山羊Agouti基因启动子活性区域探究
2015-04-11张永改张大山刘伟兰李祥龙周荣艳李兰会
张永改,郭 敏,张大山,刘伟兰,李祥龙*,周荣艳,李兰会
(1 河北科技师范学院,河北 秦皇岛,066004;2 河北农业大学;3 河北省定州市第二中学)
山羊Agouti基因启动子活性区域探究
张永改1,2,3,郭 敏1,张大山1,刘伟兰2,李祥龙1*,周荣艳2,李兰会2
(1 河北科技师范学院,河北 秦皇岛,066004;2 河北农业大学;3 河北省定州市第二中学)
对前期拼接获得的12 847 bp山羊Agouti基因序列(EF587236和JN651404)进行启动子活性区域预测,确定候选启动子区为6 450~7 100 bp,得到一个长度为2 537 bp的DNA片段,在该区域发现一个典型转录因子结合位点TATA框,并预测到HSF,SYR,GATA-1,Nkx-2,ADR1等多个转录因子结合位点;山羊基因组序列(登录号:AJPT01136617.1)中有两段序列与牛的预测启动子promoter A和promoter B同源性较高,据此设计引物扩增目的片段,同时构建12个荧光素酶报告基因质粒,瞬时转染人皮肤黑素瘤细胞A375和293T细胞检测目的片段是否具有Agouti基因启动子活性。结果表明,所构建的目的片段质粒与阴性对照相比无显著差异,说明所获得的目的片段不具有启动子活性。
山羊;Agouti基因;启动子
Agouti是指被毛具有黑色底色,尖端呈黄色特征皮毛的刺豚鼠,该词来源于南美土著印第安部落,最初将与此被毛特征相关的基因座命名为Agouti[1],现在动物毛色已成为辨别其外貌的主要特征之一[2]。Agouti是黑素皮质素受体信号转导的重要调控基因之一,在人(Homosapiens)、鸡(Gallusgallus)、牛(Bostaurus)、猪(Susscrofa)、绵羊(Ovisaries)和啮齿类等许多动物体内被发现[3~5]。Agouti基因含有多个等位基因,结构复杂,并且在不同物种中具有较高的同源性。山羊Agouti基因包含内含子1(531 bp)、外显子2(169 bp)、内含子2(1 318 bp)、外显子3(3 463 bp)和外显子4(177 bp)[6~8]。采用荧光定量PCR技术证明,不同毛色山羊皮肤组织中Agouti基因mRNA表达量为白色>黑色>棕色山羊,说明不同毛色山羊皮肤组织中的mRNA表达量不同,5’UTR变异对不同毛色山羊Agouti基因mRNA表达量有影响[9]。在所获得的山羊Agouti基因12 847 bp序列中对基因编码区上游进行启动子活性区域预测,利用TFSEARCH软件进行转录因子结合位点分析,检测到GATA-2,GATA-3,Oct-1,CdxA,HSF,GATA-1,STRE,ADR1,AP-2和AP-1等多个转录因子结合位点,确定候选启动子区为6 450~7 100 bp[10]。寻找Agouti基因启动子区域是研究Agouti基因重要环节,该工作对研究该基因遗传的分子生物学基础、指导山羊品种改良、选育及品种特异性研究具备重要理论和实践意义,同时能为我国地方山羊品种群体遗传的结构、分化及其多样性研究提供科学依据。
