深圳梧桐山毛棉杜鹃根内真菌的分离与鉴定
2015-04-29刘亚等
刘亚等
摘要[目的] 对深圳梧桐山毛棉杜鹃根内真菌进行分离、鉴定,为深圳梧桐山毛棉杜鹃优良菌株的筛选奠定基础。[方法]采用根段直接培养法从毛棉杜鹃的根内分离得到14个菌株,采用rDNAITSPCR分子生物學方法对所得菌株进行鉴定。[结果]深圳梧桐山毛棉杜鹃根内真菌丰富,所获的14个菌株均属于子囊菌门(Ascomycota);除瓶头霉属(Phialocephala)和软盘菌属(Mollisa)外,其余菌株分别与Bionectria ochroleuca、Gloeotinia temulenta、Aspergillus sydowii、Paecilomyces javanicus和Fusarium oxysporum同源或近缘,均首次在杜鹃花科植物根内发现。[结论]该研究为毛棉杜鹃菌根化育苗技术的优良菌株筛选研究奠定基础。
关键词毛棉杜鹃;根内真菌;菌落形态; rDNA ITS序列
中图分类号S718.81文献标识码
A文章编号0517-6611(2015)29-009-04
毛棉杜鹃(Rhododendron moulmainense)为杜鹃花科杜鹃属马银花亚属常绿灌木或小乔木,其花朵艳丽,花色丰富,具有较高的观赏价值。深圳市梧桐山风景名胜区有较大面积的毛棉杜鹃天然林,盛花期整体景色壮丽,是该景区的特色景点。开展毛棉杜鹃资源的开发利用,不仅具有较高的经济效益,也具有较高的社会效益和生态效益。
杜鹃花科植物为典型的菌根依赖型植物,能与真菌形成具有圈状结构的菌根共生体,即杜鹃花菌根(Ericoid mycorrhiza)。不同种类的杜鹃花属植物或不同生态环境的同种杜鹃花植物,其根内真菌的种类和组成具有较大差异性。笔者采用根段培养法,对深圳梧桐山野生毛棉杜鹃根内真菌进行分离与培养,并运用rDNAITSPCR分子生物学方法进行菌株鉴定,探讨毛棉杜鹃菌根真菌多样性,以期为毛棉杜鹃菌根化育苗技术的优良菌株筛选研究奠定基础。
1材料与方法
1.1材料梧桐山自然风景区位于广东省深圳市东部,113°17′~114°18′E,22°23′~22°43′N。2014年3月下旬于梧桐山自然风景区采集一批毛棉杜鹃野生幼苗作根内真菌的分离研究。
1.2方法
