滇西北不同海拔梯度下急尖长苞冷杉叶片的生态解剖结构特性研究
2015-05-28姜永雷冯程程程小毛黄晓霞
唐 探,姜永雷,冯程程,程小毛,黄晓霞
(西南林业大学 园林学院,云南 昆明 650224)
海拔是环境因子的综合体现,各种环境因子在海拔梯度上表现出不同的变化,这为研究植物生理形态对环境的响应提供了良好的条件[1]。在海拔梯度上,各种环境因子如温度、空气湿度、氧分压、水分状况和光照强度等均发生了不同程度上的变化[2]。这些环境因子的改变会对植物叶片结构产生一定的影响。有研究表明,在植物的进化过程中,植物叶片是对环境变化较为敏感的器官,随着环境的变化,其结构特征最能体现环境因子与植物的协同进化[3]。而针叶植物叶型特殊,有特殊的解剖结构,更能体现出环境因子与植物的协同进化[4]。因此,通过对叶片结构特征的研究,分析其在不同海拔梯度上的差异性,是研究植物对长期环境变化适应能力的重要途径。
急尖长苞冷杉(Abies georgei Orr var.Smithii(Viguie et Gaussen)Cheng et L.)又名乌蒙冷杉,是松科(Pinaceae)冷杉属(Abies)长苞冷杉(Abies georgei)的变种,为中国特有植物。据文献报道,急尖长苞冷杉分布于滇西北、川西南、藏东南的海拔2 500~4 000 m的高山地带,模式标本采自云南丽江的玉龙雪山[5]。关于急尖长苞冷杉的研究从20世纪90年代才开始,其研究主要集中在急尖长苞冷杉的群落特征[6]、林隙更新[7]、林隙灌木植被特征[8]、养分循环[9]、树干养分元素含量[10]以及水文特征[11]等方面。而有关海拔梯度对急尖长苞冷杉叶片解剖结构特征影响的研究少见报道。本文选取了滇西北丽江玉龙雪山自然保护区内的急尖长苞冷杉天然林作为试验材料,通过比较分析不同海拔高度上急尖长苞冷杉叶片的解剖结构特征,探究不同海拔高度上急尖长苞冷杉叶片形态结构对环境的响应机制,进而对认识和保护高海拔脆弱的生态系统有一定的借鉴意义,并对揭示高山植物与环境相互作用的复杂关系以及人类通过科学保护和持续利用高山植物资源具有重要的理论和实践意义。
1 材料与方法
1.1 研究地区概况
玉龙雪山位于云南省丽江市玉龙县境内(北纬 27°03′20″~27°40′,东经 100°04′10″~100°16′30″),是中国纬度最南的一座雪山。玉龙雪山南北长约35 km,东西宽约20 km,大小山峰约70余座,主峰扇子陡海拔5 596 m。玉龙雪山自然保护区东界丽鸣公路,西临虎跳峡,南起玉湖,北至大具下虎跳峡口,总面积约26万hm2,天然林面积约7 663 hm2[12]。研究区域位于玉龙雪山自然保护区内,其河谷到山顶的高差悬殊,气候垂直差异大,海拔每上升100 m,其平均气温降低约0.86℃。在整个垂直区域内,由谷到峰顶依次出现暖性、暖湿性、温凉性、北亚热带和寒温性等山地垂直气候带。急尖长苞冷杉分布在海拔3 100~3 800 m,随着海拔的升高,其在群落中的重要值的比重也逐渐增大。在海拔高度超过3 550 m时,急尖长苞冷杉成为优势种群,而少有其他种类。
