腾冲云峰山种子植物区系
2015-04-29杨晓君等
杨晓君等


摘要通过对腾冲云峰山35 km2范围内的全部种子植物进行标本的采集和考察,对云峰山植物区系进行分析研究。结果表明:云峰山种子植物多样性十分丰富。该地区有种子植物92科247属431种,其中裸子植物4科5属7种,被子植物88科242属424种。分布类型多样,形成以热带亚热带成份为主的植物区系。
关键词腾冲;云峰山;种子植物;植物区系
中图分类号S718.54文献标识码
A文章编号0517-6611(2015)29-006-03
作者简介杨晓君(1990- ),女,云南保山人,硕士研究生,研究方向:高等植物资源。*通讯作者,教授,从事高等植物资源方面的研究。
收稿日期20150824
腾冲云峰山是云南著名的道教名山,山顶有始建于明朝崇祯七年(1632年)的道观,属国家AAA级著名风景名胜区、国家地质公园。明代旅行家徐霞客赞美它为“似太华之苍龙脊”。由于道观一直将云峰山上的植物作为风水林保护,近400年来植物未被破坏过,生态系统完善,植物群落结构完整、自然,种子植物种类多样性十分丰富。
高黎贡山地区是指怒江(Salween River)和伊洛瓦底江(Irrawarddy River)之间的分水山脉和山脉两侧的地域,位于24°40′~28°30′ N。由于特殊的地质历史和独特的生态自然环境差异,该区由北向南常被分为北、中、南三段[1-3]。
云峰山是高黎贡山地区南段属于伊洛瓦底江两大支流(龙川江和槟榔江)之间的高黎贡山南支系狼牙山脉中的一个形态特别的山峰。它地处98°20′~98°28′E、25°19′~25°25′N的云南省腾冲县瑞滇盆地西南边缘。该区域属于横断山南延的高黎贡山山脉支系狼牙山区的高中山峡谷区。云峰山在一个峡谷(瑞滇盆地)的西南边缘拔地而起。从盆地海拔1 750 m开始,在2 km的距离内海拔上升700 m,山峰顶端海拔2 450 m,主峰挺拔俊秀,山顶常年云雾环绕,东西两边有陪山,陪山海拔低于主峰200 m左右,与主峰形成如汉字“山”形,成为瑞滇盆地四周最显眼的山峰。云峰山形成腾冲境内槟榔江和龙川江水系的分水岭,北面、东面、南面水流入龙川江,西面水流入槟榔江。龙川江和槟榔江都是缅甸最大河流伊洛瓦底江在中国境内的两大支流。在云峰山山谷中有瑞滇热泉的热泉显示区,温泉资源丰富,所以山谷中常年水雾缭绕。
云峰山属亚热带季风气候区,常年受印度洋西南季风控制,气候湿和,雨量充沛,年降雨量平均1 715 mm,干季雨季分明。年温差小,年平均气温为14.6 ℃,最低气温-2.2 ℃,最高气温29.5 ℃,无雨期230 d,日照充足,年均日照2 000 h,太阳总辐射量130.8 J/cm2。
该地区成土母质为第四系洪积母质,土壤主要是黄棕壤和火山灰堆积土。高黎贡山地区植物区系以及北、中、南各段大范围的植物区系均已做过大量的研究[4-9],但是对狼牙山山脉的植物区系研究则未见报导。笔者研究了云峰山近400年保护的风水林的生态系统结构特点。
1材料与方法
对云峰山35 km2范围内的全部种子植物进行了4年多批次的标本采集和考察,共采集标本1.2万号,现部分仅存云南农业大学,部分存香格里拉县高山植物园。在鉴定所采集的标本的基础上,形成腾冲云峰山地区植物名录,再应用植物区分学的研究方法对云峰山的植物区系进行分析。
2结果与分析
2.1区系的组成
