水稻不同品种幼苗期耐盐性评价
2015-05-15崔丰磊张帆涛罗向东谢建坤
周 毅,崔丰磊,杨 萍,张帆涛,罗向东,谢建坤*
(1.江西师范大学 生命科学学院,江西 南昌 330022;2.江西师范大学 高等研究院,江西 南昌 330022)
水稻是世界上最重要的粮食作物之一,全球大约60%的人口以水稻作为主要的粮食来源[1]。水稻属于甜土植物,土壤盐渍化是影响水稻产量提高的主要限制因素之一[2]。据估算,在我国受盐侵害的稻田占栽培面积的1/5,并且随着人口剧增、城市化及工业化的高速发展,有扩大和蔓延的趋势[3]。因此,筛选和培育耐盐性强的水稻品种已成为农业可持续发展的一个重要研究方向。目前,人们主要利用转基因技术以及耐盐突变体筛选等方法来提高水稻的耐盐性[2,4-10],但上述方法周期漫长,操作繁琐复杂,实际应用效果也不尽理想,因此难以得到推广。此外,水稻耐盐性是一个受多种因素综合影响的复杂数量性状,机理复杂,表型性状是内部生理生化发生变化的表现。本研究通过测定NaCl盐胁迫下7个水稻品种幼苗生长和生理生化特性的变化来探明盐处理对水稻苗期生长的影响,并且通过隶属函数法对供试水稻材料的耐盐性进行了综合评价,从而为耐盐种质资源筛选及育种提供理论依据。
1 材料与方法
1.1 实验材料
供试材料:协青早B(以下简称协B),福伊B,9311,紫稻,R463,02428,日本晴等7个水稻品种,种子由江西省农科院提供。
1.2 实验方法
1.2.1 水稻幼苗培养 供试材料种子用1 g/L HgCl2表面消毒10 min,蒸馏水冲洗干净,再用蒸馏水浸种2 d。发芽7 d后,选取大小一致的幼苗移栽到盛有蛭石的塑料盆中,以IRRI营养液[11]进行培养,每隔3 d换一次营养液。
1.2.2 盐胁迫处理 在水稻幼苗移栽大约14 d(三叶一心期)后以200 mmol/L NaCl胁迫处理5 d。试验采用3次重复,每品种每个重复种植10株。对照组则是营养液中不加入NaCl,其它培养条件都相同。
1.2.3 株高、根长和含水量的测定 株高测量从植株底部开始,一直到叶尖。根长测量从植株底部开始,一直到根尖。将幼苗去胚乳后,用电子天平称量鲜质量,再将幼苗在120℃的烘箱中烘30 min,然后置于80℃烘箱中,直至幼苗质量恒定记为干质量。利用鲜干质量差计算含水量。
