利用F2群体定位水稻谷粒长宽比QTL
2015-05-28马孟莉刘艳红苏一兰卢丙越
马孟莉,刘艳红,江 玲,雷 恩,苏一兰,卢丙越*
(1.红河学院 生命科学与技术学院/云南省高校农作物优质高效栽培与安全控制重点实验室,云南 蒙自 661100;2.南京农业大学 作物遗传与种质创新国家重点实验室,江苏 南京 210095)
水稻是世界上最主要的粮食作物之一,全球有超过50%的人口以稻米为主食,特别是亚洲地区[1]。水稻是我国第一大粮食作物,长期以来高产一直作为我国的水稻育种的主要目标,而忽视了稻米品质的改良,在人民生活水平日益提高的今天,劣质稻谷严重限制了其在国内和国际市场的竞争力[2]。深入了解稻米品质的遗传规律,鉴定、定位相关基因/QTL及进一步精细定位和克隆,对加速利用分子标记辅助选择改良水稻品质具有重要意义。
粒型(粒长、粒宽、长宽比)是稻米外观品质的主要衡量指标。近年来,随着DNA分子标记技术的不断发展,国内外研究者利用不同的群体定位到多个控制稻谷粒型的QTL,像GS3、GW5、GW2、Ghd7、GW8、GS5、qGL3 等控制粒长、粒宽的 QTL 先后被克隆(http://www.ricedata.cn/index.htm),为揭示粒型的遗传机理奠定了良好的基础,而至今未见水稻长宽比QTL克隆的相关报道。已有研究表明控制水稻谷粒长宽比的QTL广泛分布于12条染色体上,且不同的定位群体检测到的长宽比QTL存在很大差异[3],挖掘不同材料中控制谷粒长宽比的基因对深入研究粒型的遗传机制具有重要的意义。本研究以短粒粳稻品种南粳35和长粒籼稻N22构建的F2代群体为材料,通过构建遗传连锁图谱挖掘控制水稻谷粒长宽比的QTL,为进一步的遗传研究及育种利用提供参考依据。
1 材料与方法
1.1 材 料
南粳35/N22的F2群体184株及亲本N22和南粳35。
1.2 方 法
1.2.1 材料种植和性状调查 试验材料于2009年正季种植于南京农业大学土桥实验农场,5月17日播种,6月25日移栽,按照16.5 cm×23.5 cm的株行距种植亲本及南粳35/N22的F2群体的184个单株,田间管理同大田生产。成熟期收获亲本和F2单株的种子,自然干燥后每个家系随机选取10粒种子,3次重复,用游标卡尺测量粒长和粒宽,计算每粒种子的长宽比,以3次重复的平均值代表该家系的谷粒长宽比指标。
1.2.2 DNA提取及SSR分析 在分蘖期取亲本及群体内各株系的叶片1~2张提取总DNA。DNA的提取采用微量法,参照 Dellaporta等[4]的方法。待提取的 DNA样品完全溶于 TE缓冲液后,用MBA2000UV/VS光谱仪测定其浓度。用双蒸水将DNA样品稀释成20 ng/μL的工作液,作为PCR扩增反应的模板。PCR 10 μL 反应体系包括:10mmol/L Tris-HCl pH 8.3,50mmol/L KCl,1.5mmol/LMgCl2,50 μmol/L dNTPs,0.2 μmol/L 引物,0.5 U Taq polymerase和 20 ng DNA 模板。扩增反应程序为 95 ℃5 min;94℃ 30 s,55℃ 30 s,72℃ 1min,35个循环;72℃ 7min。扩增产物用8%的非变性聚丙烯酰胺凝胶分离,然后用硝酸银显色,在垩白仪上读取并记录条带。
1.2.3 QTL作图及遗传分析 连锁图谱的绘制由Lu等[5]完成。采用Windows QTL cartographer 2.5软件[6]中的复合区间作图法(CIM)进行标记与性状间的关联分析,在全基因组范围内检测谷粒长宽比位点,及每个QTL可解释表型变异的百分率。将Logarithm of odds(LOD)值2.5定为阈值,若标记区间LOD>2.5,则以该区间LOD值最高处所对应的位点为该性状的1个QTL。按Stuber等[7]方法,根据显性度(DR比值)来判断每个QTL的基因作用方式。当DR≤0.2时,基因效应为加性;当0.2<DR≤0.8时,基因效应为部分显性;当0.8<DR≤1.2时,基因效应为显性;当 DR>1.2时,基因效应为超显性。同时用基于混合线性模型的QTLNetwork 2.0[8]软件,以P=0.005为统计检测阈值,对群体中的互作QTL进行检测。
2 结果与分析
2.1 亲本南粳35与N22及F2群体的谷粒长宽比表现