1 材料和方法
1.1 试验材料
本次试验所用样品来自河北农业大学动物科技学院实验室保存的太行山羊肝脏组织样品,利用酚氯仿抽提法提取山羊肝脏组织基因组DNA。PCR扩增、细胞培养与转染等所用试剂购自天根生化科技(北京)有限公司。
1.2 目的片段引物设计及扩增
利用Primer Premier 5.0软件,对扩增的2 537 bp序列分别设计不同长度的缺失片段,构建12个含有该片段的质粒(表1,分别以片段的起止位置命名)。在全长2 537 bp片段中不包含KpnⅠ,SmaⅠ,HindⅢ酶切位点,上述3种酶的酶切位点可作为参考的限制性酶切位点。在正反向引物的5’端分别引入KpnⅠ,SmaⅠ,HindⅢ酶切位点及相应的保护碱基以便于克隆。
PCR反应总体系为40 μL:灭菌水12 μL,2×Taq Mastermix 20 μL,模板基因组、cDNA 2 μL和正反引物各3 μL。
PCR反应条件:预变性,94 ℃,5 min;变性,94 ℃,30 s;退火30 s;延伸,72 ℃,根据片段大小确定延伸时间,35个循环;终延伸,72 ℃,10 min。
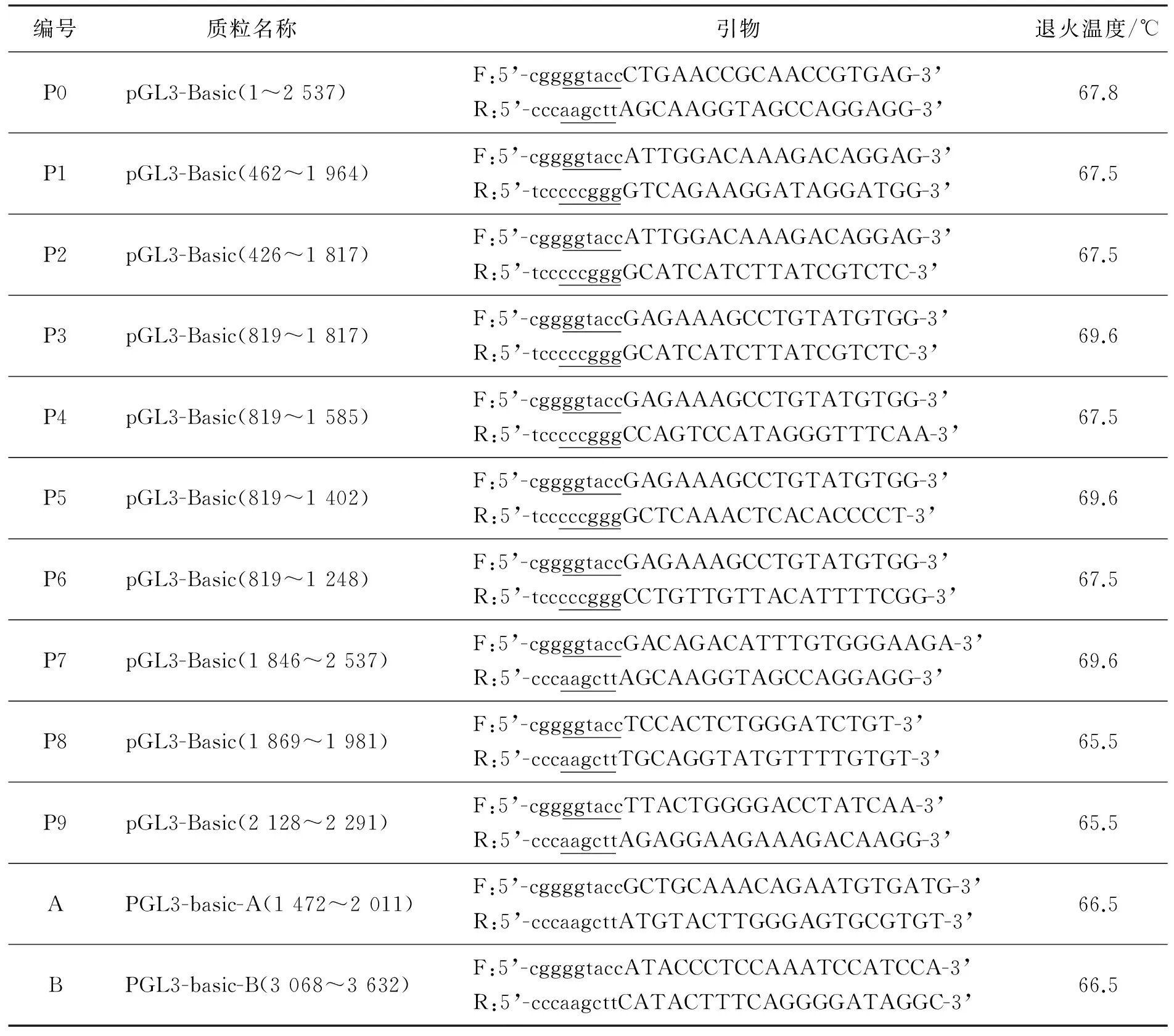
表1 扩增山羊Agouti 基因启动子预测区序列引物
注:小写字母为引入的保护碱基,下划线处为限制性酶切位点;大写字母为引物特异性序列。
1.3 目的片段的克隆及鉴定
连接pMD-19T载体体系:0.5 μL PMD-19T vector,0.5~2.0 μL PCR回收产物,2.5 μL Slution I ,灭菌的去离子水补齐到5 μL。然后离心混匀,16 ℃连接过夜然后用TransⅠ-T1感受态转化。过夜37 ℃摇床培养,利用通用引物,吸取1 μL菌液为模板做菌液PCR鉴定,选取阳性克隆,送华大基因公司测序鉴定。
1.4 重组质粒构建及鉴定
以回收的目的片段和酶切后的pGL3-Basic载体10∶1比例连接,用T4 DNA连接酶将目的片段连接到pGL3-basic载体上,构建含荧光素酶报告基因的重组质粒。菌液PCR鉴定后选阳性克隆,双酶切鉴定后华大基因进行测序鉴定。
1.5 细胞培养、转染及检测
在24孔培养板用胎牛血清体积分数为0.10的1640培养基培养293T细胞和A375细胞。培养条件为: CO2的体积分数为0.05,37 ℃温箱培养,24 h后当细胞汇合度达80%~90%且细胞生长状态良好时, 再根据Lipofectamine 2000脂质体转染说明进行转染实验,最后轻轻晃动孔板使之混匀,放入CO2的体积分数为0.05,37 ℃温箱培养。转染6 h后,用相应的含血清的正常细胞培养液换液,继续培养48 h后收获细胞。用1×PBS洗涤转染细胞1~2次,加入报告基因细胞裂解液裂解细胞和荧光剂用于检测。