1.2.1根内真菌的分离及培养。根内真菌分离培养基使用改良的马丁-孟加拉红培养基,菌株培养基使用马铃薯葡萄糖琼脂培养基(PDA)。选取毛棉杜鹃健壮纤细的根系,在清水中洗净泥土和杂质,沥干水分。在超净工作台下先在75%乙醇中浸泡15 s,无菌水漂洗3~5次;再在0.1%升汞中浸泡1~2 s,无菌水漂洗3~5次。把根系剪成长约0.5 cm的小段,用无菌滤纸吸干水滴,每个平板5条根段,置于分离培养基中培养,共50个根段。用接种棒蘸取少量最后一遍漂洗根系的无菌水,均匀涂抹于平板,作为对照排除杂菌。把培养根段和对照的平板置于25 ℃培养箱中黑暗培养14~28 d。
待菌落从根段中长出,用接种针挑取菌丝至PDA培养基上,培养7 d后观察菌落形态的一致性和均匀性。若菌落形态有差异,进行二次分离,如此反复直至纯化。把纯化后的菌株根据菌落形态特征进行描述记录和分类整理,包括菌落直径、颜色、质地、是否有液体分泌物、边缘、菌落外形等。
1.2.2菌株分子生物学鉴定。
菌株DNA提取参考庄彩云等方法。PCR扩增采用真菌通用引物,ITS1F:5′CTTGGTCATTTAGAGGAAGTAA3′,ITS4B:5′TCCTCCGCTTATTGATATGC3′。PCR扩增体系(25 μl):2×MightyAmp Buffer Ver.2(Mg2+,dNTP plus)(TaKaRa)12.5 μl;引物各0.8 μl;DNA模板20 ng;MightyAmp DNA Polymerase(1.25 U/μl)(TaKaRa)0.5 μl;ddH2O定容至25 μl。PCR反应循环参数:98 ℃预变性3 min;98 ℃变性30 s,50 ℃退火30 s,68 ℃延伸1 min,38个循环;68 ℃延伸7 min。
ITS区段PCR扩增产物的检测及测序:12 g/L PCR产物的Agarose在80 V电压下电泳40 min,BIORAD凝胶成像系统检测,若条带清晰则样品可用于测序。
PCR扩增产物测序由英潍捷基(上海)贸易有限公司完成。把得到的序列提交至GeneBank数据库进行Blast相似性比对,下载相似度较高的序列,经过ClustalX软件比对并校正后,采用MEGA6软件的NeighborJoining程序构建系统发育树。
2结果与分析
2.1菌落形态
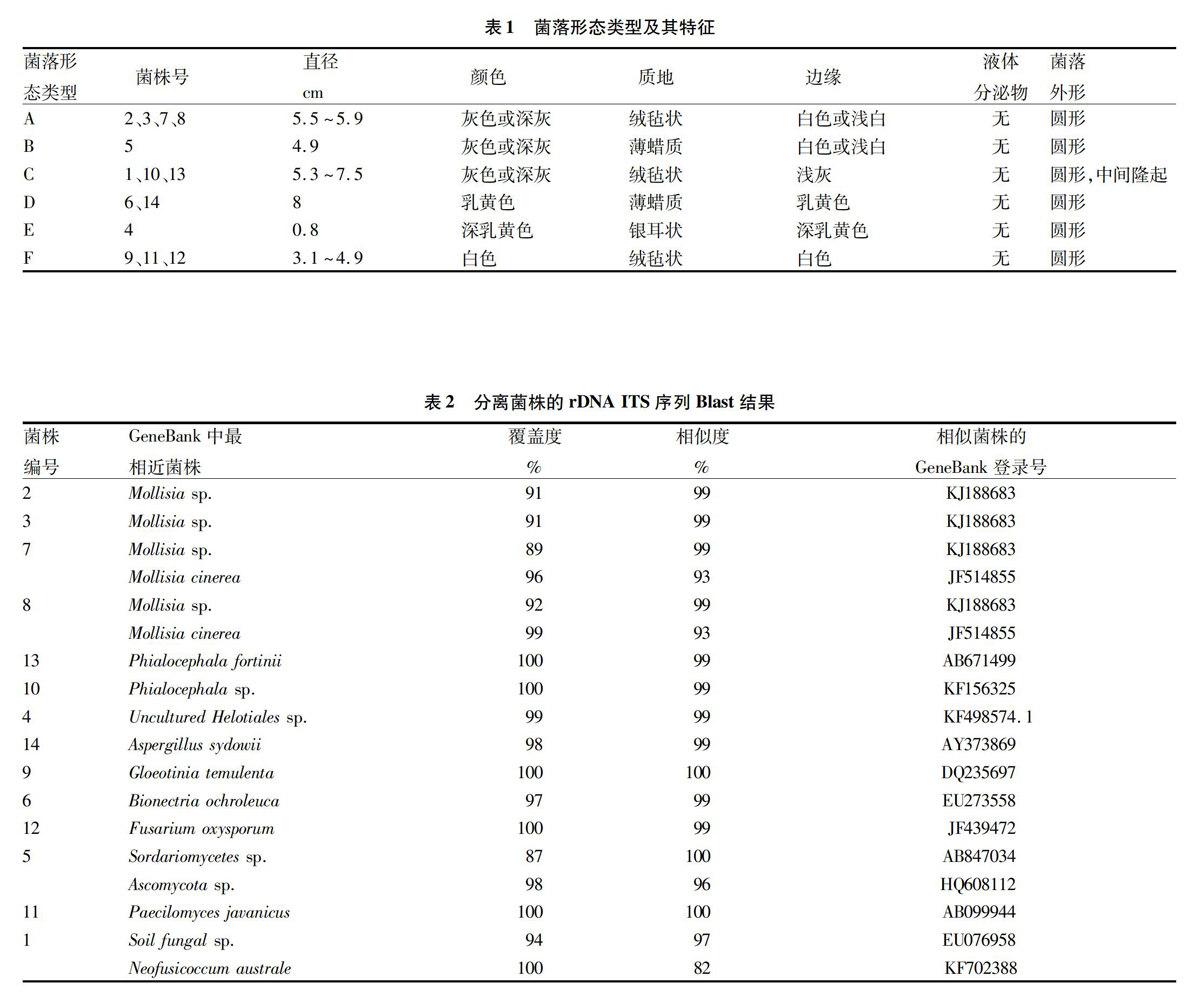
从野生毛棉杜鹃幼苗根内分离纯化14个菌株,按照菌落的形态特征分为6类(表1)。接种于PDF培养基上,置于25 ℃培养箱中暗培养21 d后,生长最快的菌株菌落直径可达8.0 cm,生长最慢的仅有0.8 cm;菌落颜色以灰色或深灰色为主,还有白色和黄色;质地多为绒毡状,还有薄蜡质和银耳状;均没有液体分泌物;少数菌落中间隆起。
