北二星蝽Eysarcoris aeneus的DNA分类学研究(半翅目:蝽科:二星蝽属)
2015-04-19赵清李敏孙溪张虎芳
赵清,李敏,孙溪,张虎芳*
(1.山西农业大学 农学院,山西 太谷 030801;2.太原师范学院,山西 太原 030000;3.南开大学 昆虫学研究所,天津 300071)
北二星蝽Eysarcoris aeneus的DNA分类学研究(半翅目:蝽科:二星蝽属)
赵清1,李敏2,孙溪3,张虎芳1*
(1.山西农业大学 农学院,山西 太谷 030801;2.太原师范学院,山西 太原 030000;3.南开大学 昆虫学研究所,天津 300071)
本文基于形态和分子数据证明北二星蝽Eysarcorisaeneus和尖角二星蝽E.parvus为同一个种。通过扩增中国二星蝽属的4个种,尖角二星蝽E.parvus,北二星蝽E.aeneus,二星蝽E.guttigerus和广二星蝽E.ventralis的线粒体COI基因部分片段(约560 bp),共获得28条序列。计算其种间和种内遗传距离,并基于邻接法(NJ)、最大似然法(ML)和贝叶斯推断法(BI)进行系统进化分析;同时,解剖这些种的生殖节发现,尖角二星蝽和北二星蝽之间的生殖节差异远小于与本属内其他种间的生殖节差异,因此,基于遗传距离、系统发育分析以及生殖节形态差异的结果均显示北二星蝽和尖角二星蝽之间的差异远小于这两个种与属内其他种之间的差异。我们得出(1)北二星蝽和尖角二星蝽为同一种;(2)DNA分类学可以利用线粒体COI基因解决有争议的分类学问题,同时综合利用形态和分子数据能够为此类问题的解决提供新的方法和视角。
半翅目;蝽科;二星蝽属;北二星蝽;DNA分类学
二星蝽属(EysarcorisHahn,1834)隶属于半翅目Hemiptera异翅亚目Heteroptera蝽科Pentatomidae蝽亚科Pentatominae。二星蝽成虫、若虫可通过吸食寄主植物的嫩枝、幼茎或叶片的汁液,致植株生长发育受阻,籽粒不饱满,部分种类可危害水稻、麦类、棉花、大豆、胡麻、高粱、甘薯、茄子等作物,是重要的农业害虫[1]。据世界主要产稻国家研究报道,所有为害稻类的昆虫中,二星蝽属的广二星蝽E.ventralis,北二星蝽E.aeneus和E.trimaculatus位居前列[2,3]。本属昆虫分布较广,在蝽科中由于体小,小盾片基角处具黄白色光滑小圆斑而极易鉴定[4]。
其中,北二星蝽E.aeneus和尖角二星蝽E.parvus在形态上极为相近,唯有前胸背板侧角不同(图1中1~8),前者侧角末端圆钝且伸出较短,后者末端尖锐呈针状且伸出较长;生殖器差异较小,仅在阳茎的系膜顶叶、系膜侧叶的长短、骨化程度有区别(图2 Ie,IIe);另外,这2种在有些地方为同域分布(表1)。因此该2种的分类地位一直有争议,Josifov(1978)将此2种合并为北二星蝽E.aeneus,将尖角二星蝽E.parvus做次异名处理[5],但是未提供详细的证据。对于这些形态特征有变异的分类处理,除了借助外部形态特征及雄性外生殖器的比较外,还应该提供其他证据,如分子证据。