1.2 研究材料
急尖长苞冷杉叶片采自丽江玉龙雪山自然保护区内的不同海拔(A:3 100 m;B:3 250 m;C:3 400 m;D:3 550 m;E:3 700 m)梯度上。2 013年5月,根据急尖长苞冷杉在玉龙雪山自然保护区内的分布区域,在玉龙雪山3 100~3 800 m 的海拔内,每隔150 m设 A(3 100 m)、B(3 250 m)、C(3 400 m)、D(3 550 m)、E(3 700 m)5个海拔梯度。每个海拔梯度随机选取5个大小为3 m×3 m的样地,样地选择在阳坡和相近坡度,且物种组成及土壤环境条件基本一致,每样地选取胸径大小及长势相近的无病虫害的成年树1株,每海拔梯度共5株进行叶片取样。取样过程中,每株选取树冠下部东、南、西、北4个方向上一年生枝条上的成熟叶片等量若干,且应尽量缩小观察、取样的空间和时间上的差异。取样结束后,立即用0.1 mol/L的磷酸缓冲液(pH 6.8)轻轻除去表面的污物,FAA(50%乙醇∶甲醛38%∶冰醋酸99.5%=90∶5∶5)野外固定并保存。记录相关采样地的详细信息,并带回实验室进行切片制作及叶片解剖结构参数的测定。
1.3 研究方法
1.3.1 切片制作和叶片解剖结构参数的测定 切片制作方法参考李芳兰的方法[13]。在光学显微镜(Nikon ECLIPSE Ti-S,LEICA DM 2000)下观测叶片表皮角质层厚度、上表皮厚度、栅栏组织厚度、海绵组织厚度,栅栏组织细胞的长短径、叶片的长短径、韧皮部厚度、木质部厚度、转输组织厚度、等11项指标,每项指标重复测量5次,取其平均值。
1.3.2 统计分析 所有的数据计算分析都采用SPSS 11.5统计分析软件进行一元方差分析(one-way ANOVA),分析海拔梯度上叶片特征参数的差异,平均数间的多重比较采用 Ducan’s检验方法。0.01≤P≤0.05时为差异显著,P≤0.01 时为极显著。
2 结果与分析
2.1 不同海拔梯度下急尖长苞冷杉叶片解剖形态比较
图1是各个海拔梯度上急尖长苞冷杉叶片解剖结构形态图的比较。切片观察表明,急尖长苞冷杉叶片的结构由表皮组织、叶肉组织和叶脉组织3部分组成。图中可以看出,急尖长苞冷杉叶片上表皮由2层细胞组成,且排列紧密;表皮细胞外壁覆盖着一层角质层,图中海拔3 550 m处的叶片角质层较为明显(较厚些),位于近上表皮细胞部位的栅栏组织细胞呈长柱形,总体排列紧密;但海拔3 100 m和3 400 m处的叶片栅栏组织细胞排列较为疏松且相对较薄;相对于其他海拔梯度上的叶片叶脉组织厚度而言,海拔3 550 m处的叶片叶脉组织稍厚些。因此可以看出,各海拔梯度上复杂的环境因子的差异使得急尖长苞冷杉叶片各形态结构产生了一定的差异,需结合不同海拔叶片解剖结构具体参数(图2、图3、表1)进行更深入的比较,分析其叶片形态结构对环境的响应机制。
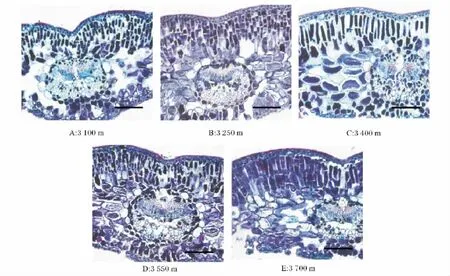
