烟蚜茧蜂寄生对麦二叉蚜体内解毒酶和保护酶活性的影响
2015-05-10燕赛英杜孟芳尹新明安世恒王同朝贺德先
燕赛英,杜孟芳,尹新明,安世恒,王同朝,贺德先
(1.河南农业大学植物保护学院,小麦玉米作物学国家重点实验室,河南郑州450002;2.河南农业大学农学院,河南 郑州450002)
麦二叉蚜Schizaphis graminum(Rondani)是小麦上的重要害虫,分布于全国各产麦区。其在麦类叶片正、反两面或基部叶鞘内外吸食汁液,影响植株光合作用,致麦苗黄枯或伏地不能拔节,严重时麦株不能正常生长、抽穗,直接影响产量,此外可传带小麦黄矮病,传毒能力较强。5月上、中旬大量繁殖,为危害高峰期,可引起黄矮病流行。而目前传统的防蚜方法为喷洒化学农药,这对麦田天敌种群及生态环境将产生诸多负面影响。烟蚜茧蜂Aphidius gifuensis Ashmead是专性寄生蚜虫的一种内寄生蜂[1],可寄生桃蚜 Myzus persicae(Sulzer)、麦长管蚜Sitobion avenae(Fabricius)、萝卜蚜Lipaphis erysimi(Kaltenbach)、麦二叉蚜 Schizaphis graminum(Rondani)等,是蚜虫生物防治上的重要寄生蜂。由于寄生蜂必须依赖寄主昆虫才能生存,而寄主昆虫则不一定被寄生,因而二者的选择压力不同,所以形成防御和反防御的关系。寄生蜂的选择压力导致寄主昆虫演化出现新的防御性状,降低寄生蜂的寄生率和生存力;寄主昆虫新的防御性状又迫使寄生蜂演化产生新的反防御的性能,用于克服寄主的免疫反应,这种防御与反防御的选择压力造成它们之间的协同进化。寄主应对寄生物的免疫机制主要有细胞免疫反应(吞噬、成瘤、包囊作用等)和体液免疫。故寄生蜂幼虫想要正常发育,必须改变寄主代谢,克服寄主免疫系统的防御反应[2]。烟蚜茧蜂雌蜂产卵时同时注入分泌物保护蜂卵,这些因子在克服寄主免疫、延长寄主若虫期,调控内分泌系统,使蜂卵及幼虫免于寄主的细胞免疫反应(吞噬、成瘤、包囊作用等)和体液免疫中发挥重要作用,而解毒酶和保护酶作为抵御外界寄生因子的直接作用者,通过其活性变化可直观得到寄主与寄生蜂之间相互作用关系。解毒酶主要存在于昆虫中肠、线粒体、脂肪体以及高尔基体中,能够代谢大量的内源或外源化合物,使昆虫的生理代谢活动维持在正常水平。乙酰胆碱酯酶(AChE)是生物神经传导中一个关键性酶,也是昆虫体内唯一一种胆碱酯酶,其主要作用为催化神经递质乙酰胆碱的分解,从而终止神经冲动的传导。酸性磷酸酯酶、碱性磷酸酯酶为昆虫体内重要的解毒代谢酶,通过水解、氧化、还原、轭合等参与各种外源物的代谢。需氧生物都有自由基的产生与清除这2个机制,而自由基具有高度化学活性,若两者之间平衡失调,便会损伤生物机体引起病变。昆虫体内存在的超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)等保护酶系统可保护昆虫机体免受氧自由基的伤害。目前,国内外对烟蚜茧蜂的研究仅局限于其生物学特性、生态学以及繁殖和释放[3-9]等方面,未涉及寄生后寄主生理生化的变化。本研究以烟蚜茧蜂为试虫,通过研究寄生体系中麦二叉蚜乙酰胆碱酯酶Acetylcholinesterase(AChE)、酸性磷酸酯酶Acid phosphastase(ACP)、过氧化氢酶Catalase(CAT)、过氧化物酶Pemxidase(POD)、总超氧化物歧化酶 Total superoxide dismutase(T-SOD)活性的变化规律,探讨寄生蜂寄生后对寄主体内生理生化的调控,为研究两两之间关系提供理论基础。