1.2.4 各项生理生化指标的测定 叶绿素含量测定采用浸提法[12]。电导率的测定按照汪月霞等[13]方法进行测定。可溶性蛋白质含量的测定参照李大红[14]的方法进行测定。脯氨酸含量采用磺基水杨酸法进行测定[15]。MDA含量采用硫代巴比妥酸显色法[14]进行测定。SOD活性的测定采用的是碧云天公司的总SOD活性检测试剂盒(产品编号S0109)。
1.2.5 不同水稻品种耐盐性评价 参照张国新等[16]的隶属函数法对7个不同水稻品种的耐盐性进行鉴定。试验数据统计分析和作图由Excel完成。
2 结果与分析
2.1 NaCl处理对水稻不同品种幼苗株高、根长的影响
在200 mmol/L NaCl处理的盐胁迫中,各个品种的水稻幼苗生长都受到了一定程度的抑制(图1、图2)。就株高而言,受盐胁迫影响最为显著的是紫稻,与对照相比株高下降了30.0%,影响最小的是日本晴,下降率只有0.5%。而根长受盐胁迫影响最为明显的也是紫稻,下降率达到43%,而受影响最小的是9311,与对照相比,根长提高了0.56%。

图1 NaCl处理对不同水稻品种幼苗株高的影响Fig.1 Effect of NaCl stress on the plant height of different rice varieties during seedling stage
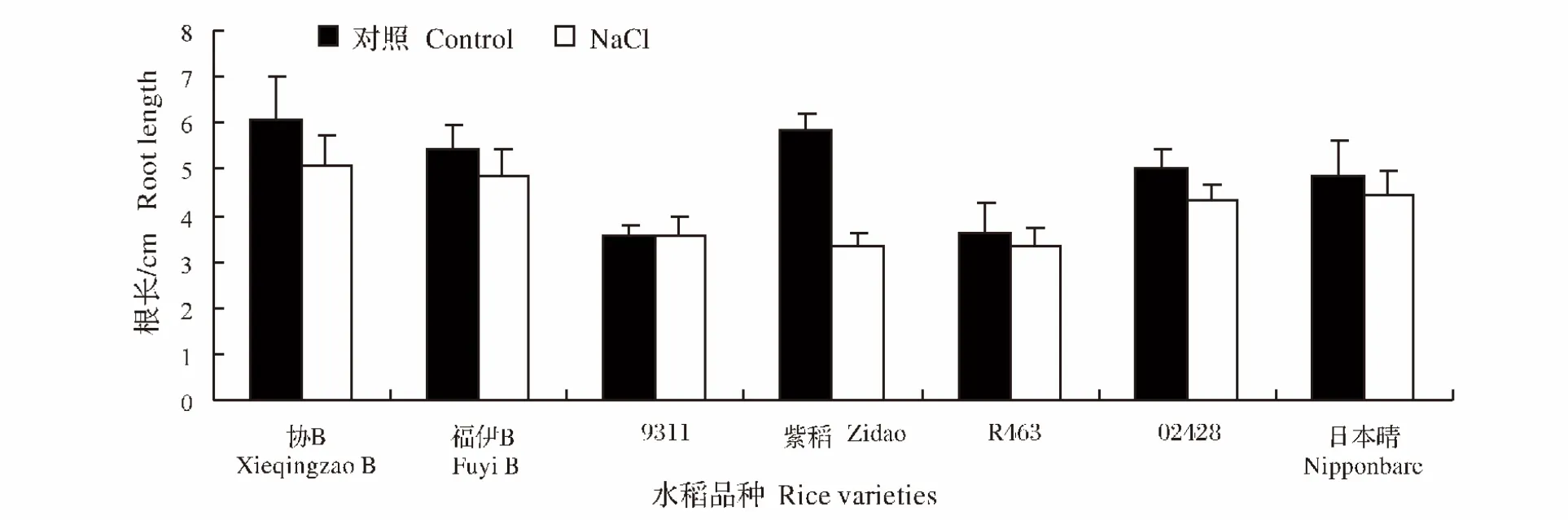
图2 NaCl处理对不同水稻品种幼苗根长的影响Fig.2 Effect of NaCl stress on the root length of different rice varieties during seedling stage
2.2 NaCl处理对水稻不同品种幼苗含水量的影响
植株相对含水量是衡量植物耐逆性的一个重要指标。盐胁迫处理后,各个品种水稻幼苗相对含水量呈降低趋势。其中下降最显著的是紫稻,下降率为52.0%,而9311下降最小,下降率为17.6%(图3)。
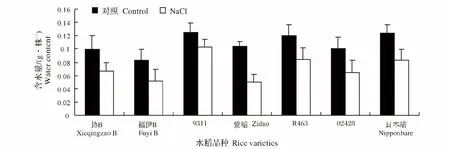
图3 NaCl处理对不同水稻品种幼苗含水量的影响Fig.3 Effect of NaCl stress on the water content of different rice varieties during seedling stage