图1 不同海拔急尖长苞冷杉叶片解剖形态比较Fig.1 A comparative study on anatomic form at gradient elevation of Abies georgei var.smithii leaves
2.2 海拔梯度对急尖长苞冷杉叶片表皮组织结构的影响
通过对不同海拔下急尖长苞冷杉叶片表皮组织结构参数(图2)比较发现,叶片表皮角质层厚度的变化范围在(7.52±1.29)~(8.57±0.6)μm。随着海拔的升高,叶片表皮角质层的厚度呈现出先增大后减小的趋势,并且在海拔3 550 m附近达到最大值(8.57±0.61)μm。但是在各个海拔梯度上的差异并不显著;在海拔3 550 m以下时,叶片上表皮厚度呈现出增大的趋势,海拔平均每升高150 m,其叶片上表皮厚度平均增加5.06μm,且在海拔3 550m处增厚显著,达到(44.14±4.35)μm。当海拔超过3 550 m时,其上表皮厚度显著降低。在各海拔梯度上,急尖长苞冷杉叶片的上表皮厚度最大相差 1.5 倍。
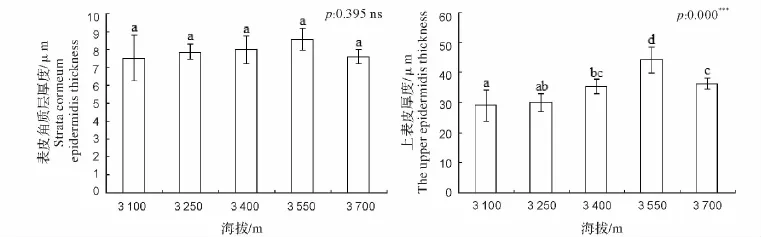
图2 不同海拔急尖长苞冷杉叶片表皮组织结构参数比较Fig.2 A comparative study on epidermal tissues at gradient elevation of Abies georgei var.smithii leaves
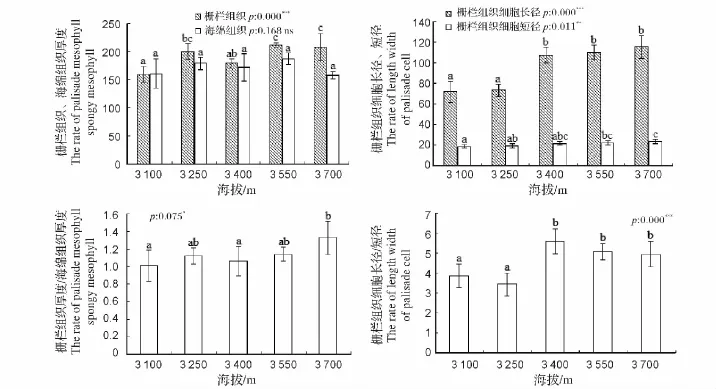
图3 不同海拔急尖长苞冷杉叶片叶肉组织结构参数比较Fig.3 A comparative study on mesophyll tissues at gradient elevation of Abies georgei var.smithii leaves
2.3 海拔梯度对急尖长苞冷杉叶片叶肉组织结构的影响
图3显示了急尖长苞冷杉叶片叶肉组织结构参数在不同海拔梯度下的差异,通过比较发现,叶肉组织中栅栏组织和海绵组织的厚度分别在(158.68±14.32)~ (212.03±3.72)μm 与(157.60±5.95)~(186.90±9.90)μm,并且,随着海拔的上升,两者的厚度呈现出相似的变化。在海拔3 550 m附近,栅栏组织和海绵组织的厚度分别达到最大值。在各海拔梯度间,栅栏组织厚度的变化呈现出显著的差异,而海绵组织厚度的变化不显著,这可能是由于栅栏组织对海拔梯度上环境因子的变化更加敏感造成的。在海拔上升的的过程中,栅栏组织与海绵组织厚度的比值(栅栏系数)总体上呈上升的趋势,且当海拔超过3 400 m时,其比值显著上升;图3中,栅栏组织细胞长短径均随着海拔的上升而呈现出增大的趋势,其中栅栏组织细胞长径在海拔3 250~3 400 m时变化明显,而其短径在整个海拔梯度上的变化都比较显著。栅栏组织细胞长短径的比值(栅栏细胞系数)总体上呈上升趋势,在海拔3 400 m时,比值上升显著,而在海拔3 400 m以上的情况下,则无明显变化。