由表1可知,该地区有种子植物92科、247属、431种(含亚种和变种),分别占高黎贡山南段种子植物科、属、种的47.92%、28.13%和15.35%,分别占云南省种子植物科、属、种的31.83%、11.56%和3.08%,分别占全国种子植物科、属、种的31.62%、7.62%和1.77%,其中裸子植物4科、5属、7种,被子植物88科、242属、424种。
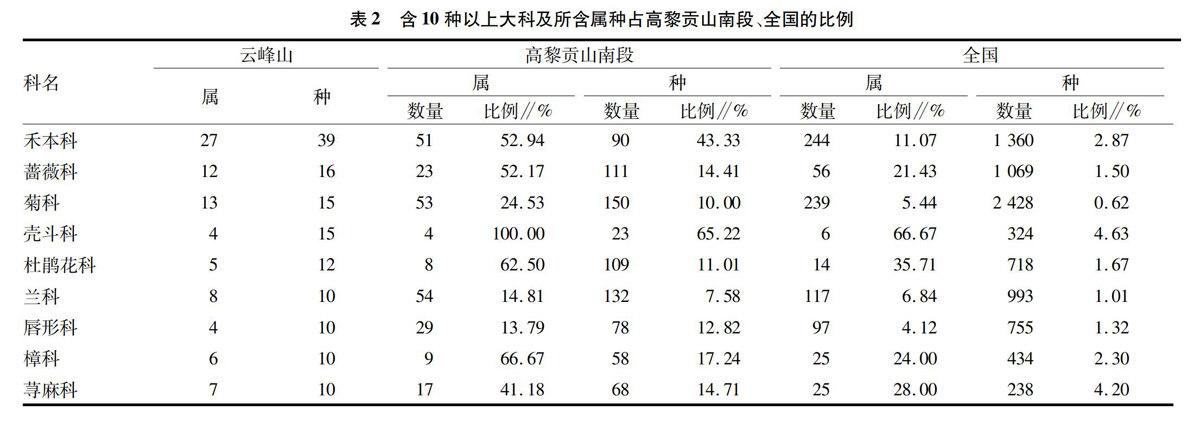
据调查,发现壳斗科、山茶科、樟科、金缕梅科、松科、杜鹃
花科、蔷薇科是构成群落结构中乔木层和灌木层的主要树种。区域内由石栎、青冈、银木荷、马蹄荷、厚皮香、紫点杜鹃等种类为乔木层灌木层的中山温性常绿阔叶林是最主要的植被类型。在较老的青冈、石栎树干上或树干顶端枯死部分常附生着兰科的伏生石豆兰、疣鞘贝母兰、盆距兰、细茎石斛等植物。
根据吴征镒等和李锡文[11]对科的分布区类型系统的划分,云峰山地区种子植物92个科可划分为8个类型和4个变型。在这么小的范围内有如此丰富的科分布类型,说明该地区科级水平上的地理成分比较复杂,联系广泛。
由表3可知,该区中有世界分布29科,含种类最多的9个科的4个(禾本科(Gramineae)、蔷薇科(Rosaceae)、菊科(Compositea)、唇形科(Labiatea))属于这一分布类型,占该地区92科的31.52%,近1/3;熱带分布类型(含变型)有42科,占所有92个科的45.65%,接近一半比例,占除世界分布科以外的科类的66.67%,表明该区系内热带成分占主导地位;温带分布类型(含变型)有20个科,占所有92科的21.73%,东亚分布类型有1个科;温带分布类型的科数不到热带分布类型科数的一半。由此可知,该区域是以热带成分科为主的植物区系类型。
2.3属级统计分析
按吴征镒、应俊生的中国种子植物属的分布区类型划分方案[12],该区域247个属可以被划分为13类型和11个变型,缺少温带亚洲分布类型。可以看出,该区域植物成分比较复杂。这与该区是地理上古南大陆和古北大陆相互结合的交叉地带相关。许多源于印度—马来植物区系成分和东亚植物区系成分(含北温带)在这里融合发展,共同演变成今天植物区系外貌,成为世界重要的生物多样性中心。
2.3.1世界分布。
由表4可知,该区系中有世界分布属33属,占整个区系总属数的13.36%。这33个属的植物绝大部分是草本植物。它是该区系中草本植物的主要构成成分,主要有蔊菜属(Rorippa)、毛莨属(Ranunculus)、悬钩子属(Rubus)、车前(Plantago)、灯芯草属(Juncus)、苔草属(Carex)、剪股颖属(Agrostis)、马唐属(Digitaria)等。
2.3.2热带分布。
该区系中的热带分布类型(2~7)6个类型都齐全,并且含有6个变型,共计128属,占除世界分布属以外属数的59.81%,含有植物168种(含亚种和变种),占除世界分布属种类植物以外种类数目的43.64%。在该区系科的分布类型和属的分布类型中,热带分布类型占除世界分布的科和属以外的科、属数比例分别为66.67%和59.81%,可以看出该区系植物是以热带、亚热带植物为主。