图1 山羊基因组DNA琼脂糖凝胶电泳图
2 结果与分析
2.1 山羊基因组DNA的提取结果
提取的DNA条带致密、整齐、清晰,并且无明显拖尾(图1),说明DNA完整性较好,不需要纯化,可稀释后直接做模板进行后续PCR扩增。
2.2 目的片段启动子区活性验证
2.2.1 目的片段序列比对及扩增结果 目的片段A和B与云南黑山羊Agouti基因部分序列(登录号为AJPT01136617.1)同源性分别达92%和94%,其他片段同源性100%。利用模山羊基因组,TransStart Taq酶及引物进行PCR扩增,质量浓度为10.0 g/L的琼脂糖凝胶电泳检测,在约750,2 500 bp处有明亮条带(图2)。

图2 山羊Agouti基因启动子预测区 PCR扩增结果
2.2.2 重组质粒pMD19-T/2537/A/B的PCR鉴定与双酶切鉴定 利用重组质粒pMD19-T/2537和pMD19-T-A以及pMD19-T-B为模板,NS0和NA0为引物进行PCR扩增。质量浓度为10.0 g/L的琼脂糖凝胶电泳检测,在约2 500,750 bp处有明亮条带(图3)。
利用限制性内切酶HindⅢ和KpnⅠ对重组质粒pMD19-T/2537和pMD19-T-A,pMD19-T-B进行双酶切,质量浓度为10.0 g/L的琼脂糖凝胶电泳检测(图4)。
2.2.3 重组质粒pGL3-Basic/px/A/B的PCR与双酶切鉴定 利用重组质粒pGL3-Basic/px/A/B为模板及相应引物进行PCR扩增,质量浓度为10.0 g/L的琼脂糖凝胶电泳检测,电泳结果图谱为图5,双酶切电泳结果见图6。
2.2.4 测序鉴定 将上述经PCR和酶切鉴定正确的重组质粒pGL3-Basic/px/A/B送北京华大基因有限公司进行测序。结果表明,克隆到的目的片段与pGL3-Basic/px中所得山羊Agouti基因序列相似性高达90%以上,表明已成功构建了12个包含山羊Agouti基因重组质粒。
2.2.5 细胞转染及目的片段启动子活性鉴定 对A375细胞和293T细胞分别进行绿色荧光蛋白转染,结果表明,在m(脂质体)∶m(质粒总量)为1∶1时,转染效率最高,约为85%(图7,图8),不同转染比例的结果见表2。