2.4 海拔梯度对急尖长苞冷杉叶片叶脉组织结构及长短径比值的影响
表1是不同海拔下急尖长苞冷杉叶片叶脉组织结构参数的比较。在整个海拔梯度上,随着海拔的上升,韧皮部厚度、木质部厚度和转输组织厚度的变化趋势相似,总体上呈现出先增大后减小的趋势,并在海拔3 550 m 附近达到最大值,其厚度分别为:(51.07±3.37)μm、(46.09±4.09)μm、(69.04±5.32)μm。在海拔3 550 m以下,海拔平均每升高150 m,韧皮部、木质部和转输组织厚度相应增大7.81%、14.44%和3.24%。在相邻的海拔梯度上,韧皮部和木质部的厚度变化总体上差异显著,而转输组织的厚度变化不明显。
在海拔3 400 m以上,叶片长短径的比值随着海拔的上升而增大(表1),且在3 700 m处显著增大。这可能是因为海拔升高导致温度迅速下降,使得细胞原生质透水性降低,叶片长短径比值的增大,有利于叶绿体垂直叶表排列和充分采集光照,大大提高了光合效率以及植物对水分的利用率[14]。
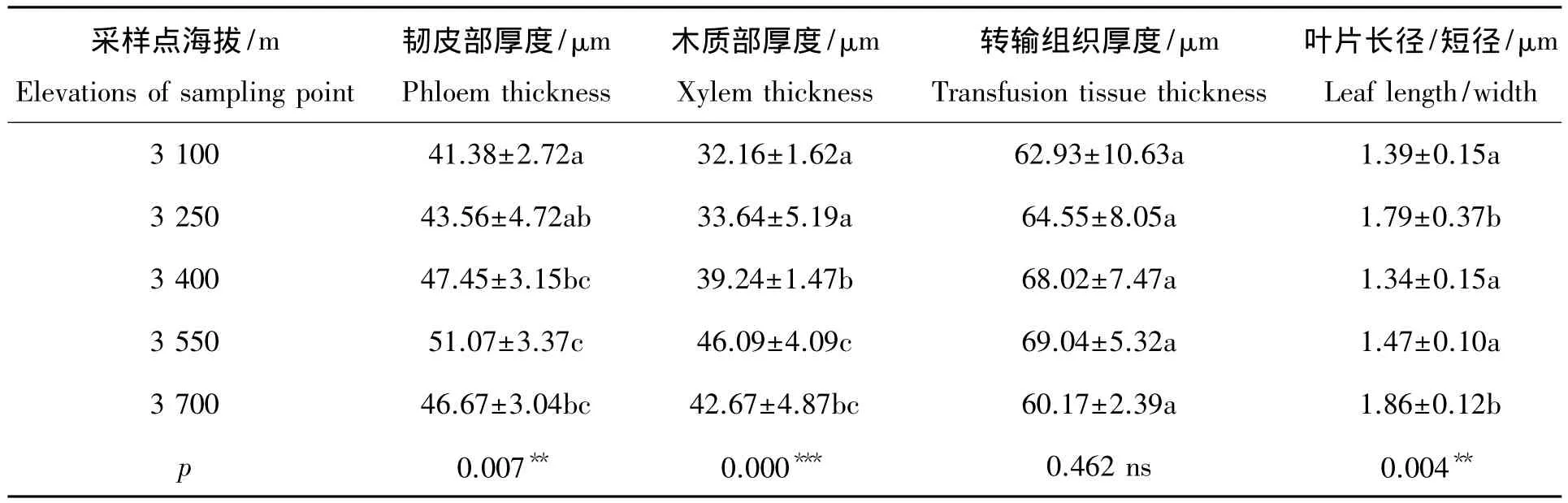
表1 不同海拔急尖长苞冷杉叶片叶脉组织结构参数及长短径比值的比较Tab.1 A comparative study on vein tissues and leaf length/w idth at gradient elevation of Abies georgei var.smithii leaves
3 讨论与结论
植物在生长发育过程中受到内在因子和外在环境的影响,同一种植物在不同的环境中生长,也会表现出很大的差异性。在不同的海拔高度上,其环境因子会发生不同程度上的变化。随着海拔的升高,温度和水分会逐渐限制植物的生长发育,尤其是叶片对环境的变化反应更为敏感[15]。当环境中温度、水分、光照强度等环境因子发生变化时,叶片的外部形态和内部解剖结构也会产生细微的差异[16]。长期生长在玉龙雪山上的急尖长苞冷杉,其叶片形态结构与所在海拔的环境之间往往表现出高度的统一。
植物叶片的表皮细胞和表皮角质层具有防止水分过度蒸腾,抵抗外界风雨和病虫侵害的作用[17]。当海拔在3 550 m以下时,急尖长苞冷杉表皮角质层厚度和上表皮厚度与海拔高度呈正相关,这可能是由于海拔上升,昼夜温差就会逐步加大,氧气和二氧化碳分压降低,叶片表皮角质层和上表皮厚度的增大,可以提高叶片表面的结构强度,抵御高海拔气流变化和强辐射造成的伤害[18]。在海拔超过3 550 m时,叶片的上表皮厚度显著减小,且角质层厚度也有减小的趋势,这说明急尖长苞冷杉种群分布的正面影响逐渐减弱。