泛热带分布61属(含变型),占热带分布的47.66%,占该区系的24.70%,是热带分布的最大、最主要类群,也是该植物区系的最重要组成成分。该类群主要含有胡椒属(Peperomia)、冷水花属(Pilea)、风仙花属(Impatiems)、冬青属(Ilex)、鹅掌柴属(Schefflera)、卫予属(Euonymuus)、紫金牛属(Ardisia)、合欢属(Acacia)、菝契属(Smilax)。
热带亚洲和热带美洲间断分布有7属,占热带分布的5.47%,如柃属(Eurya)、木姜子属(Litsea)、水车哥属(Saurauiy)、白珠树属(Gaultheria)等。这些属是该区系内植物群落结构中亚乔木灌木层的重要构成树种。
旧世界热带分布15属(含变型),占热带分布的11.72%,如楼梯草属(Elatostema)、乌蔹莓属(Cayratia)、酸筒子属(Embelia)、八角枫属(Alangium)、娃儿藤属(Tylophora)、天门冬属(Asparagus)。
热带亚洲至热带大洋洲分布7属(含变型),占热带分布的5.47%,如梓属(Cinnamomum)、崖藤属(Tetrastigma)、兰属(Cymbidium)、野牡丹属(Melastoma)等。
热带亚洲至热带非洲分布9属,占热带分布的7.03%,如水府属(Debregeasia)、铁仔属(Myrsine)、莠竹属(Microstegium)、芒(Miscanthus)、菅属(Themeda)。
热带亚洲分布29属(含变型),占热带分布的22.66%,是热带分布的第二大类型,如含笑属(Michelia)、山胡椒属(Lindera)、润楠属(Machilus)、新梓属(Neocinnamomum)、山茶属(Camellia)、木荷属(Schima)、马蹄荷属(Exbucklandia)、假木荷属(Craibiodendron)、崖角藤属(Rhaphidophora)、绞股蓝属(Gynostemme)、树萝卜属(Agapetes)、芒毛苣苔属(Aeschynanthus)、石斛属(Dendrobium)、棕叶芦属(Thysanolsena)等。该类型是该区系植物群落结构乔木、灌木、草本的重要植物成分,是该区系内热带亚热带代表植物,为主要建群树种和特色树种,在群落中种群密度大。
2.3.3温带分布。温带分布5个主要类型和5个变型,共计83属,占除世界分布属以外的38.86%,含植物216种(含变种),种数占除世界分布以外的38.79%。
该区系中温带分布科和属的数量都远远低于热带分布的数量,但是温带分布属所含的植物种类数量多于热带分布属所含的种类数目。该区域是热带植物与温带植物的交融地带。这与高黎贡山南段植物区系主要成分性质是一致的。
北温带分布42属,占温带分布的50.60%,占区系内的17.00%,不仅是温带分布中的最大类群,而且是该区系内的第二大类群,42属中有植物58种。北温带分布类型中的植物是构成该区系植物群落结构之一的类型。许多种类是该区系中的主要树种,如松属(Pinus)、小檗属(Berberis)、蔷薇属(Rosa)、绣线菊属(Spiraea)、樱属(Cerisus)、栎属(Quercus)、杜鹃花属(Rhododendron)、 荚蒾属( Viburnum)、艾蒿属(Artemisia)、婆婆纳属(Veronica)、稗属(Echinochloa)、画眉草属(Eragrostis)。杜鹃花属含有7种,栎属含有4种,是种类较多的大属。在北温带分布类型中,有一个变型,8-4北温带和南温带间断分布,其越桔属(Vaccinium)、婆婆纳、杨梅(Myrica)、荨麻(Vrtica)中的植物种类也是群落结构中个体密度较大的种类。