1 材料与方法
1.1 供试昆虫
麦二叉蚜采自河南农业大学科教园区小麦田,采回后放入种植有新鲜小麦的花盆中分开饲养多代,直到得到未被寄生及外来生物感染的室内纯化种群。烟蚜茧蜂同样采自河南农业大学科教园区小麦田,采回后用麦二叉蚜饲养繁殖多代。室内温度为(27±2)℃,相对湿度为(70±10)%,光照条件 L∶D=14 h∶10 h。
1.2 方法
1.2.1 烟蚜茧蜂对麦二叉蚜的寄生 正常的麦二叉蚜为对照组;被烟蚜茧蜂寄生的麦二叉蚜为处理组。所选取麦二叉蚜均为2龄且个体大小一致的若蚜。取780头麦二叉蚜若蚜,以每管20头的量放入装有新鲜麦苗叶子的2 mL离心管中,同时每管放入7头同批次24 h内羽化并已交配的雌性烟蚜茧蜂,寄生30 min后取出雌蜂,将若蚜放入种植有新鲜小麦的纸杯中隔离饲养。同时,另取780头麦二叉蚜若蚜进行相同人为处理,但不放入烟蚜茧蜂,以此作为对照组。
1.2.2 粗酶液的制备 将对照组的15头蚜虫放入1.5 mL离心管中,加1 mL预冷的磷酸缓冲液(含0.1%的Triton X-100),冰浴下充分研磨,在4℃下12 000 r· min-1离心15 min,上清液即为粗酶液,提取后-20℃保存备用,分析前解冻。以同样方法同一时间对处理组的蚜虫进行制样,重复3次。以此获得寄生 0、1、2、3、4、5、6、8、10、12、24、48、72 h后的样品及对照组粗酶液。
1.2.3 解毒酶活性测定 采用紫外可见光分光光度计对酶的活性进行测定,仪器采用上海光谱仪器有限公756型紫外可见分光光度计。
乙酰胆碱酯酶(AChE)、酸性磷酸酯酶(ACP)、碱性磷酸酯酶(AKP)活性测定方法参照南京建成生物工程研究所各酶活测定试剂盒。
1.2.4 保护酶活性测定 测定方法同解毒酶活性的测定方法。过氧化氢酶(CAT)、过氧化物酶(POD)、总超氧化物歧化酶(T-SOD)的活性测定方法参照南京建成生物工程研究所各酶活测定试剂盒。
1.2.5 数据处理 数据采用 SPSS18.0软件和Excel 2003进行分析。数据均为平均值±标准误(Means±SE),处理组与对照组之间的差异采用Tukey氏检验进行比较,以0.05作为显著性水平(P<0.05),以 0.01作为极显著性水平(P<0.01)。
2 结果与分析
2.1 烟蚜茧蜂寄生对2龄麦二叉蚜体内乙酰胆碱酯酶(AChE)活性的影响
烟蚜茧蜂寄生麦二叉蚜后0~72 h寄主体内AChE活性测定结果见图1。处理组与对照组AChE活性总体变化趋势基本一致。处理组在寄生后2、12、24、48、72 h 时 AChE 活性明显低于对照组,且在12、72 h时极显著低于对照。处理组在寄生后4、10 h时AChE活性略高于对照组。通过测定结果,说明烟蚜茧蜂的寄生引起了麦二叉蚜体内AChE活性的变化,可能由于烟蚜茧蜂的产卵对寄主的解毒能力产生了干扰。
2.2 烟蚜茧蜂寄生对2龄麦二叉蚜体内酸性磷酸酯酶(ACP)活性的影响
烟蚜茧蜂寄生麦二叉蚜后0~72 h寄主体内ACP活性测定结果见图2。处理组与对照组ACP活性总体变化趋势稍有不同。处理组在寄生后3、5 h时ACP活性明显低于对照组,但在24 h时ACP活性极显著高于对照组。通过测定结果,说明烟蚜茧蜂的寄生对麦二叉蚜体内ACP活性产生影响,在3、5 h时对寄主体内ACP活性抑制效果较明显,在24 h对寄主体内ACP活性激活效果极其明显,24 h后又呈下降趋势。