43卷29期刘 亚等深圳梧桐山毛棉杜鹃根内真菌的分离与鉴定
2.2菌株ITS序列鉴定及系统发育分析
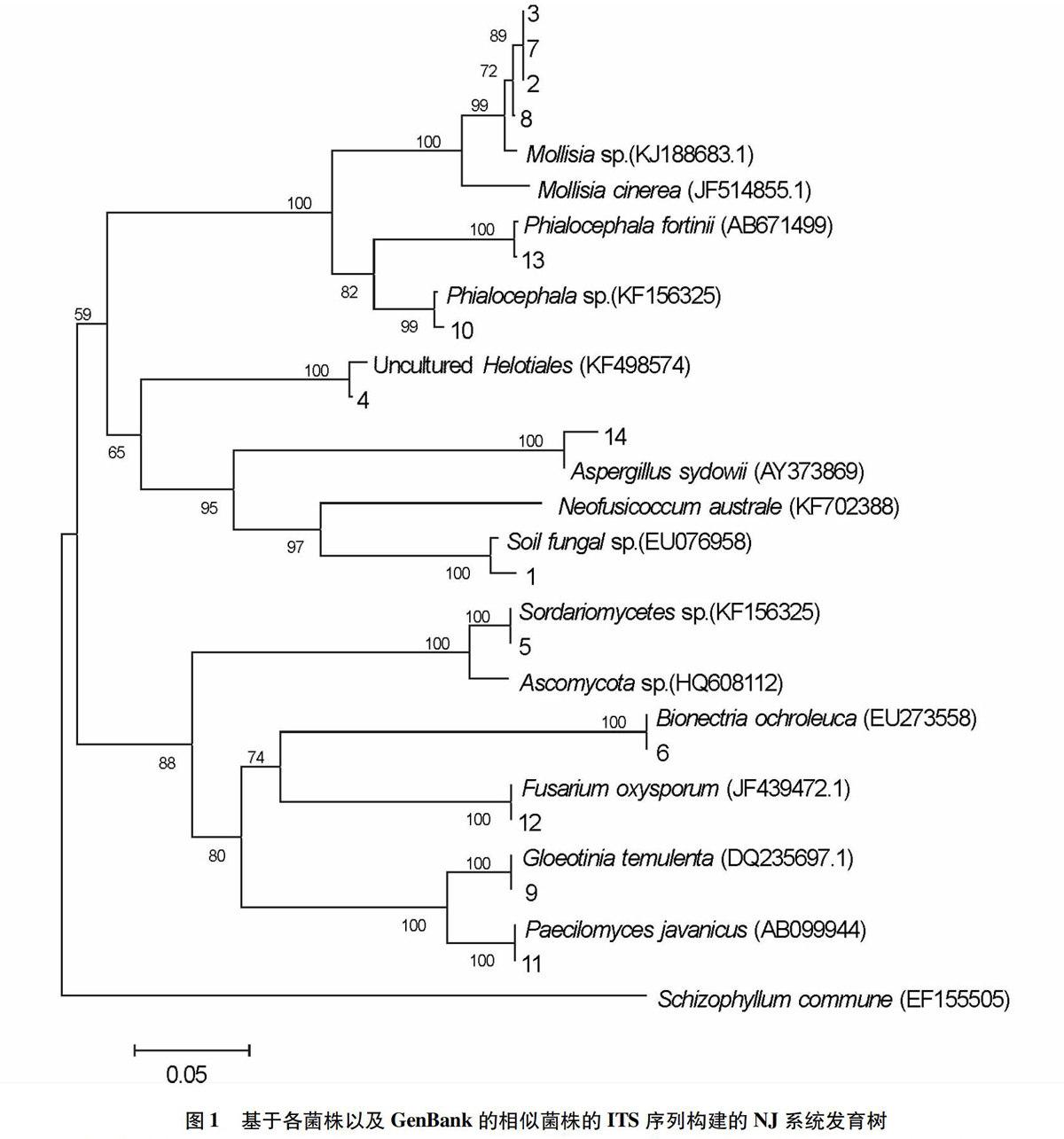
把测得的序列提交至GeneBank数据库进行Blast相似性比对(表2)。下载相似度最高及较高的序列,以亲缘关系相对较远的Schizophyllum commune(GeneBank登录号EF155505)为外群,构建NJ系统发育树(图1),14个菌株聚为11类。其中,菌株2、3、7、8与Mollisia聚在一起获得99%的支持率,序列相似度均达到99%;菌株7、8与Mollisia cinerea的序列相似度均达到93%。菌株13和10与Phialocephala 具有99%的序列相似度,菌株13与P. fortinii菌株(GeneBank登录号AB671499)的相似度为99%。菌株1和菌株4分别与GeneBank登录号EU076958 菌株 (Uncultured Helotiales sp.) 和KF498574.1菌株 (Sordariomycetes sp.) 具有较高的相似度(表2)。
此外,菌株6、9、11、12和14分别与Bionectria ochroleuca、Gloeotinia temulenta、Paecilomyces javanicus、Fusarium oxysporum、Aspergillus sydowii等菌株聚在一起,序列相似度在99%以上,表明它们与对应菌株同源或为同一菌种。
3结论与讨论
3.1结论
从深圳梧桐山毛棉杜鹃野生幼苗根段中分离得到14个内生真菌的菌株。依据菌落形态特征分为6类,菌落颜色以灰色或深灰色为主;基于rDNAITS序列分析和NJ系统发育树,14个菌株聚为11类,均隶属于子囊菌门,其中柔膜菌目菌株的数量(菌株2、3、7、8、13、10、4、9)最丰富,占菌株总数的57.1%。
3.2讨论
3.2.1菌株数量和菌株类型数量。
张春英从浙江、湖南等4个样地采集了24株云锦杜鹃(每个样地分别采集6株,取200个根段),从800个根段中共分离培养得到280个菌株。应用菌落形态、菌丝体显微特征和rDNAITS片段的RFLP图谱分析,确定了14个RFLP类型和3个未能鉴定的类型。在14个RFLP类型中,有7个类型与现有的杜鹃花类菌根菌亲缘关系密切,分别为Oidiodendron maius、Cryptosporiopsis ericae和5种尖苞树科植物的内生真菌的同源或近缘种;2个类型为黑色有隔内生真菌,即Phialocephala类。