东亚和北美间断分布14属,占温带分布的16.87%,如石楠属(Photinia)、石栎属(Lithocarpus)、 勾儿茶属(Berchemia)、爬山虎属(Parthenocissus)、米饭花属(Lyonia)。
安徽农业科学2015年
旧世界温带其变型(地中海区、西亚和东亚间断分布)含有5个属。地中海区、西亚至中亚分布变型(地中海区至温帶、热带亚洲、大洋洲和南美洲间断分布)含有木樨榄属(Olea)、云南木樨榄(O.yuennanensis)。
中亚分布1属,东亚分布(含变型)20属,占温带分布的24.10%,可以看出该区系地域已是东亚分布的边缘,但仍分布着大量的东亚分布类型的植物种类。其中,东亚(东喜马拉雅—日本)分布10属,如沿阶草属(Ophiopogon)、棕榈属(Trachycarpus)、兔儿风属(Ainsliaea)、白及属(Bletilla)、虎刺属(Damnacanthus)、沿阶草属在该区系内有4种之多。中国—喜马拉雅变型分布9属、如八月瓜属(Holboellia)、鬼吹箫属(Leycesteria)、珊珊苣苔属 (Corallodiscus)、鞭打绣球(Hemiphragma)、吊石苣苔(Lysiontus)、筒冠花(Siphocranion)。中国—日本变型分布有半夏属(Pinellia)半夏(P.ternata)。
中国特有分布3属。杉属(Cunninghamia)的杉木(C.lanceolata)在腾冲被广泛种植,在该区系中也许是遗生树种,其长势很好。在腾冲,它是最主要的大面积造林树种,也是当地农民房前屋后的绿化树种。另外,有喜树属(Camptotheca)的喜树(C.acuminata)、牛筋条属(Dichotomanthus)的牛筋条(D.tristaniacarpa)。
2.4与邻近植物区系的比较
云峰山位于高黎贡山植物区系范围西南部边缘区域,处于古南大陆的边缘,其植物区系与高黎贡山南段有很大的相似性,同时其区别也非常明显,与高黎贡山北段的差别较显著。
在云峰山区系中最大10科中,世界分布4科,热带分布5科,温带分布仅1科,热带分布与温带分布之比为5∶1。云峰山区系中最大10个科与高黎贡山南段[13]的最大科有9科是一致的,不同科是云峰山区系的壳斗科,而高黎贡山南段区系是蝶形花科。在高黎贡山南段最大10科中,世界分布6科,热带分布3科,温带分布1科,热带分布与温带分布之比为3∶1;在高黎贡山北段区系最大10科中,世界分布7科,热带2科,温带1科,热带分布与温带分布之比为2∶1。在92个科中,云峰山区系有42个热带分布科,20个温带分布科,两者比为2∶1。高黎贡山南段区系有192科,其中81个热带分布科,39个温带分布科,两者比为1.33∶1.00。可见,云峰山区系的热性成分高于整个高黎贡山南北段区系。这可能是其地理上更接近于印度—马来区系的原因。
3结论与讨论
(1)云峰山地区种子植物种类丰富,在约35 km2的范围内有种子植物92科247属431种(包括亚种和变种),分别占高黎贡山南段的科、属、种类比例47.92%、28.13%和15.35%,分别占云南省种子植物的科、属、种类的比例3183%、11.56%和3.08%,分别占全国种子植物的科、属、种类的比例31.62%、7.62%和1.77%。