在过去的几年中,DNA条形码作为一个分子生物学不可或缺的方法吸引了人们的注意力。它是利用标准化的DNA片段进行物种鉴定的方法[6],能够帮助非分类学专家进行物种鉴定。线粒体细胞色素C氧化酶基因COI具有引物通用性高、进化速率快、变异速率在种内相对小,在种间又相对大[6,7]等优势,其5’端648 bp的片段成为动物分类与鉴别的理想DNA 条形码[8~10]。另外,DNA条形码在提高物种鉴定效率、发现新种和隐存种以及研究系统发育和进化关系等方面提供新的思路和研究工具[11]。因此,DNA条形码被越来越多地应用于分类,但许多学者建议COI条形码被用于新种描述或新异名时应与形态、地理分布等相结合。
本文基于形态和分子证据探究北二星蝽和尖角二星蝽两者的关系。形态学方面解剖雄性生殖节并进行比较,分子生物学方面通过扩增线粒体COI基因序列(约560 bp),计算其种内种间遗传距离并构建系统进化树。预期基于两种数据的研究结果推断(1)北二星蝽和尖角二星蝽是否为同一种;(2)DNA条形码是否可以有效地解决有争议的分类学问题。

图1 北二星蝽、尖角二星蝽、广二星蝽和二星蝽的成虫背腹面观Fig.1 Habitus of E.aeneus,E.parvus,E.ventralis and E.guttigerus,dorsal and ventral view注:1~2.北二星蝽(1.背面观; 2.腹面观); 3~4.尖角二星蝽(3.背面观; 4.腹面观); 5~6.广二星蝽(5.背面观; 6.腹面观); 7~8.二星蝽(7.背面观; 8.腹面观)Note:1~2.E.aeneus (1.dorsal view; 2.ventral view); 3~4.E.parvus (3.dorsal view; 4.ventral view); 5~6.E.ventralis (5.dorsal view; 6.ventral view); 7~8.E.guttigerus (7.dorsal view; 8.ventral view)
1 材料与方法
1.1 形态学
雄性生殖器取自回软的标本,将其放入2%的NaOH溶液中并在灯下烤30 min,之后置于清水中数分钟使阳茎充分膨胀。观察时需让阳茎膨胀充分至完全伸出阳茎鞘之外,必要时可以辅助一些器械。观察完之后,将生殖器保存于甘油当中,和原标本放在一起。成虫的背腹面照片采用Nikon SMZ1000照相。所用标本均来自于南开大学昆虫标本馆。
1.2 分子生物学
选取尖角二星蝽16头、北二星蝽8头、广二星蝽和二星蝽各2头,试验中所得数据均来自于酒精浸制标本提取(标本详细信息见表1)。
基因组的提取采用试剂盒(康为世纪CW0546)。证据标本除胸部肌肉用于提取基因外,其他部分酒精保存,存放于南开大学昆虫所。PCR扩增采用如下引物:YT1F(5-AAACTATTAACCTTCAAAG-3)/HCO2198(5-GTCGTGGAAGAAGATTAGTTGTTCTAT-3)。