2.3 NaCl处理对水稻不同品种幼苗叶绿素含量的影响
盐胁迫处理后,各个品种水稻幼苗的叶绿素含量大部分呈降低趋势。其中下降最显著的是协B,下降率为62.9%,而R463的叶绿素含量提高了1.5%(图4)。
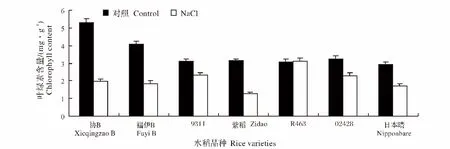
图4 NaCl处理对不同水稻品种幼苗叶绿素含量的影响Fig.4 Effect of NaCl stress on the chlorophyll content of different rice varieties during seedling stage
2.4 NaCl处理对水稻不同品种幼苗细胞质膜透性的影响
细胞质膜是细胞与环境之间的界面,各种逆境对细胞的影响首先作用于质膜,逆境胁迫对质膜结构和功能的影响通常表现为选择透性的下降甚至丧失、电解质与某些小分子有机物质的大量外渗。盐胁迫处理后,各个品种水稻幼苗相对电导率均呈增加趋势。其中提高最显著的是9311,提高率为275.4%,而协B增加最小,提高率为103.8%(图5)。
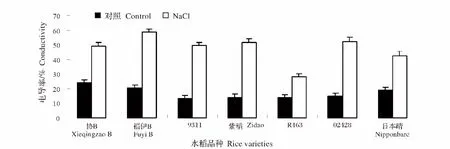
图5 NaCl处理对不同水稻品种幼苗相对电导率的影响Fig.5 Effect of NaCl stress on the conductivity of different rice varieties during seedling stage
2.5 NaCl处理对水稻不同品种幼苗可溶性蛋白含量的影响
植物体内蛋白质质量分数增加是植物在逆境的适应表现,可作为鉴定植物相对抗性的指标。盐胁迫处理后,协B、福伊B和9311幼苗的可溶性蛋白含量发生了显著的提高,提高率分别为48.7%、20.4%以及12.6%,而紫稻的可溶性蛋白含量下降了8.2%,其它品种的含量并没有发生显著变化(图6)。
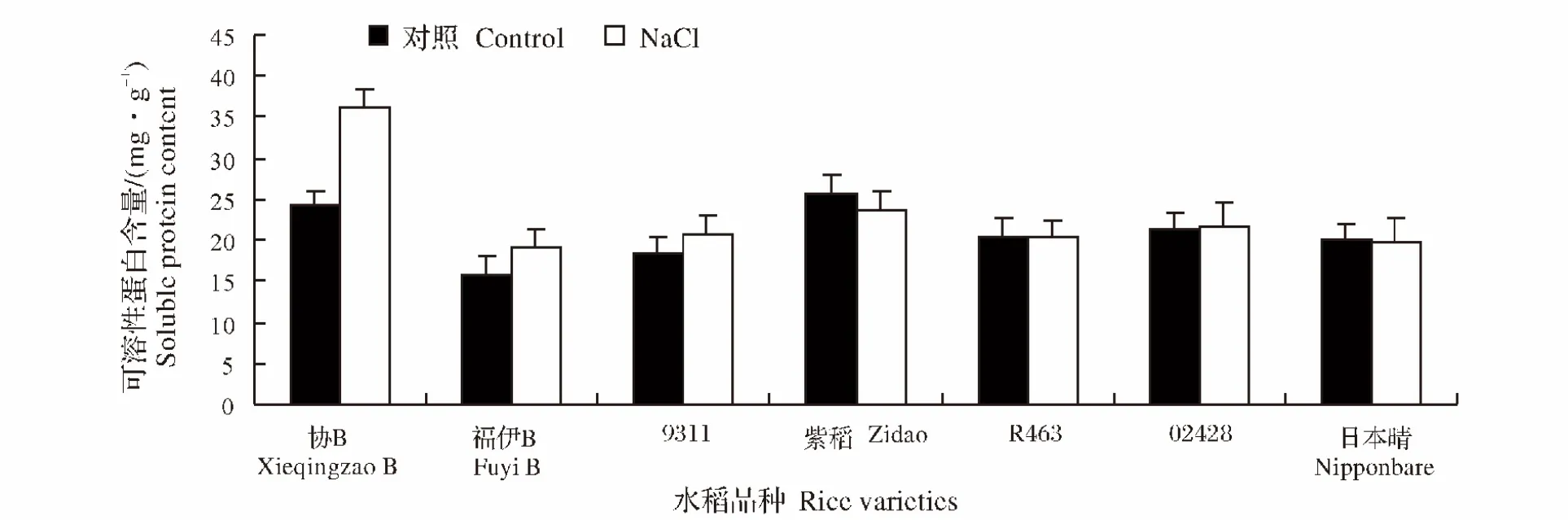