(2)云峰山地区种子植物区系组成成份复杂,生物多样性丰富。中国种子植物科的14个分布区类型有8个,29个变型中有4个。中国种子植物属的15个分布区类型中有13个,31个变型中11个。其中,世界分布属有33属,占总数的13.36%;热带分布128属,占总数的51.82%,占除世界分布以外属数的59.81%;温带分布83属,占总数的33.60%,占除世界分布属以外属数的38.79%;中国特有属3属。
(3)云峰山地区植物区系基本上是热带、亚热带区系性质。从科总数看,热带亚热带分布类型共42科,占除世界分布29科以外所有科数的66.67%,是温带分布类型的2倍多,热带成分明显。从最大的9个科看,世界分布4个科,热带亚热带分布4个科,而温带分布只有1个科。从属数看,热带亚热带分布128属,占除世界分布属以外属数的5981%,其中泛热带分布61属,热带亚洲分布(印度—马来)29属,而温带分布只有83属。
(4)云峰山地区属于高黎贡山南段范围,两者的种子植物区系成分基本一致,而与高黎贡山北段植物区系差异明显。这一区域属于东亚植物区系的中国—喜马拉雅区的南段,与印度—马来植物区系交汇,印度—马来区系占明显优势,形成以热带亚热带成分为主的植物区系特征。
参考文献
[1]
李嵘,刀志灵, 纪运恒,等.高黎贡山北段种子植物区系研究[J].云南植物研究,2007,29(6):601-615.
[2] 李恒,郭辉军,刀志灵,等.高黎贡山种子植物特有种研究[M]//李恒,郭辉军,刀志灵.高黎贡山植物.北京:科学出版社, 2000:250-324.
[3] 刘伦辉,刀志灵,郭辉军, 等.高黎貢山的植被[M]//李恒,郭辉军,刀志灵.高黎贡山植物.北京:科学出版社, 2000:6-48.
[4] 吴征镒, 周浙昆, 李德铢, 等.世界种子植物科的分布区类型系统[J].云南植物研究,2003,25(3):245-257.
[5] 吴征镒,朱彦丞.云南植被[M].北京:科学出版社,1987:81-793.
[6] 李嵘,李恒,刀志灵,等.中国植物区系新资料[J].武汉植物学研究,2003,21(2):153-154.
[7]李锡文,李捷.横断山脉地区种子植物区系的初步研究[J].云南植物研究,1993,15(3):217-231.
[8] 汪建云, 母其爱, 蔺如涛.高黎贡山自然保护区南段生物走廊带维管植物名录(1)[J].云南师范大学学报,2008,28(3):57-63.
[9] 汪建云, 母其爱, 蔺如涛.高黎贡山自然保护区南段生物走廊带维管植物名录(2)[J].云南师范大学学报,2008,28(4):52-59.
[10] 刘经伦, 崔明昆, 汪建云,等.高黎贡山南段种子植物区系科属的两种不同排序方法及其意义[J].云南师范大学学报,2012,32(3):66-73.
[11] 李锡文.中国种子植物区系统计分析[J].云南植物研究,1996,18(4):363-384.
[12] 应俊生.中国种子植物特有属的分布区学研究[J].植物分类学报,1996,34(5):479-485.
[13] 刘经伦,崔明昆,汪建云,等.高黎贡山南段种子植物区系的特有现象[J].广西植物,2013,33(2):269-274.