图3 重组质粒pMD19-T-2537和pMD-T-A,pMD-T-B的PCR产物电泳图

图4 质粒pMD19-T/2537和pMD-T-A,pMD-T-B双酶切电泳图

图5 重组质粒pGL3-Basic/px/A/B PCR产物电泳图

图6 重组质粒pGL3-Basic/px/A/B双酶切电泳图

图7 A375细胞转染绿色荧光蛋白 图8 293T细胞转染绿色荧光蛋白
根据试验设计,在m(脂质体)∶m(质粒总量)为1∶1时,进行双荧光素酶报告基因两种质粒最佳比例的探索,结果表明,A375细胞转染pGL-3 Control和pRL-TK最佳比例为100∶1,293T细胞的最佳比例为800∶1(图9)。
根据所探索的最佳转染效率结果,转染体系见表3。
报告基因载体瞬时转染A375细胞及293T细胞。48 h后收获细胞并裂解细胞,计算相对Luc活性值。综合多次实验结果,用SPSS 19.0软件进行数据的统计分析,A375细胞和293T细胞阳性对照Luc值远高于阴性对照,说明细胞转染的效果比较好,阴性对照与pGL3-Basic/px/A/B的Luc值相比差异不显著(p>0.05),说明pGL3-Basic/px/A/B均不具备启动子活性(表4)。
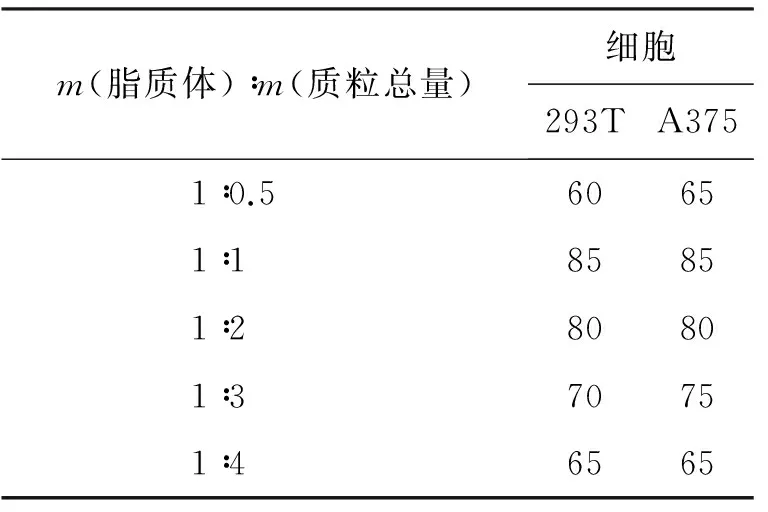
表2 不同lip和GFP质粒比例对细胞转染效率的影响 %