图2 北二星蝽、尖角二星蝽、广二星蝽和二星蝽的雄性生殖节图Fig.2 Male genitalia of E.aeneus,E.parvus,E.ventralis and E.guttigerus注:I: 北二星蝽; II: 尖角二星蝽; III: 广二星蝽; IV: 二星蝽; a: 生殖囊背面观; b: 生殖囊腹面观; c:阳基侧突端面观; d: 阳基侧突侧面观; e: 阳茎Note:I: E.aeneus; II: E.parvus; III: E.ventralis; IV: E.guttigerus; a: dorsal view of pygophore; b: ventral view of pygophore; c: apical view of paramere; d: lateral view of paramere; e: aedeagus
PCR扩增的反应体积为50 μL,2 μL上游和下游引物,2~5 μL的基因组模板,25 μL的Mix预混液(康为世纪CW0556),加ddH2O补足至50 μL。PCR反应为92 ℃变性40 s,45 ℃退火40 s,72 ℃延伸1 min共5个循环,之后再92 ℃变性40 s,51 ℃退火40 s,72 ℃延伸1 min,共33个循环,反应前94 ℃预变性2 min,反应后72 ℃再延伸8 min,最后4 ℃保温。
经PCR扩增后,用1.0%的琼脂糖凝胶电泳检测,若条带整齐明亮,将PCR产物送至金维智测序公司直接测序。
获得数据后,使用BioEdit7.0[12]打开峰图文件,检查序列的单一性,对得到的每条序列进行校对、拼接,并结合峰图进行修正,确定准确无误后将其另存为Fasta格式文件;然后导入Mega 6.0[13]软件中,用无脊椎动物线粒体遗传密码子表对COI基因进行翻译,适当调整阅读框检查有无终止密码子存在,以排除假基因的干扰。将所得序列在NCBI上进行BLAST搜索,得到的同源序列大多数为蝽科线粒体基因组的COI片段。在BioEdit中,用ClastalW进行比对,去掉两端不整齐的序列。用DAMBE4.0.36[14]计算单倍型个数,不同的单倍型用于遗传距离的计算和系统进化树的分析。用Mega6.0基于Kimura-2-Parameter[15]计算种间和种内遗传距离。
系统进化树基于邻接法(NJ)[16],最大似然法(ML)和贝叶斯推断法(BI)构建。最大似然法和邻接法由PAUP* 4.10b10[17]windows版本执行。贝叶斯法由MrBayes 3.1[18]运算,最适合数据集的进化模型利用Modeltest 3.7[19]测得采用GTR+I模型,运行1 000 000代,每1 000代抽样一次,“burn in”25%的样本后构建系统进化树。所构建的进化树均采用TreeView1.6.6查看编辑。
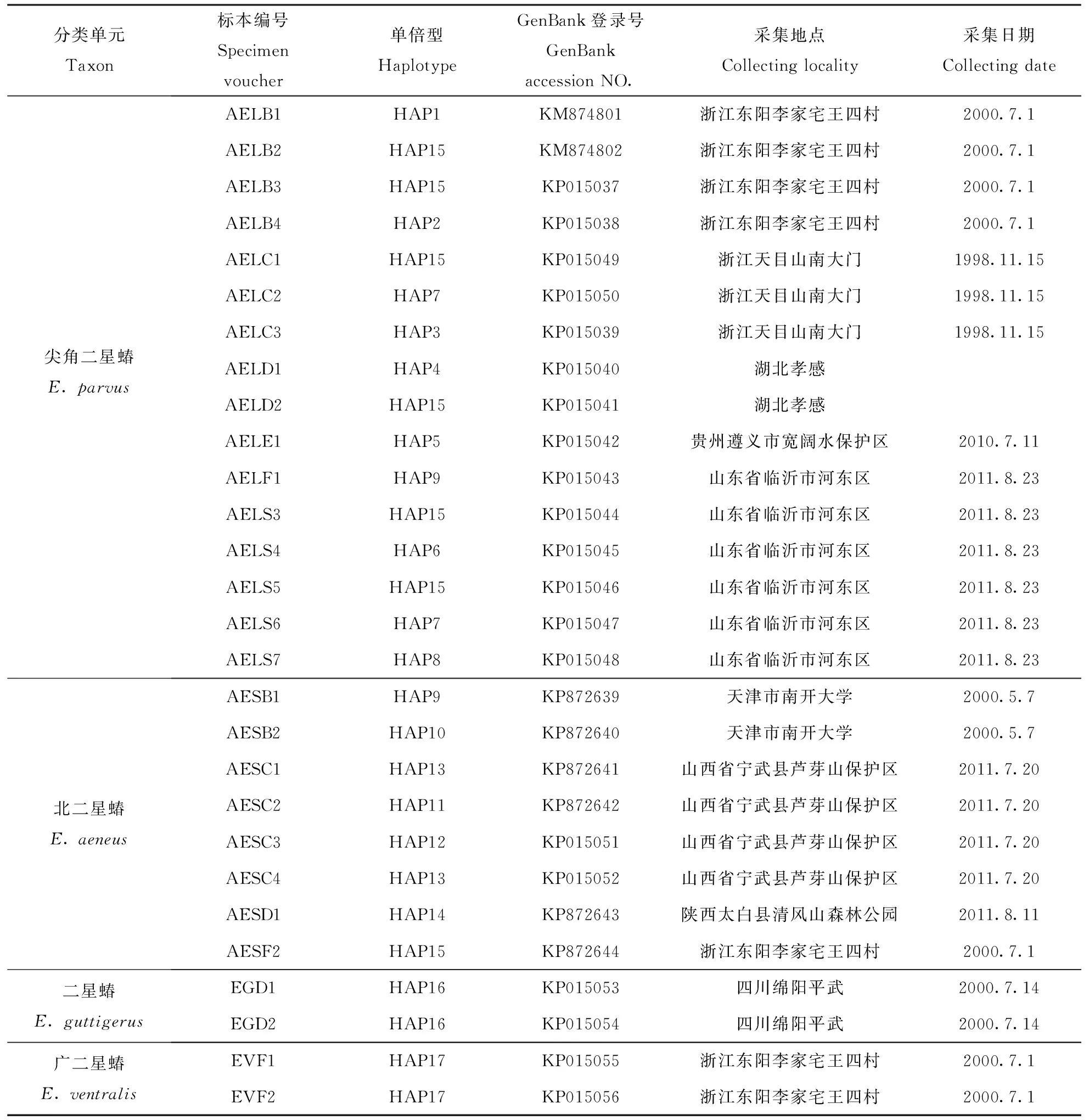
表1 实验所用标本的详细信息
2 结果与分析
2.1 形态学数据
北二星蝽Eysarcorisaeneus(Scopoli,1763)(图1)
CimexaeneusScopoli,1763: 122.