图1 2龄麦二叉蚜被烟蚜茧蜂寄生后体内AChE活性随时间的变化Fig.1 The AChE activities of the second instar nymph Schizaphis graminum in relation to the different time after having been parasitized by Aphidius gifuensis

图2 2龄麦二叉蚜被烟蚜茧蜂寄生后体内ACP活性随时间的变化Fig.2 The ACP activities of the second instar nymph Schizaphis graminum in relation to the different time after having been parasitized by Aphidius gifuensis
2.3 烟蚜茧蜂寄生对2龄麦二叉蚜体内碱性磷酸酯酶(AKP)活性的影响
烟蚜茧蜂寄生麦二叉蚜后0~72 h寄主体内AKP活性测定结果见图3。处理组与对照组AKP活性总体变化趋势稍有不同。处理组在寄生后2、3、5、24、48 h 时 AKP 活性明显低于对照组,仅在10、12 h时AKP活性略高于对照。通过测定结果,说明烟蚜茧蜂的寄生干扰了麦二叉蚜体内AKP解毒活性,在0~8 h、24~72 h期间AKP活性受到抑制,低于正常水平。
2.4 烟蚜茧蜂寄生对2龄麦二叉蚜体内过氧化氢酶(CAT)活性的影响
烟蚜茧蜂寄生麦二叉蚜后0~72 h寄主体内CAT活性测定结果见图4。处理组与对照组CAT活性总体变化趋势基本一致。处理组在寄生后3、6 h时CAT活性略低于对照组,但在24 h时显著高于对照。通过测定结果,说明烟蚜茧蜂的寄生在3、6 h时干扰了寄主体内CAT的保护能力,这有利于寄生蜂卵的发育,烟蚜茧蜂卵在室内温度(27±2)℃,相对湿度(70±10)%,光照条件L∶D=14 h∶10 h的环境内经过20~40 h孵化成幼虫,故推测24 h时因为幼虫的孵出,使寄主体内微环境遭到破坏,代谢加强,导致H2O2过多积累,诱导CAT活性增大。
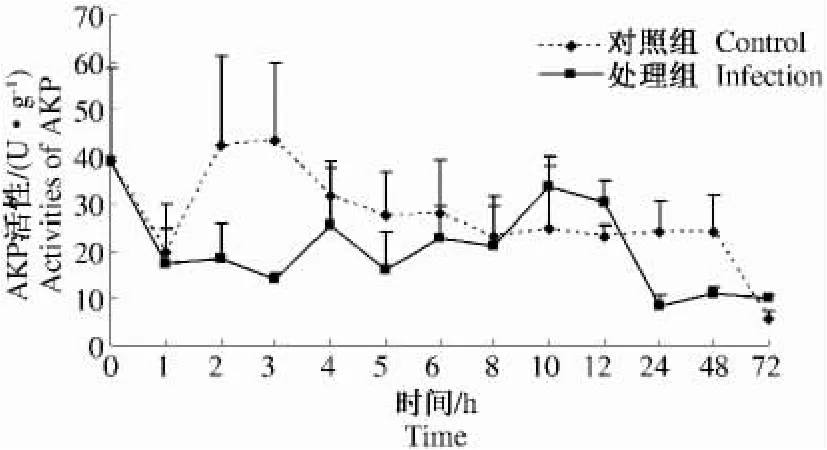
图3 2龄麦二叉蚜被烟蚜茧蜂寄生后体内AKP活性随时间的变化Fig.3 The AKP activities of the second instar nymph Schizaphis graminum in relation to the different time after having been parasitized by Aphidius gifuensis

图4 2龄麦二叉蚜被烟蚜茧蜂寄生后体内CAT活性随时间的变化Fig.4 The CAT activities of the second instar nymph Schizaphis graminum in relation to the different time after have been parasitized by Aphidius gifuensis
2.5 烟蚜茧蜂寄生对2龄麦二叉蚜体内过氧化物酶(POD)活性的影响