杨兵采用与张春英的研究方法,从上海、杭州、宁波3个地区18株毛鹃的600个根段内,分离培养得到366个菌株,确定了19个RFLP类型,其中9个类型为尖苞树科尖苞树属植物的内生真菌,另有8个未知的子囊菌和2个其他未知类型。刘振华从吉林、重庆、云南3个样地的牛皮杜鹃(Rhododendron chrysanthum)、小杜鹃(Rhododendron)、越橘(Vaccinium)等9种共24株杜鹃中,分离培养得到500个菌株,利用菌落形态结合rDNAITS序列分析鉴定出3种杜鹃花类菌根真菌Oidiodendron maius、Meliniomyces variabilis、Cryptosporiopsis ericae,1种黑色有隔内生真菌Phialocephala fortinii和2种尖苞树科植物内生真菌或其近缘种。
深圳梧桐山毛棉杜鹃自然群落中极少见到野生幼苗,采集的样本较少,但在50个根段内分离得到14个菌株,经过ITS序列分析被歸为11类不同菌种的同源或近缘种。虽然采样植株较少,菌株数量不多,但菌株类型较丰富,反映了梧桐山毛棉杜鹃根内真菌也具有较高的菌种多样性。
3.2.2菌落形态分类和ITS分子分类结果的比较。
传统的真菌鉴定方法依据菌落形态特征及菌体的光学显微结构,但真菌形态结构复杂,而且不少菌种的菌落形态特征和生理生化指标在不同培养基上会发生变化。另一方面,大多数菌根真菌在培养过程中不产生孢子,更增加了菌种鉴定的困难。随着GeneBank数据库的建立和生物信息学的发展,应用rDNAITS序列分析进行真核生物的系统发育、分类鉴定和多样性研究已成为一种有效且快速的方法。
其他学者对杜鹃花类菌根菌的研究也表明不同菌落形态的菌株的rDNA序列具有同源性。在该研究所获得的14个菌株中,根据菌落形态特征和rDNA ITS序列分析分别划分为6类和11类,所获结果稍有差异。其中,同属菌落形态C类的菌株1、10、13,在NJ系统发育树上并未聚在一起,菌株1与菌株10和13的分支距离很远。分别同属菌落形态D类的菌株(菌株6、14)和菌落形态F类的菌株(菌株9、11、12),也在NJ系统发育树上分离。然而,有些菌落形态类似的菌株,如菌株2、3、7、8,在rDNA序列上具有一致性,反映了它们是同一种或近缘种。由此可见,rDNAITS序列分析结果更可信。
3.2.3菌株鉴定。