CimexfucatusRossi,1790: 235.
CimexperlatusFabricius,1794: 125.
Eysarcorisperlatusvar.spinicollisPuton,1881: 76; 1881: 56
Eysarcorisperlatusvar.ventralisHorvath,1882: 219.
EysarcorisparvusUhler,1896: 258.
Eysarcorisaeneusvar.peeziTamanini,1961: 122.
Eysarcorisaeneusf.nigriventrisStichel,1962: 781.
鉴别特征:头侧叶稍长于中叶,具铜绿色金属光泽。胝区具一黑斑,方形。前胸背板侧角略伸出,或呈尖刺状伸出。小盾片倒三角形,基角各具一长椭圆形的斜置黄白斑,末端外缘常具3个小黑斑。腹部腹面中央均匀漆黑,边缘呈锯齿状,两侧各具一长短不等的黑纵纹。
形态特征:体卵圆形,淡黄褐色,密被黑色粗刻点。
头:黑色,具铜绿色光泽,端部略下倾,基部中央具浅色短纵纹;侧叶末端圆钝,稍长于中叶;复眼褐色,单眼红色;触角黄褐色,第IV节端部及第V节黑。头部腹面黑,具铜绿色金属光泽;喙黄褐色,末端伸达后足基节处。
胸:前胸背板宽大于长,前半略下倾,后半隆拱;胝区具一近方形黑斑;侧角末端圆钝,略伸出,部分个体侧角呈尖刺状伸出较长。小盾片倒三角形,基角具一斜列的长椭圆形黄白斑;末端圆钝,不超过前翅革片末端,边缘常具3个小黑点斑。各胸部侧板黄褐色,具黑色粗刻点;臭腺沟缘呈耳状,超过后胸侧板一半。足淡黄褐色,腿节亚端部具一小黑斑。
腹:腹部腹面淡黄褐色,基部中央无前指的突起;中央具黑色纵带,约占腹部1/3,边缘锯齿状,其两侧各具一长短不等的黑纵纹。气门黑色。
雄虫生殖节(图2):生殖囊杯状,宽大于长,腹后缘中央呈宽“V”状内凹,两侧呈脊状加厚,略向腹面翻卷(图2 Ia,IIa,Ib,IIb)。阳基侧突较细长,末端指状(图2 Id,IId)。阳茎端较短,向端部渐细;系膜顶叶深二叉状,内侧骨化;系膜侧叶亦分叉,部分骨化(图2 Ie,IIe)。
此种的前胸背板侧角变异较大:部分个体侧角末端圆钝,伸出体外较短;部分个体侧角末端呈尖刺状,水平伸出较长。解剖其雄性生殖节发现这两类个体的生殖节差异与其他种比起来差异小很多,如生殖囊背腹后缘的弯曲程度(图2:Ia,Ib,IIa,IIb),阳茎端的长短、系膜顶叶的形状及的长短、系膜侧叶的骨化程度及长短(图2:Ie,IIe)等在种间差异很大,而在这两类个体之间变化相对小。
2.2 分子数据
2.2.1 遗传距离分析结果
经测序共获得28条序列,用DAMBE统计得出有17个单倍型,其中尖角二星蝽16条序列10个单倍型,北二星蝽8条序列7个单倍型,而北二星蝽和尖角二星蝽的部分序列出现共享单倍型“HAP9”“HAP15”。利用MEGA6.0分析COI序列(558bp)中共有保守位点(Conserved)453个,变异位点(Variable)105个,简约信息位点(Parsim-info)101个,单突变子(Singleton)4个。统计碱基组成显示,密码子第一位T、C、A、G平均含量分别为40%、8.2%、48.3%、3.5%,密码子第二位T、C、A、G平均含量分别为25%、17.5%、30.7%、26.9%,密码子第三位T、C、A、G平均含量分别为45%、25.6%、13.4%、16.2%,而G+C平均含量为33.6%,远低于A+T的含量,符合线粒体基因的高A-T含量的特性。
利用MEGA6.0对COI基因基于K-2-P计算种内/种间遗传距离(表2)。由表2可以看出广二星蝽与北二星蝽、尖角二星蝽、二星蝽的遗传距离分别为0.163 7、0.165 8、0.110 2,而尖角二星蝽和北二星蝽的种间遗传距离为0.0119。根据Hebertetal.(2004)提出的标准,种间遗传距离应大于种内距离的10倍,所以尖角二星蝽和北二星蝽应该为同一个种。

表2 基于COI基因种内-种间遗传距离Table 2 The intra-and inter-species genetic distance(K2P)of COI
2.2.2 系统发育分析
我们选取本属的广二星蝽E.ventralis作为外群,系统进化树的结果显示,基于NJ、ML和BI构建的系统进化树结果基本一致,且节点支持率较高(图3~图5)。在各进化树中二星蝽E.guttigerus及早的分出,北二星蝽和尖角二星蝽聚为一大支,且节点支持率均较高,为100或1.00,虽支系内部略有差异,但总体来说无论是基于哪种算法的系统进化结果均不支持二星蝽和尖角二星蝽的单系性。