图6 NaCl处理对不同水稻品种幼苗可溶性蛋白含量的影响Fig.6 Effect of NaCl stress on the soluble protein content of different rice varieties during seedling stage
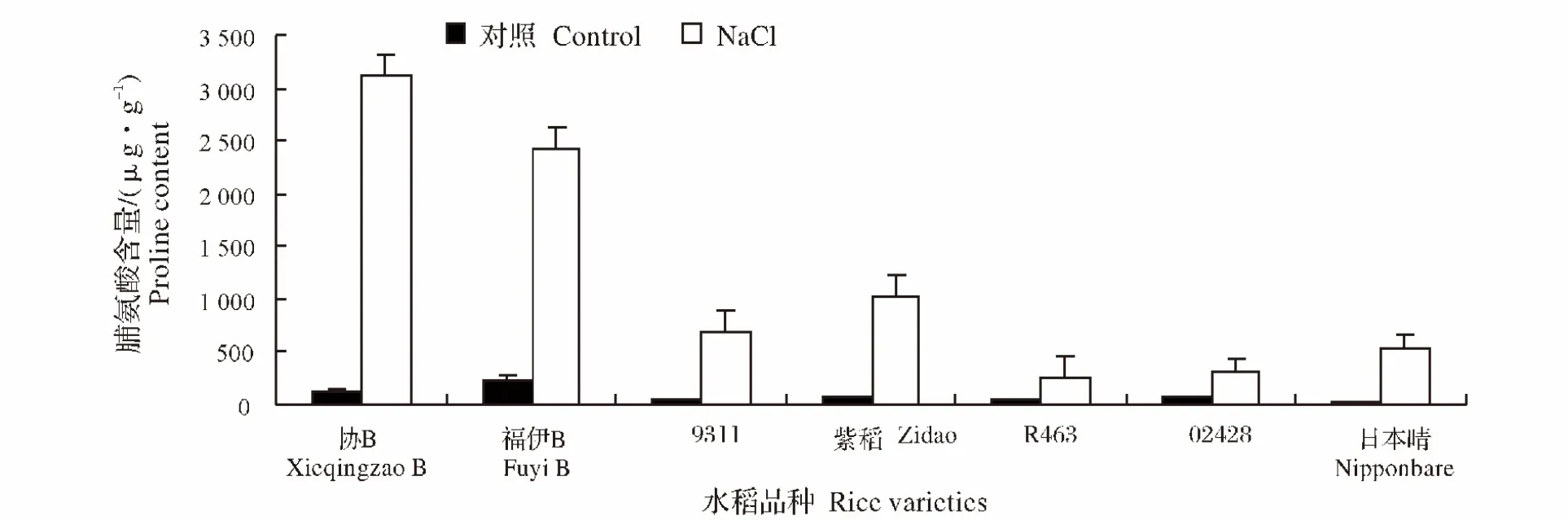
图7 NaCl处理对不同水稻品种幼苗脯氨酸含量的影响Fig.7 Effect of NaCl stress on the proline content of different rice varieties during seedling stage
2.6 NaCl处理对水稻不同品种幼苗脯氨酸含量的影响
脯氨酸是植物体内一种小分子的渗透物质,当植物受到盐胁迫时,脯氨酸的累积能提高植物的耐盐性。在盐胁迫下,各个品种水稻幼苗脯氨酸含量均明显高于对照组,其中协B的脯氨酸含量提高的最为显著,提高率为2 308.1%,而02428的提高程度最小,提高率为326.5%(图7)。
2.7 NaCl处理对水稻不同品种幼苗MDA含量的影响
MDA是膜脂过氧化的主要产物,其量的变化可作为衡量逆境胁迫对植物造成氧化损害的指标。盐胁迫下的水稻幼苗叶片的MDA含量均呈明显上升趋势。其中,日本晴的MDA含量与对照组相比增加了312.0%,而02428的增加的最少,增加率为53.3%(图8)。
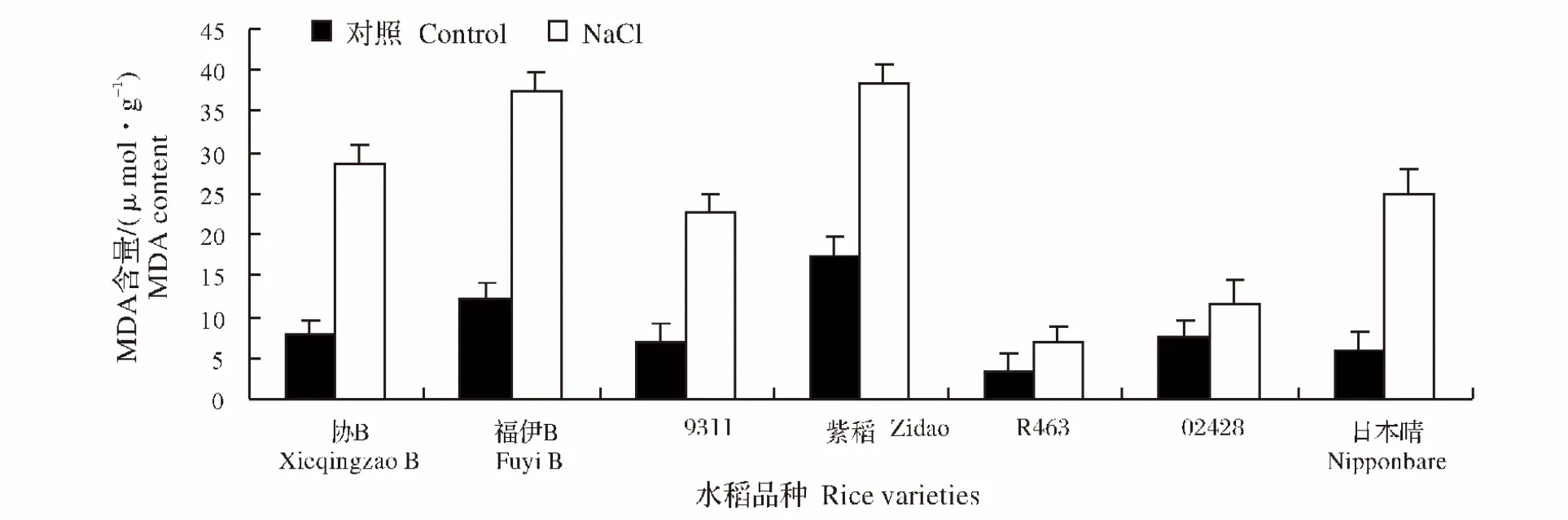
图8 NaCl处理对不同水稻品种幼苗MDA含量的影响Fig.8 Effect of NaCl stress on the MDA content of different rice varieties during seedling stage
2.8 NaCl处理对水稻不同品种幼苗SOD活性的影响
SOD是重要的活性氧清除酶,当外来胁迫导致大量活性氧产生时,它能及时有效清除自由基,保护细胞免受活性氧胁迫的伤害。盐胁迫下的水稻幼苗的SOD活性与对照组相比发生了不同程度的增加。其中,协B的SOD活性提高的最为显著,提高率为943.7%,而紫稻的提高程度最小,提高率为 2.0%(图 9)。
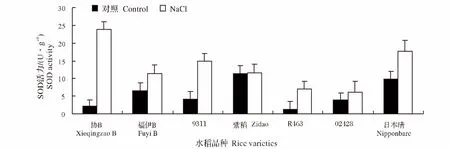