目前国内外已有研究表明,从杜鹃花科植物中分离并回接确认为杜鹃花类菌根真菌的都属于子囊菌类。该研究对毛棉杜鹃进行菌根分离得到的14个菌株均属于子囊菌门,分别属于柔膜菌目、肉座菌目、散囊菌目。其中菌株2、3、7、8与皮盘菌科软盘菌属Mollisia同源;菌株7和8与M. cinerea的序列相似度达93%,反映了它们具有较高的同源性。杨秀丽[11]和刘凤红等[12]分别在大兴安岭地区的杜香(杜鹃花科杜香属)和长白山地区的笃斯越橘根内也分离得到同属Mollisia minutella的同源菌株;刘仁阳等在梵净山雷山杜鹃的根内也分离得到M. cinerea的同源菌株,反映了Mollisia属菌种可与杜鹃花科植物形成菌根共生体。
该研究还分离获得了2个与Phialocephala具有较高同源性的菌株,其中1个与P. fortinii具有99%的序列相似度(菌株13)。Phialocephala属的真菌通常被归为深色有隔内生真菌[13]。近年有关Phialocephala属的真菌在杜鹃花科植物根内出现的研究已有报道,如杨秀丽[11]、刘凤红等[12]和刘振华[9]分别报道了Phialocephala属同源或近缘真菌为大兴安岭地区和长白山地区杜鹃花科植物菌根菌的优势种。
该研究未分离到Hymenoscyphus eircae、Oidiodendron sp.、Cryptosporiopsis ericae、Meliniomyces variabilis等典型的杜鹃花类菌根菌种。值得关注的是分离获得的Bionectria ochroleuca、Gloeotinia temulenta、Aspergillus sydowii(聚多曲霉)、Paecilomyces javanicus(爪哇拟青霉菌)和Fusarium oxysporum(尖孢镰刀菌)的同源或近缘种,均是首次在杜鹃花科植物根内出现。其中,Bionectria ochroleuca对多种植物病原菌具有拮抗作用[14-15];Gloeotinia temulenta和Fusarium oxysporum通常为植物致病性真菌[16-18];Paecilomyces javanicus为昆虫天敌真菌[19]。该研究回接了2个曲霉菌株到毛棉杜鹃幼苗上,未见有致病性且具有促生作用(另文报道),它们与毛棉杜鹃的生态关系还有待进一步研究。
参考文献
[1] 刘振华,姚娜,杨凯,等.树枫杜鹃菌根真菌分离与鉴定[J].林业科学研究,2012,25(6):795-797.