图3 基于COI基因构建的Bayesian系统进化树Fig.3 Phylogenetic tree of Bayesianbased on COI gene

图4 基于COI基因构建的NJ系统进化树Fig.4 Phylogenetic tree of NJ based on COI gene

图5 基于COI基因构建的ML系统进化树Fig.5 Phylogenetic tree of ML based on COI gene
3 结论与讨论
Josifov(1978)虽然将北二星蝽和尖角二星蝽合并为一个种,但是文中未提供详细证据,亦未提供雄性外生殖节图。本文通过解剖北二星蝽与尖角二星蝽的雄性生殖节,详细比较两种生殖节各部分结构,并与广二星蝽及二星蝽的雄性生殖节结构进行比较,结果显示尖角二星蝽和北二星蝽的形态差异与同属其他种相比差异极小,不足以达到种的界限。同时扩增了二星蝽属内4个种的COI基因,计算其种内种间遗传距离,结果显示北二星蝽与尖角二星蝽间的种间遗传距离为0.0119,与本属内的种内遗传距离相似,且与其他种的种间遗传距离在0.139 1~0.165 8之间,如此大的差距,给予Josifov(1978)提出的次异名处理提供了强有力的证据支持。另外,基于三种算法构建的系统进化树结果均不支持两者的单系性。因此,无论从形态数据还是分子数据我们均能证明尖角二星蝽和北二星蝽为同一种。
同时,该研究也证明,DNA分类学能够为传统分类学服务。当遇到形态数据与分子数据有异议时,我们需要重新审视鉴定结果,还应该结合生态学、地理分布等来共同分析其物种界限。毫无疑问,DNA 条形码可以帮助一些毫无头绪的属种缩小鉴定范围;同时,传统分类学又能从形态上对分子结果进行检测。在研究过程中,我们必须将两种方法相互补充、完美结合才能最大限度发挥DNA分类学的优势。
[1]章士美.中国经济昆虫志(第50册)[M].北京: 科学出版社.1995:169.
[2]Lee JG,Hong SS,Kim JY,et al.Occurrence of stink bugs and pecky rice damage by stink bugs in paddy fields in Gyeonggi-do,Korea[J].Korean Journal of Applied Entomology,2009,48(1): 37-44.
[3]Nasiruddin M,Roy RC.Rice field insect pests during the rice growing seasons in two areas of Hathazari,Chittagong[J].Bangladesh Journal of Zoology,2012,40(1): 89-100.
[4]萧采瑜.中国蝽类昆虫鉴定手册,第I册(半翅目,异翅亚目)[M].北京: 科学出版社,1977: 76-89.
[5]Josifov MV,Kerzhner IM,Heteroptera aus Korea.II.Teil (Aradidae,Berytidae,Lygaeidae,Pyrrhocoridae,Rhopalidae,Alydidae,Coreidae,Urostylidae,Acanthosomatidae,Scutelleridae,Pentatomidae,Cydnidae,Plataspidae)[J].Fragmenta Faunistica,1978,23(9): 137-196.
[6]Hebert PDN,Cywinska A,Ball SL,DeWaard JR.Biological identifications through DNA barcodes[J].Proceedings of the Royal Society of London Series B-Biological Sciences,2003a,270: 313-321.
[7]Hebert PDN,Ratnasingham S,deWaard JR.Barcoding animal life: cytochrome oxidase subunit 1 divergences among closely related species[J].Proceedings of the Royal Society of London Series B-Biological Sciences,2003b,270: S96-S99.