亲本N22和南粳35的平均谷粒长宽比分别为2.88和2.11,t检测表明亲本谷粒长宽比之间的差异达到极显著水平(P≤0.01)。F2群体的谷粒长宽比介于1.94~3.00,平均为2.46;谷粒长宽比呈双向超亲分离,表现为近正态分布(图1),表明谷粒长宽比是由多基因控制的数量性状,符合QTL作图的要求。
2.2 谷粒长宽比QTL定位
利用Windows QTL cartographer2.5软件在全基因组范围对控制谷粒长宽比的QTL进行检测。在第3、4和5染色体上共检测到3个 QTL(图 2),LOD 值分别为6.10、2.62 和 21.81,贡 献 率 分 别 为5.44%、4.97% 和 58.06%,qLWR-4增效基因来自南粳35外,qLWR-3和qLWR-5增加谷粒长宽比的基因均来自长粒型籼稻品种 N22(表1),其中qLWR-5的LOD和贡献率最大,来自N22的等位基因可以增加0.20的长宽比表型变异,为控制谷粒长宽比的主效QTL;基因作用方式分析表明qLWR-3为超显性、qLWR-4为显性、qLWR-5为部分显性;利用QTLNetwork 2.0软件没有检测到互作QTL。
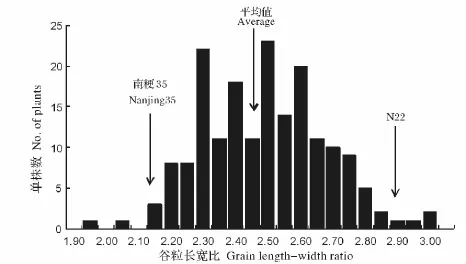
图1 南粳35/N22 F2群体谷粒长宽比分布频率Fig.1 Frequency distribution of grain length-width ratio in Nanjing35/N22 F2 population
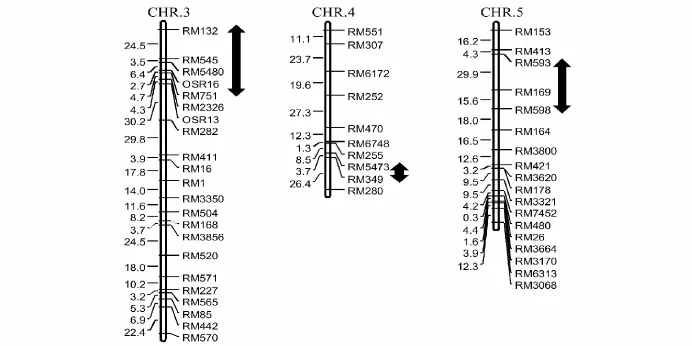
图2 南粳35/N22 F2群体检测的谷粒长宽比QTL在染色体上的分布Fig.2 The location of QTL for grain length-width ratio in Nanjing35/N22 F2 population
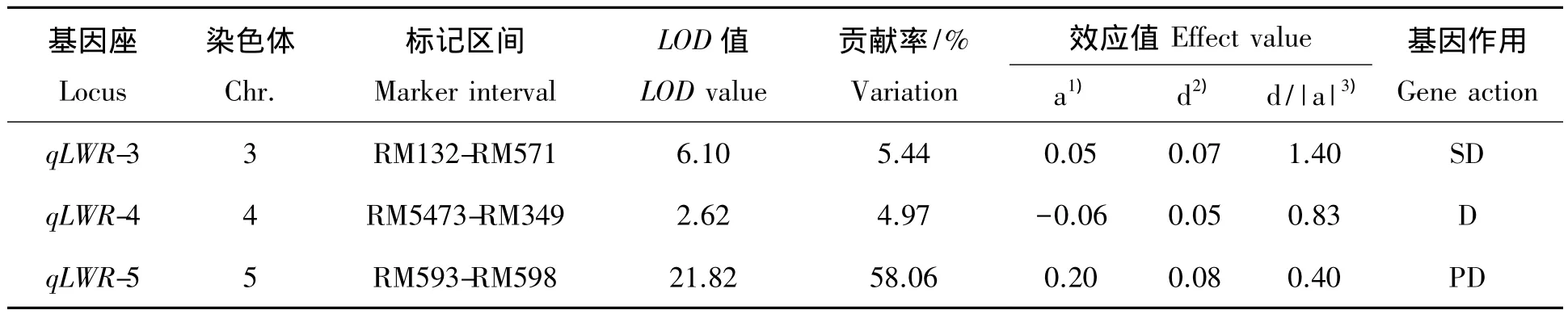
表1 南粳35/N22 F2群体检测的谷粒长宽比QTLTab.1 QTL identified for grain length-w idth ratio in Nanjing35/N22 F2 population
3 结论与讨论
水稻谷粒长宽比可以用来表示谷粒的形状,是衡量稻谷外观品质的重要指标之一。通过不同亲本构建的群体定位结果不尽相同,表明不同水稻品种中控制谷粒长宽比的基因存在差异,研究不同品种长宽比的遗传规律,将有利于充分揭示谷粒长宽比表型变化的大量基因座位,为相关基因分离及进一步克隆提供重要的信息。本研究利用长粒型籼稻品种N22和短粒粳稻品种南粳35构建的F2群体对谷粒长宽比QTL进行定位,共检测到3个控制谷粒长宽比的QTL,qLWR-5 LOD值为21.82,贡献率接近60%,是控制谷粒长宽比的主效QTL,其邻近的分子标记可用于分子标记辅助选择育种。已有研究表明长宽比是多基因控制的数量性状,以加性效应为主,广义遗传率和狭义遗传率均较高[3],本研究中检测到的3个QTL则表现为超显性、显性和部分显性,与前人研究存在差异,可能是亲本遗传背景不同引起。本研究未检测到上位性QTL对谷粒长宽比的影响,与谭耀鹏等[9]研究得出的谷粒外观性状受互作对的影响较小结果是一致的。
qLWR-3位于第3染色体短臂,与已克隆的GS3基因距离较远[10],而与Bai等[11]定位的qLWR3a和陈冰蠕等[12]定位的qlw3a位置相似,且对表型的贡献率均在10%以内,推测应为同一QTL。本研究中qLWR-4定位在第4染色体长臂末端标记RM551和RM518之间,赵明芳等[13]利用单片段代换系在第4染色体长臂末端也检测到一个控制谷粒长宽比的QTL qRLW-4,但与本研究的位置存在一定的偏差,是否属同一位点还需进一步验证。本研究中qLWR-5可以解释58.06%的表型变异,是控制谷粒长宽比的主效QTL,分析发现该区间包含已克隆的控制粒宽基因GW5[14]和控制籽粒大小的基因GS5[15],研究表明GW5编码一个144氨基酸组成的核定位蛋白,通过泛素蛋白酶体途径调节籽粒宽,而GS5编码一个丝氨酸羧肽酶,该基因的高效表达能促进细胞分裂而增加细胞数目,使水稻籽粒增大,因此qLWR-5很可能与GW5或GS5等位,鉴于该位点对粒型的重要性,可在今后的水稻育种中加以利用。
[1]FAO.Statistical databases[DB].Food and Agriculture Organization(FAO)of the United Nations,2004.