烟蚜茧蜂寄生麦二叉蚜后0~72 h寄主体内POD活性测定结果见图5。处理组与对照组POD活性总体变化趋势基本一致。处理组在寄生后5、6、8 h时POD活性低于对照组,且在6 h时活性显著低于对照组。在1、12、24 h时POD活性高于对照,且在24 h时活性极显著高于对照。通过测定结果,说明烟蚜茧蜂的寄生对麦二叉蚜体内POD活性产生了一定影响,1 h时POD活性增强,利于寄主保护自身机体,12、24 h时POD活性明显增加,有利于寄主昆虫的正常发育,也有利于寄生蜂获得更多营养。
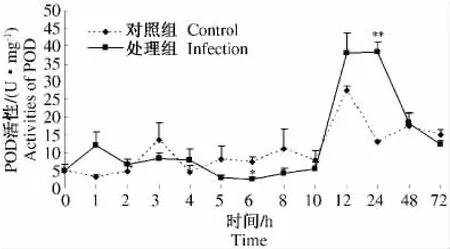
图5 2龄麦二叉蚜被烟蚜茧蜂寄生后体内POD活性随时间的变化Fig.5 The POD activities of the second instar nymph Schizaphis graminum in relation to the different time after having been parasitized by Aphidius gifuensis
2.6 烟蚜茧蜂寄生对2龄麦二叉蚜体内总超氧化物歧化酶(T-SOD)活性的影响
烟蚜茧蜂寄生麦二叉蚜后0~72 h寄主体内T-SOD活性测定结果见图6。处理组与对照组T-SOD活性总体变化趋势基本一致。处理组在寄生后1、4、24 h时T-SOD活性稍高于对照组,在5 h时略低于对照。测定结果说明,烟蚜茧蜂的寄生对麦二叉蚜体内T-SOD活性有一定的影响,在1、4 h时寄主代谢系统担负着寄主和寄生蜂卵的共同发育,氧气消耗量增加,从而诱导T-SOD活性稍有升高,推测24 h时幼虫孵出,使 T-SOD活性略有升高。

图6 2龄麦二叉蚜被烟蚜茧蜂寄生后体内T-SOD活性随时间的变化Fig.6 The T-SOD activities of the second instar nymph Schizaphis graminum in relation to the different time after having been parasitized by Aphidius gifuensis
3 结论与讨论
寄生蜂影响寄主的寄生因子主要有类病毒颗粒(VLP)或类病毒样纤丝(VLF)、毒液、萼液(主有含PDV、卵巢蛋白)等。寄生蜂产生的毒蛋白如毒液、萼液和畸形细胞能使半翅目蚜虫免疫抑制、发育受阻[10]。俞瑞鲜[11]提出萼液通过毒液而实现对寄主的作用,而畸形细胞和幼蜂的存在又能明显改变寄主的行为。可得出幼蜂的存在保证了畸形细胞正常发挥作用。曹婷婷[12]用0.05VRE浓度的毒液或0.05 FE·μL-1浓度的毒液和萼液混合物处理离体条件下小菜蛾血细胞,使其失去了包囊作用。说明寄生蜂产卵初期毒液和萼液共同作用抑制了寄主的免疫系统正常运作,使蜂卵进行发育,随着蜂卵的生长,刺激产生的畸形细胞开始发挥反包囊作用,进一步抑制或破坏寄主的免疫系统。李艳红[13]得出毒液和萼液能显著延长若虫的发育;正常寄生后,寄生蜂毒蛋白使寄主始终处于抑制状态,并对其免疫系统产生抑制作用,毒蛋白量的增加加大了对寄主的抑制程度。
麦二叉蚜免疫系统中的AChE是一种在神经传递过程中起重要作用的酶,此酶一旦被抑制达一定程度时,会使动物过度兴奋而死亡[14]。本研究发现,麦二叉蚜被寄生后,寄生蜂通过干扰胆碱能突触而引起神经系统的障碍,从而抑制AChE活性,使昆虫生理生化过程失调和破坏。与小菜蛾幼虫被半闭弯尾姬蜂寄生后,AChE活性降低,结果相似[15]。有研究表明,从低等生物大肠杆菌到高等动植物组织以及人类肝脏等都发现有ACP的存在[16,17]。在缺乏抗原抗体特性的昆虫体内,ACP的作用很重要,是溶酶体的标志酶。