图9 NaCl处理对不同水稻品种幼苗SOD活性的影响Fig.9 Effect of NaCl stress on the SOD activity of different rice varieties during seedling stage
2.9 水稻不同品种幼苗耐盐性综合评价
利用隶属函数计算方程,对9个生长及生理生化指标的变化率(表1)进行数量转换,量化结果见表2。水稻耐盐性是一个多因素作用的结果,因此单一指标很难真正反映品种的耐盐性,而各种生长及生理生化指标的综合评价更能揭示品种真实耐盐力。各指标平均值最大的为协B,其值为0.683 9;其次为R463,其值为0.592 6;紫稻最小,仅为0.143 4;其它品种在0.352 6 ~0.558 4。从大到小依次依次为:协B、R463、9311、日本晴、福伊B、02428、紫稻(表2)。说明协B耐盐性最强,紫稻耐盐性最弱。
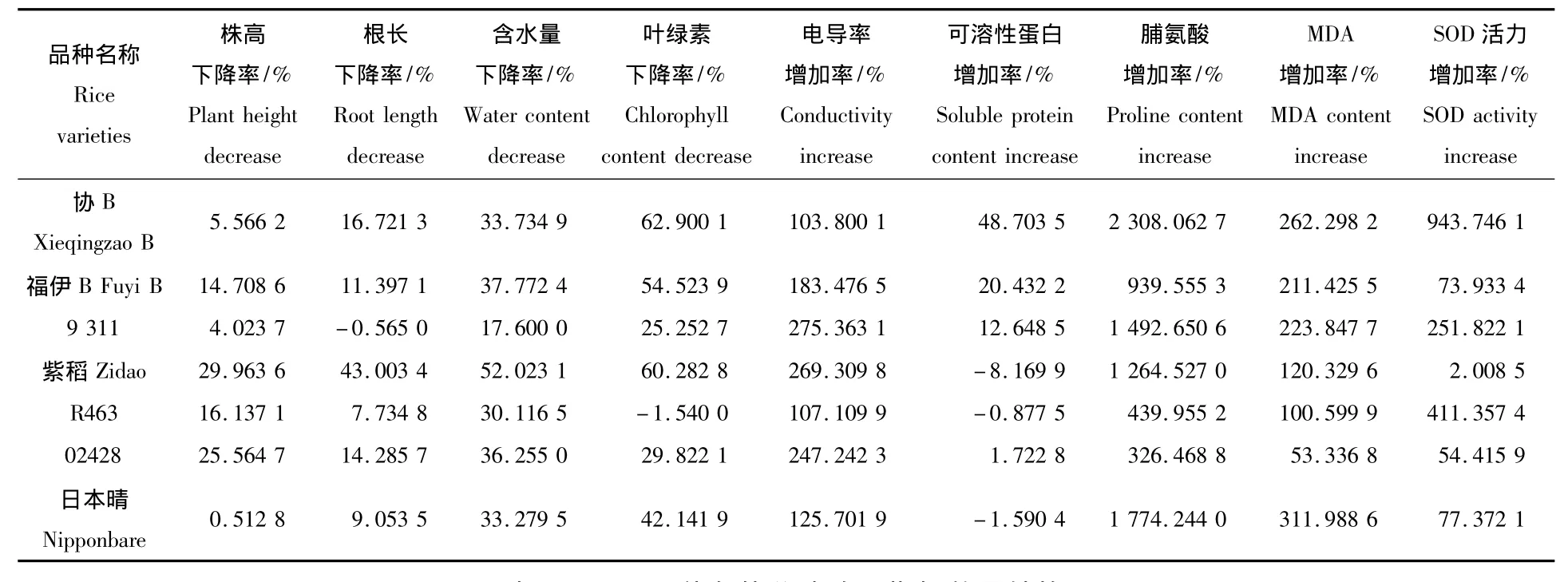
表1 各项生长及生理生化指标变化率Tab.1 The rate of change in the growth and physiological and biochemical characteristics

表2 不同品种在盐分胁迫下指标数量转换Tab.2 The index conversion of different rice varieties under salt stress
3 讨论
水稻耐盐性是一种复杂的生理特性,目前对水稻耐盐性的评价主要通过表型指标和生理生化指标进行[17]。盐胁迫处理对植物的生长发育有明显的抑制作用。研究发现NaCl胁迫处理对水稻以及甜椒幼苗的株高、根长都有显著的影响[18-19]。本研究结果表明,经过5 d的200 mmol/L NaCl处理后,7个水稻品种幼苗表现出不同程度的株高变矮、根长变短(除9311以外),说明水稻幼苗生长受到抑制。
叶片相对含水量是植物叶片细胞水分生理状态的反映,在一定程度上反映了植物组织水分亏缺程度。本实验结果表明,盐胁迫处理后,各个品种水稻幼苗的叶片含水量均发生了不同程度的降低,这个研究结果与金美芳等[18]的研究结果是一致的,其机理可能是盐胁迫导致水稻幼苗内离子的过量积累,造成外界溶液的水势降低,产生渗透胁迫引起细胞脱水,水稻组织出现生理性缺水。
