,Sokoine University of Agriculture,Morogoro,Tanzania,p 134
Seaby RM,Henderson PA(2006a)Species diversity and richness 4. Pisces Conservation Ltd,Lymington,p 123
Seaby RM,Henderson PA(2006b)Community analysis package 4. Pisces Conservation Ltd,Lymington,p 123
Stattersfield AJ,Crosby MJ,Long AJ,Wege DC(1998)Endemic bird areas of the world:priorities for biodiversity conservation. BirdLife conservation series No 7,BirdLife International, Cambridge,p 846
Temu RPC,Andrew SM(2008)Endemism of plants in the Uluguru Mountains,Morogoro,Tanzania.For Ecol Manag 255:2858–2869
Top N,Mizoue N,Ito S,Kai S,Nakao T,Ty S(2009)Effects of population density on tree structure and species richness and diversity of trees in Kampong Thom Province,Cambodia. Biodivers Conserv 18:717–738
Tuomisto H,Ruokolainen K(2005)Environmentalheterogeneity and the diversity of pteridophytes and Melastomataceae in Western Amazonia.Biol Skr 55:37–56
Valencia R,Balslev H,Pay YM(1994)High tree alpha-diversity in Amazonian Ecuador.Biodivers Conserv 3:21–28
Mrema JP(2006)Conservation of Brachylaena huillensis O.Hoffm (Asteraceae)in Dindili Forest Reserve Morogoro.MSc Thesis, Addis Ababa University,Ethiopia,p 86
Whitmore TC(1989)Southeast Asia tropicalforests.In:H Lieth and MJH.Werger(eds),Ecosystems of the World.Tropical rainforests ecosystems,Elsevier,Amsterdam,195–218
Zegeye H,Teketay D,Kelbessa E(2006)Diversity,regeneration status and socio-economic importance of vegetation in the islands of Lake Ziway,South-Central Ethiopia.Flora 201:483–498
15 May 2014/Accepted:17 July 2014/Published online:30 April 2015
©Northeast Forestry University and Springer-Verlag Berlin Heidelberg 2015
Project funding:This work was supported by the Dar es Salaam University College of Education(DUCE)and Deutscher Akademischer Austausch Dienst(DAAD).
The online version is available at http://www.springerlink.com.
Corresponding editor:Zhu Hong.
✉David Sylvester Kacholi kacholi78@yahoo.com
1Crop Production Systems in the Tropics,Georg-August University Goettingen,Grisebachstr.6,37077 Goettingen, Germany
2Departmentof BiologicalSciences,Dares Salaam University College of Education(DUCE), P.O.Box 2329,Dar es Salaam,Tanzania
3International Crops Research Institute for the Semi-Arid Tropics(ICRISAT),Patancheru 502324,Andhra Pradesh, India
杂志排行

Journal of Forestry Research的其它文章
- Management of pests and diseases of tropical sericultural plants by using plant-derived products:a review
- Gamma generalized linear model to investigate the effects of climate variables on the area burned by forest fire in northeast China
- Brazilian savanna re-establishment in a monoculture forest: diversity and environmental relations of native regenerating understory in Pinus caribaea Morelet.stands
- Carbon storage and sequestration rate assessment and allometric model development in young teak plantations of tropical moist deciduous forest,India
- Use of infrared thermal imaging to diagnose health of Ammopiptanthus mongolicus in northwestern China
- Analysis of three types of triterpenoids in tetraploid white birches (Betula platyphylla Suk.)and selection of plus trees