在酸性条件下,ACP能通过水解作用将表面带有磷酸酯的异物破坏,达到预防感染的目的。而碱性磷酸酶(AKP)不仅参与机体内某些营养物质的代谢过程,而且还是生物体内重要的解毒体系[18]。本研究中ACP活性处理组在寄生后3、5 h时ACP活性明显低于对照组,说明寄生蜂的寄生影响寄主体内ACP的释放,而24 h时剧增,显著高于对照组,推测此时寄主吞噬、降解了一部分外源物,使寄生蜂的寄生因子没有初期强烈,故在24 h时大量释放ACP为自身解除毒害。AKP活性仅在10、12 h时略高于对照,在 2、3、5、24、48 h 时均受抑制,说明寄生蜂寄生后蜂卵及卵液作为异物引起麦二叉蚜体内解毒酶系强烈的反应,而24 h后寄生蜂幼虫开始发育,与畸形细胞共同影响寄主体内小环境,致使寄主解毒效果下降,酶活性降低。
昆虫保护酶系活性与杀虫剂、昆虫病毒、微孢子虫等外界刺激物的强度及昆虫耐药性和抗逆性等相关[19-22]。昆虫体内存在着超氧化物歧化酶(SOD)、过氧化氢酶(CAT)和过氧化物酶(POD)等保护酶系统,在昆虫各种生理作用或生化反应过程中起着清除氧自由基、保护昆虫机体免受损伤的重要作用[23]。本研究中,CAT作为机体清除氧自由基的重要组成部分,其活性在24 h显著高于对照,这与前人的研究结果相似[24-25]。POD 活性处理组在寄生后5、6、8 h时POD活性低于对照组,且在6 h时活性显著低于对照组,说明寄生蜂的寄生导致麦二叉蚜体内ATP合成受阻,致使能量供应不足,抑制了寄主体内POD的活性,而在24 h时活性极显著高于对照,说明24 h时左右试虫体内供能逐渐恢复正常,寄主免疫系统产生了细胞免疫反应来保护寄主自身机体;SOD能催化超氧化物阴离子自由基发生歧化反应生成H2O2,从而清除阴离子自由基,麦二叉蚜被寄生后体内H2O2量的增加未达到抑制T-SOD活性的程度,故处理组在寄生后1、4、24 h时T-SOD活性反而稍高于对照组,仅5 h时略低于对照,处理组与对照组T-SOD活性总体变化趋势基本一致。
本研究旨在从麦二叉蚜被烟蚜茧蜂寄生后体内6种酶活性的变化这个方面为出发点,研究了6种酶随时间变化的总体趋势。其中,CAT活性变化规律与前人的研究结果基本一致,但也存在差异,可能由于不同供试昆虫体内酶种类、浓度及其组成不同,或昆虫个体生理机能的变化、蜕皮等原因造成的。昆虫的免疫防御是多种酶相互作用的结果,不同时间段起主导作用的酶不同,各种酶相互配合、协同作用,才能维持昆虫机体的正常生理活动。而寄生蜂必须依赖寄主昆虫才能生存,寄生成功后既不能严重破坏寄主的免疫系统,又要保证蜂卵在寄主体内不被损害,正常发育,故寄生蜂寄生成功后相关寄生因子相互作用阶段性影响寄主体内各种酶的活性,保证寄主在蜂卵发育时期继续生长,而在蜂幼虫化蛹时能及时变成僵蚜确保幼蜂正常羽化。
[1] BARLOW C A.The influence of temperature on the growth of experimental populations of Myzus persicae(Sulzer)and Macrosiphum euphorbiae(Thomas)(Aphididae) [J].Canadian Journal of Zoology,1962,40:145-156.
[2] 尹丽红,王琛柱,钦俊德.多分DNA病毒及其在寄生蜂与寄主关系中的作用[J].昆虫学报,2001,44(1):109-118.