叶绿素含量是体现叶片衰老程度的指标之一,很多研究表明,各种细胞器中受盐分影响比较敏感的是叶绿素。盐胁迫下植物叶片中叶绿素含量下降,可能是由于叶绿素酶活性增强,促使叶绿素分解[20]。盐胁迫下植物体内叶绿素含量的变化较为复杂,本试验结果表明,盐胁迫处理后,除R463幼苗的叶绿素含量无明显变化外,其余6个品种的水稻幼苗的叶绿素含量都发生了明显的下降。下降程度由高到低依次为协 B、紫稻、福伊 B、日本晴、02428、9311、R463。
研究发现,细胞膜是逆境对植物伤害的原初位点,尤以质膜的伤害最为严重。质膜在受到盐胁迫后透性增大和膜脂过氧化,从而损害膜的正常生理功能,进而影响细胞的代谢作用,最终导致植物代谢过程发生紊乱[21]。本研究中,盐胁迫下各个品种水稻幼苗的电导率都发生了明显的增加,说明NaCl处理后,水稻幼苗叶片的选择透性明显降低,电解质以及某些小分子有机物质发生大量外渗,从而影响幼苗的生长发育。
研究发现,渗透调节是植物适应盐胁迫的基本特征之一,盐胁迫可提高水稻幼苗可溶性蛋白和脯氨酸等的含量[22]。可溶性蛋白含量与植物的抗逆性之间存在正相关,植物抗逆性的提高可能涉及特异可溶性蛋白质形成[14]。本研究结果表明,盐胁迫处理后,只有部分品种(协B、福伊B、9311)水稻幼苗的可溶性蛋白含量提高了,具体机理还有待进一步研究。Hichri等[23]的研究结果表明,盐胁迫条件下,植物体内大量积累脯氨酸。本研究结果表明,在盐胁迫条件下,各个品种水稻幼苗的脯氨酸含量都发生了明显的增加,说明脯氨酸作为渗透溶质增强植物的抗逆性。
MDA是细胞膜质过氧化产物,它的积累可作为植物叶片细胞膜受破坏程度的指标。此外,MDA含量与植物耐盐性密切相关,许多研究表明,耐盐的植物MDA含量比较低[24]。本研究结果表明,在盐胁迫条件下,7个水稻品种幼苗的MDA含量都有大幅度的上升,说明各个水稻品种的细胞膜均受到一定程度的伤害。
许多植物抗逆研究已经表明SOD酶活性与植物抗逆性有极为密切的关系[25]。本研究结果表明,在盐胁迫条件下,除紫稻外,其余6个品种的水稻幼苗的SOD活性都发生了显著的提高。水稻耐盐性是一个多因素作用的结果,机理复杂,隶属函数分析提供了一条在多指标测定基础上对材料特性进行综合评价的途径,将其应用于水稻耐盐育种的研究,可大大提高水稻耐盐性筛选的可靠性[16]。本研究利用隶属函数法对各项生长及生理生化指标进行综合评价,发现在所试验的7个水稻品种中协B是相对比较耐盐的。
[1]Xu D Q,Huang J,Guo S Q,et al.Overexpression of a TFIIIA-type zinc finger protein gene ZFP252 enhances drought and salt tolerance in rice(Oryza sativa L.)[J].Febs Lett,2008,582(7):1037-1043.
[2]Schmidt R,Mieulet D,Hubberten H M,et al.Salt-responsive ERF1 regulates reactive oxygen species-dependent signaling during the initial response to salt stress in rice[J].Plant Cell,2013,25(6):2115-2131.
[3]刘少华,王仁雷,刘青,等.NaCl预处理对高盐胁迫下两系杂交稻幼苗生长的影响[J].河南农业大学学报,2013(2):128-131.
[4]Sun L,Zhang Q,Wu J,et al.Two rice authentic histidine phosphotransfer proteins,OsAHP1 and OsAHP2,mediate cytokinin signaling and stress responses in rice[J].Plant Physiol,2014,165(1):335-345.
[5]Wang S,Uddin M I,Tanaka K,et al.Maintenance of chloroplast structure and function by overexpression of the rice MONOGALACTOSYLDIACYLGLYCEROL SYNTHASE gene leads to enhanced salt tolerance in tobacco[J].Plant Physiol,2014,165(3):1144-1155.