[8]Hebert PDN,Stoeckle MY,Zemlak TS,et al.Identification of birds through DNA barcodes[J].Plos Biology,2004,2(7): 1657-1663.
[9]Ashfaq M,Akhtar S,Khan AM,et al.DNA barcode analysis of butterfly species from Pakistan points towards regional endemism[J].Molecular Ecology Resources,2013,13(5): 832-843.
[10]Yang F,Shi ZY,Bai SL,et al.Comparative studies on species identification of Noctuoidea moths in two nature reserve conservation zones (Beijing,China) using DNA barcodes and thin-film biosensor chips[J].Molecular Ecology Resources,2014,14(1): 50-59.
[11]Agnarsson I,Kuntner M.Taxonomy in a changing world: seeking solutions for a science in crisis[J].Systematic Biology,2007,56: 531-539.
[12]Hall TA.Bioedit: a user-friendly biological sequences alignment editor and analysis program for Windows 95/98/NT [J].Nucleic Acids Symposium Series,1999,41: 95-98.
[13]Tamura K,Stecher G,Peterson D,et al.MEGA6: Molecular Evolutionary Genetics Analysis version6.0 [J].Molecular Biology and Evolution,2013,30(12):2725-2729.
[14]Xia X,Xie Z.DAMBE: software package for data analysis in molecular biology and evolution [J].Journal of Heredity,2001,92(4): 371-373.
[15]Song H,Buhay JE,Whiting MF,et al.Many species in one: DNA barcoding overestimates the number of species when nuclear mitochondrial pseudogenes are coamplified [J].Proceedings of the National Academy of Sciences of the United States of America,2008,105: 13486-13491.
[16]Saitou N,Nei M.The neighbor-joining method: a new method for reconstructing phylogenetic trees [J].Molecular Biology and Evolution,1987,4: 406-425.
[17]Swofford DL.PAUP*: Phylogenetic Analysis Using Parsimony (and other methods).Version 4 (beta 10) [M].Sinauer Associates,Sunderland,Massachusetts,2003:94-96.
[18]Huelsenbeck JP,Ronquist F.MrBayes: Bayesian inference of phylogeny [J].Bioinformatics,2001,17: 754-755.
[19]Posada D,Crandall KA.Modeltest: testing the model of DNA substitution [J].Bioinformatics,1998,14: 817-818.
(编辑:武英耀)
Study on DNA Taxonomy ofEysarcorisaeneus(Hemiptera: Pentatomidae:Eysarcoris)
Zhao Qing1,Li Min2,Sun Xi3,Zhang Hufang1*
(1.CollegeofAgronomy,ShanxiAgriculturalUniversity,TaiguShanxi030801,China;2.TaiyuanNormalUniversity,TaiyuanShanxi030000,China;3.CollegeofLifeSciences,NankaiUniversity,Tianjin300071,China)
This paper based on morphology and molecular data to prove thatEysarcorisaeneus andEysarcorisparvusare the same species.We performed on 28 specimens,amplified about 560bp segment of the mitochondrial cytochrome coxidase I gene (COI) of 4 species,E.parvus,E.aeneus,E.guttigerusandE.ventralis.Computed the inter- and intra-species genetic distances and constructed the phylogenetic trees based on NJ,ML and BI methods.At the same time,we dissected the male genitalia of these species,compared their differences.All the results of the genetic distances,phylogenetic analysis and genitalia morphological characters showed that the difference and the genetic distance betweenE.aeneusandE.parvusmuch less than the difference amongE.aeneus,E.ventralisandE.guttigerus.So we concluded (1)E.aeneusandE.parvusare the same species; (2) DNA barcoding can resolve the argued taxonomy problems using the partial segment of mitochondrialCOIgene.The fact that based on molecular and morphological data can provide the new ideas and new perspective to resolve the taxonomy.
Heteroptera;Pentatomidae;Eysarcoris;Eysarcorisaeneus;DNA taxonomy
22015-03-10
2015-04-04
赵清(1985-),女(汉),山东临沂人,讲师,博士后,研究方向:DNA分类学
*通讯作者:张虎芳,教授,硕士生导师;Tel: 0354-6288225;E-mail: zh_hufang@sohu com
国家自然科学基金(31440078);山西省教育厅高等学校科技创新基金(2014131);山西农业大学科技创新基金(2014017)
S433.3
A
1671-8151(2015)03-0241-08