[2] 刘仁阳,欧静,李冠楠,等.梵净山雷山杜鹃根部真菌分离与鉴定[J].西北农业学报,2014,23(4):178-185.
[3] 徐胜男,王济,张凌云,等.马缨杜鹃菌根真菌的种类组成[J].贵州农业科学,2014,42(6):76-78,79.
[4] 张春英,尹丽娟,王胤,等.一种简捷的分离杜鹃花类菌根真菌的方法[J].生物技术,2007,37(6):28-32.
[5] 庄彩云,李潞滨,胡陶,等.适用于rDNA ITS分析的兰属菌根真菌培养及DNA提取方法[J].北京农学院学报, 2007,22(3):4-6.
[6] GARDES G M,BRUNS T D.ITS primers with enhanced specificity for basidiomycetes application to the identification of mycorrhizae and rusts[J].Molecular ecology,1993,2(2):113-118.
[7] 张春英.云锦杜鹃菌根及其菌根真菌多樣性研究[D].北京:北京林业大学,2008.
[8] 杨兵.毛鹃菌根真菌多样性初步研究[D].郑州:河南农业大学,2010.
[9] 刘振华.杜鹃花菌根真菌分离鉴定及多样性分析[D].北京:中国林业科学研究院,2010.
[10] 杨秀丽,闫伟.笃斯越橘菌根真菌多样性[J].微生物学报,2015,55(2):214-219.
[11] 杨秀丽.大兴安岭兴安落叶松森林生态系统菌根及其真菌多样性研究[D].呼和浩特:内蒙古农业大学,2010.
[12] 刘凤红,程显好,顾亮,等.长白山野生笃斯越橘根系内生菌资源调查[J].山东农业科学,2015,47(1):41-46.
[13] 刘茂军,张兴涛,赵之伟.深色有隔内生真菌(DSE)研究进展[J].菌物学报,2009,28(6):888-894.
[14] 刘琴英,蒋冬花,齐育平,等.淡色生赤壳菌Bo-1菌株拮抗物质的分离纯化、解析及活性分析[J].植物保护学报,2014,41(1):41-44.
[15] 常伟,宋福强,王占斌,等.引进菌株Bionectria ochroleuca对几种植物病原真菌的拮抗效应[J].中国农学通报,2014,30(18):166-170.
[16] 陈石,李春雨,易干军,等.尖镰孢菌致病机理研究进展[J].中国农学通报,2011,27(13):74-78.
[17] 肖敏,曾向萍,严婉荣,等.海南豇豆枯萎病病原鉴定及生物学特性初步研究[J].基因组学与应用生物学,2015,34(2):345-349.
[18] 汪静,梁宗锁,康冰,等.文山三七根腐病病原真菌的鉴定与药剂防治[J].西北林学院学报, 2015,30(1):158-163.
[19] 马骏,刘强,荣湘民,等.防治斜纹夜蛾的爪哇拟青霉菌株筛选及培养[J].湖南农业大学学报(自然科学版), 2007,33(4):466-470.