[6]Ghosh A,Pareek A,Sopory S K,et al.A glutathione responsive rice glyoxalase II,OsGLYII-2,functions in salinity adaptation by maintaining better photosynthesis efficiency and anti-oxidant pool[J].Plant J,2014,80(1):93-105.
[7]Serra T S,Figueiredo D D,Cordeiro A M,et al.OsRMC,a negative regulator of salt stress response in rice,is regulated by two AP2/ERF transcription factors[J].Plant Mol Biol,2013,82(4/5):439-455.
[8]Tuteja N,Sahoo R K,Garg B,et al.OsSUV3 dual helicase functions in salinity stress tolerance by maintaining photosynthesis and antioxidant machinery in rice(Oryza sativa L.cv.IR64)[J].Plant J,2013,76(1):115-127.
[9]Chen L J,Wuriyanghan H,Zhang Y Q,et al.An S-domain receptor-like kinase,osSIK2,confers abiotic stress tolerance and delays dark-induced leaf senescence in rice[J].Plant Physiol,2013,163(4):1752-1765.
[10]Huda K M,Banu M S,Garg B,et al.OsACA6,a P-type IIB Ca(2+)ATPase promotes salinity and drought stress tolerance in tobacco by ROS scavenging and enhancing the expression of stress-responsive genes[J].Plant J,2013,76(6):997-1015.
[11]Zhang F T,Cui F L,Zhang L X,et al.Development and identification of a introgression line with strong drought resistance at seedling stage derived from Oryza sativa L.mating with Oryza rufipogon Griff[J].Euphytica,2014,200(1):1-7.
[12]王素平,郭世荣,胡晓辉,等.盐胁迫对黄瓜幼苗叶片光合色素含量的影响[J].江西农业大学学报,2006,28(1):32-38.
[13]汪月霞,孙国荣,王建波,等.NaCl胁迫下星星草幼苗MDA含量与膜透性及叶绿素荧光参数之间的关系[J].生态学报,2006(1):122-129.
[14]李大红.水稻RACK1基因(OsRACK1)在盐胁迫响应中的功能研究[D].扬州:扬州大学,2008:123.
[15]Troll W,Lindsley J.A photometric method for the determination of proline[J].J Biol Chem,1955,215(2):655-660.
[16]张国新,王秀萍,鲁雪林,等.隶属函数法鉴定水稻品种耐盐性[J].安徽农学通报,2011(1):36-37.
[17]陈洁.水稻幼苗耐盐性的定量鉴定及耐盐生理生化研究[D].儋州:华南热带农业大学,2003.
[18]金美芳,何菊芬.NaCl胁迫对水稻(Oryza sativa)种子萌发和幼苗生长的影响[J].福建师大福清分校学报,2010(2):6-10.
[19]宋旭丽,侯喜林,胡春梅,等.NaCl胁迫对超大甜椒种子萌发及幼苗生长的影响[J].西北植物学报,2011(3):569-575.
[20]Deinlein U,Stephan A B,Horie T,et al.Plant salt-tolerance mechanisms[J].Trends Plant Sci,2014,19(6):371-379.
[21]陈莎莎,兰海燕.植物对盐胁迫响应的信号转导途径[J].植物生理学报,2011(2):119-128.
[22]Munns R.,Tester M.Mechanisms of salinity tolerance[J].Annual Review of Plant Biology,2008,59:651-681.
[23]Hichri I,Muhovski Y,Zizkova E,et al.The solanum lycopersicum Zinc Finger2 cysteine-2/histidine-2 repressor-like transcription factor regulates development and tolerance to salinity in tomato and arabidopsis[J].Plant Physiol,2014,164(4):1967-1990.
[24]Xie Z,Duan L,Tian X,et al.Coronatine alleviates salinity stress in cotton by improving the antioxidative defense system and radical-scavenging activity[J].J Plant Physiol,2008,165(4):375-384.
[25]武宝玕,格林·托德.小麦幼苗中过氧化物歧化酶活性与幼苗脱水忍耐力相关性的研究[J].Journal of Integrative Plant Biology,1985(2):152-160.