[2]王忠华,方振华,干建彗.稻米外观品质性状遗传与分子定位研究进展[J].生命科学,2009,21(3):444-451.
[3]宫李辉,高振宇,马伯军,等.水稻粒形遗传的研究进展[J].植物学报,2011,46(6):597-605.
[4]Dellaporta S L,Wood J,Hicks JB.A plant DNA minipreparation:version II[J].Plant Molecular Biology Reporter,1983,1(4):19-21.
[5]Lu B Y,Xie K,Yang CY,etal.Genetic analysis of two weak dormancymutants derived from strong seed dormancy wild type rice N22(Oryza sativa)[J].Journal of Integrative Plant Biology,2011,53(5):338-346.
[6]Wang SC,Basten C J,Zeng Z B.Windows QTL Cartographer Version 2.5[CP].http://statgen.ncsu.edu/qtlcart/WinQTLCart.htm.
[7]Stuber CW,Lincoln SE,Wolff DW,et al.Identification of genetic factors contributing to heterosis in a hybrid from two elite maize inbred lines usingmolecularmarkers[J].Genetics,1992,132:823-839.
[8]Yang J,Zhu J.Predicting superior genotypes in multiple environments based on QTL effects[J].Theoretical and Applied Genetics,2005,110:1268-1274.
[9]谭耀鹏,李兰芝,李平,等.利用DH群体定位水稻谷粒外观性状的QTL[J].分子植物育种,2005,3(3):314-322.
[10]Fan C C,Xing Y Z,Mao H L,et al.GS3,amajor QTL for grain length and weight and minor QTL for grain width and thickness in rice,encodes a putative transmembrane protein[J].Theor Appl Genet,2006,112:1164-1171.
[11]Bai X F,Luo L J,Yan W H,et al.Genetic dissection of rice grain shape using a recombinant inbred line population derived from two contrasting parents and finemapping a pleiotropic quantitative trait locus qGL7[J].BMC Genetics,2010,11:16.
[12]陈冰蠕,石英尧,崔金腾,等.利用 BC2F2高代回交群体定位水稻籽粒大小和形状QTL[J].作物学报,2008,34(8):1299-1307.
[13]赵明芳,张桂权,曾瑞珍,等.基于单片段代换系的水稻粒型 QTL加性及上位性效应分析[J].作物学报,2011,37(3):469-476.
[14]Weng JF,Gu SH,Wan X Y,etal.Isolation and initial characterization of GW5,amajor QTL associated with rice grain width and weight[J].Cell Res,2008,18:1199-1209.
[15]Li Y B,Fan CC,Xing Y Z,et al.Natural variation in GS5 plays an important role in regulating grain size and yield in rice[J].Nature Genetics,2011,43(12):1266-1269.