[3] 毕章宝,季正端.烟蚜茧蜂生物学研究Ⅰ.发育过程和幼期形态[J].河北农业大学学报,1993,16(2):2-8.
[4] 毕章宝,季正端.烟蚜茧蜂生物学研究Ⅱ.成虫生物学及越冬[J].河北农业大学学报,1994,17(2):38-44.
[5] 毕章宝,季正端.烟蚜茧蜂生物学研究Ⅳ.繁殖力、内禀增长力、功能反应及对桃蚜的抑制作用[J].河北农业大学学报,1996,19(3):1-5.
[6] 汤玉清,陈珠梅.烟蚜茧蜂生物学特性的初步研究[J].福建农学院学报,1984,13(2):119-124.
[7] 路 虹,石宝才,张芝利.烟蚜茧蜂生殖特性研究[J].华北农学报,1993,8(1):76-79.
[8] 王文夕,李巧丝.寄主密度对烟蚜茧蜂生殖特性的影响[J].华北农学报,1996,11(4):52-57.
[9] 忻亦芬.烟蚜茧蜂繁殖利用研究[J].生物防治通报,1986,2(3):108-111.
[10] HOTTA M,OKUDA T,TANAKA T.Cotesia kariyai teratocytes:growth and development[J].Journal of Insect Physiology,2001,47:31 -41.
[11]俞瑞鲜.菜蛾盘绒茧蜂毒液对寄主小菜蛾生理调控机制的研究[D].杭州:浙江大学,2006.
[12]曹婷婷.菜蛾盘绒茧蜂毒液和萼液对寄主小菜蛾幼虫的免疫调控研究[D].杭州:浙江大学,2008.
[13]李艳红.烟蚜茧蜂毒蛋白对寄主生长发育的影响[J].吉林农业科学,2014,39(4):43-46.
[14]唐培安,邓永学,王进军.甲酸乙酯对米象乙酰胆碱酯酶和羧酸酯酶的影响[J].植物保护,2007,33(1):44-47.
[15]黄 芳,章金明,郦卫弟,等.半闭弯尾姬蜂寄生对小菜蛾幼虫解毒酶系的影响[J].浙江农业科学,2012(5):700-703.
[16] SAINI M S,ETTEN R L.An essential carboxylic acid group in human prostate acid phosphatase[J].Biochemicaet Biophysica Acta,1979,568(2):370-376.
[17] TOHE A,UEDA Y,KAKIMOTO S I,et al.Isolation and characterization of acid phosphatase mutants in Saccharomyces cerevisiae[J].Biochem J,1973,113(2):727-738.
[18]何海琪,孙 凤.中国对虾酸性和碱性磷酸酶的特性研究[J].海洋与湖沼,1992,23(5):555-560.
[19]李周直,沈惠娟,蒋巧根,等.几种昆虫体内保护酶系统活力的研究[J].昆虫学报,1994,37(4):399-403.
[20]蒋志胜,尚稚珍,万树青,等.光活化杀虫剂a-三噻吩的电子自旋共振分析及其对库蚊保护酶系统活性的影响[J].昆虫学报,2003,46(1):22-26.
[21] WANG Y,OBERLEY L W,MURHAMMER D W.Evidence of oxidative stress following the viral infection of two Lepidopteran insect cell lines[J].Free Radical Biology and Medicine,2001,31(11):1448-1455.
[22] LOZINSKAYA Y L,SLEPNEVA I A,KHRAMTSOV V V,et al.Changes of the antioxidant status and system of generation of free radicals in hemolymph of Galleria mellonella larvae at microsporidiosis[J].Journal of Evolutionary Biochemistry and Physiology,2004,40(2):119-125.
[23]王满囷,李周直.滞育期间鞭角华扁叶蜂保护酶系统活力[J].林业科学,2002,38(4):100-104.
[24]冯从经,戴华国,符文俊.腰带长体茧蜂寄生后亚洲玉米螟体内抗氧化酶活性及组织特异性[J].南京农业大学学报,2002,25(3):31-35.
[25]梁光红,陈友荣,黄居昌.寄生对橘小实蝇幼虫体内过氧化物酶活性的影响[J].中南林业科技大学学报,2007,27(2):48-51.