叶肉是植物叶片光合作用的主要部位,植物的光合作用效率会受到栅栏组织和海绵组织厚度、栅栏细胞形态等各方面变化的影响[19]。急尖长苞冷杉叶片的栅栏组织和海绵组织厚度总体上呈先增大后减小的趋势,但栅栏组织厚度的变化较海绵组织更为显著。这说明栅栏组织对海拔梯度上环境因子的变化更为敏感,这与李芳兰[20]关于四川黄栌(Cotinus szechuanensis)叶片的生态解剖研究结果类似。在海拔3 550 m以下,随着海拔的升高,栅栏组织和海绵组织的厚度总体呈增大的趋势,说明海拔上升时,温度下降而降水量增加,良好的水分条件促使叶片的生长和厚度的增大,使得植物能够适应湿冷的环境并起到保温效果,而且可以有效抵御强辐射给叶肉细胞带来的伤害。栅栏组织厚度的明显增大,可以增加光合作用,提高光合效率,弥补叶片狭小带来的负面效应[21]。栅栏组织和海绵组织厚度比值(栅栏系数)与海拔梯度呈正相关,说明高海拔上急尖长苞冷杉叶片已经形成了防止水分过多散失的适应性结构特征,李杰等[22]对不同海拔高度胡枝子(Lespedeza bicolor)叶片解剖结构特征的研究中也得出了相似的结论。Cald等[23]认为,当海拔升高时,短波辐射会不断增强,使得叶片栅栏组织细胞产生保护性的稳定遗传结构,能够防止叶面温度升高引起的组织灼伤。本研究中,高海拔的急尖长苞冷杉叶片栅栏细胞系数(栅栏组织细胞长短径比)较大,说明叶片栅栏细胞建立起了叶肉细胞与外界环境的一道屏障,抵御短波辐射带来的伤害。这在戴怡龄等[24]的橘黄罂粟(Papaver croceum)叶片结构特征研究和安黎哲等[25]的火绒草(Leontopodium leontopodiodes)叶片结构特征研究中也有所报道。但与陈晓莉[26]关于祁连圆柏(Sabina przewalskii)叶片的研究结果相反,这可能是由于祁连圆柏特殊的生物学特性和关键性限制因子的差异造成的。
叶脉作为叶的输导组织和支撑结构,可以为叶片提供水分和无机盐、输出光合产物等,保证叶的生理功能顺利进行。急尖长苞冷杉叶片叶脉中的韧皮部厚度、木质部厚度和转输组织厚度随着海拔梯度的上升而呈现出先增大而减小的趋势,当海拔高度超过3 550 m时,韧皮部和木质部厚度下降明显,这可能是因为高海拔低温条件下,植物叶片的生长季节缩短,细胞生长缓慢,导致光合作用的产物消耗较少,使得输送有机质的韧皮部厚度和输送无机盐和水分的木质部厚度变薄[27]。
综合分析表明,由于玉龙雪山不同海拔梯度上环境因子的改变,导致了急尖长苞冷杉叶片解剖结构不同程度上的差异性,充分体现出其适应生态环境的多样性。表现为:当海拔在3 550 m以下时,急尖长苞冷杉叶片表皮角质层厚度、上表皮厚度、栅栏组织厚度、海绵组织厚度、栅栏组织细胞长短径、栅栏系数、栅栏细胞系数、韧皮部厚度、木质部厚度及转输组织厚度随着海拔的升高,总体上呈增加趋势,并逐渐进入适合生长的区域。3 550 m海拔附近为玉龙雪山中急尖长苞冷杉生长最适宜的环境。当海拔高于3 550 m时,急尖长苞冷杉叶片表皮角质层厚度、上表皮厚度、栅栏组织厚度、海绵组织厚度、韧皮部厚度、木质部厚度及转输组织厚度明显减小,体现了急尖长苞冷杉对高海拔地区生存环境的适应,以延长和保存生命。
[1]王勋陵,王静.植物形态结构与环境[M].兰州:兰州大学出版社,1989:105-148.
[2]王为义.高山植物结构特异性的研究[J].高原生物学集刊,1985,4(4):19-34.
[3]李芳兰,包维楷.植物叶片形态解剖结构对环境变化的响应与适应[J].植物学通报,2005,22(增刊):118-127.
[4]张明明,高瑞馨.针叶植物叶片比较解剖及生态解剖研究综述[J].森林工程,2012,28(2):9-10.
[5]中国科学院中国植物志编辑委员会.中国植物志[M].北京:科学出版社,1978:78.
[6]任青山,王景升,张博,等.藏东南冷杉原始林不同形态水的水质分析[J].东北林业大学学报,2002,30(2):52-54.
[7]罗大庆,郭泉水,薛会英,等.藏东南亚高山冷杉林林隙特征与干扰状况研究[J].应用生态学报,2002,13(7):777-780.
[8]罗大庆,郭泉水,边巴多吉,等.西藏色季拉山冷杉原始林林隙更新研究[J].林业科学研究,2002,15(5):564-569.
[9]辛学兵,翟明普.西藏色季拉山冷杉林生态系统的养分循环[J].林业科学研究,2003,16(6):668-676.
[10]卢杰,辛学兵,罗大庆,等.急尖长苞冷杉林树干养分元素含量研究[J].林业科学研究,2003,16(6):44-47.
[11]王景升,任青山,兰小中.急尖长苞冷杉原始森林降水分配格局[J].林业科技,2002,27(6):7-10.
[12]杨少华,薛润光,陈翠,等.滇西北玉龙雪山生物多样性现状及其保护对策[J].西南农业学报,2008,21(3):863-869.
[13]李芳兰,包维楷,刘俊华,等.岷江上游干旱河谷海拔梯度上白刺花叶片生态解剖结构特征研究[J].应用生态学报,2006,17(1):5-10.
[14]Cald M M.Solar UV irradiation and the growth and developmentof higher plants[J].Photo-physiology,1993,1(6):125-143.
[15]祁建,马克明,张育新.辽东栎叶特性沿海拔梯度的变化及其环境解释[J].生态学报,2007,27(3):931-937.
[16]刘全宏,王孝安,田先华,等.太白红杉叶的形态解剖学特征与环境因子的关系[J].西北植物学报,2001,21(5):885-893.
[17]贺学礼.植物学[M].西安:陕西科学技术出版社,2001:54-55.
[18]李有忠.海拔高度的变化对植物叶片内部结构的影响[J].青海师范大学学报:自然科学版,1995(4):34-40.
[19]Li F L,Bao W K,Liu JH.Eco-anatomical characteristics of Sophora davidii leaves along an elevation gradient in upper Minjiang River dry valley[J].Chinese Journal of Applied Ecology,2006,17(1):6-8.
[20]李芳兰,包维楷,吴宁.岷江上游干旱河谷海拔梯度上四川黄栌叶片的生态解剖[J].应用与环境生物学报,2007,13(4):486-491.
[21]蔡永立,达良俊.亚热带东部壳斗科6种常绿植物叶的生态解剖[J].应用与环境生物学报,2002,8(5):460-466.
[22]李杰,贺学礼,李景侠.河北小五台山不同海拔高度胡枝子叶片解剖结构特征的比较[J].河北大学学报:自然科学版,2007,27(2):184-187.
[23]李广民,王秀杰.中国植物学会60周年年会学术报告及论文摘要汇编[C].北京:中国科技出版社,1993:190.
[24]戴怡龄,安黎哲,陈拓,等.寒区不同海拔橘黄罂粟叶片结构特征的比较研究[J].西北植物学报,2004,24(3):495-503.
[25]安黎哲,张满,刘艳红,等.乌鲁木齐河源区不同海拔的火绒草叶片结构特征的比较研究[J].冰川冻土,2004,26(4):474-481.
[26]陈晓莉.不同海拔青海云杉和祁连圆柏生理生态适应性研究[D].兰州:甘肃农业大学,2008.
[27]张立杰,蒋志荣.青海云杉种群分布格局沿海拔梯度分形特征的变化[J].西北林学院学报,2006,21(2):64-68.