图9 不同pGL-3 Control和pRL-TK比例的Luc相对活性值

表3 A375和293T细胞的转染体系

表4 A375和293T细胞中片段pGL3-Basic/px/A/B的Luc活性比值
3 讨 论
实验室前期所测山羊Agouti基因部分序列(EF587236.2)与(JN651404)拼接序列长度为12 847 bp,预测出山羊Agouti基因候选启动子区为6 450~7 100 bp。将绵羊CH243-455O4的反向互补序列与12 847 bp(EF587236+JN651404)的拼接序列进行比对分析,获得12 847 bp序列之前的部分序列,拼接后得到长度为27 793 bp的山羊Agouti基因部分基因组序列。比对发现,这条长达27 793 bp的基因序列与NCBI中部分山羊Agouti基因片段(登录号为AJPT01136619.1,AJPT011366120.1,AJPT01136621.1,AJPT01136622.1,AJPT01136623.1)的部分序列都能比对上,并可以形成连续的片段。通过本次实验中所获得的片段,将山羊基因组片段(登录号为AJPT01136617.1,AJPT011366118.1,AJPT011366119.1)的所有序列拼接起来,直到与前期获得的27 793 bp片段有重合部分,这样就获得与27 793 bp拼接序列共36 067 bp,再加上之前的27 793 bp共63 860 bp,这样从所得的63 860 bp的序列来看,前期选取的候选启动子区域范围为56 180~58 717 bp,这2 537 bp的候选区域包括了Agouti基因外显子2的前44 bp。根据Michael等[11]对牛的预测启动子片段promoter A和promoter B与山羊基因组序列(登录号为AJPT01136617.1)比对,结果:promoter A与山羊基因组序列(登录号为AJPT01136617.1)1 472~2 011 bp位置相对应。Promoter B与山羊基因组序列(登录号为AJPT01136617.1)3 068~3 632 bp位置相对应,但是promoter B的前100 bp没有比对上。通过实验构建所获得的目的片段重组质粒转染细胞验证结果为:所获得目的片段均无启动子活性。从本次实验所选取的区域与实验室前期所选取的区域中间有16 481 bp。经过试验探索,接下来的工作可以从以下两方面进行:在这16 481 bp范围内,以2 000 bp为一个片段,可以逐段预测,根据预测结果并进行blast比对,来寻找一段保守性强,并且预测值高的片段进行启动子区寻找;根据前期课题组对5’非翻译区外显子1剪接体类型的研究,根据各种类型剪接体在基因组的相对位置,找到最靠近5’端剪接体之前的序列,预测启动子活性并进行试验验证;基于Norris等[12]对白色绵羊的ITCH启动子控制ASIP的表达的研究来设计引物,对其试验验证。
[1] 朱春凤,冯永山.Agouti基因及其表达产物的结构与功能[J].黑龙江医药,2012,25(3):401-402.
[2] 舒鸣,肖兵南,陈铁桥.牛毛色遗传的研究进展[J].中国牛业科学,2008,34(1):15-17,20.
[3] 杨云青,高丹玫,郭宗圣.Agouti和Agouti相关蛋白的一些生物学内涵[J].动物学杂志,2008,43(5):144-152.
[4] 朱智明,费宇行,石湘芸.Agouti基因及其相关基因与肥胖[J].天津医药,2000,28(11):702-704.
[5] 范瑞文,董常生,赫晓燕,等.哺乳动物毛色色素Agouti基因位点的研究进展[J].动物医学进展,2004,25(3):59-61.
[6] 唐春娟.山羊Agouti基因与毛色表型相关分析及其遗传变异的研究[D].保定:河北农业大学,2008.
[7] 赵静雯.山羊Agouti基因遗传多样性研究[D].保定:河北农业大学,2006.
[8] Li X L,Zhao J W,Tang C J,et al.Sequencing of part of the goatagoutigene and SNP identification[J].Biochemical genetics,2010,48(1-2):152-156.
[9] 张丽英.不同毛色山羊皮肤组织Agouti基因mRNA差异表达和5’UTR变异对表达影响[D].保定:河北农业大学,2013.[10] 吴伟伟.山羊Agouti基因5’UTR变异及ASIP蛋白表达定位研究[D].保定:河北农业大学,2012.
[11] Girardot M,Martin J,Guibert S.Widespread expression of the bovineAgoutigene results from at least three alternative promoters[J].Pigment Cell Res,2005,18:34-41.
[12] Norris B J,Whan V A.A gene duplication affecting expression of the ovine ASIP gene is responsible for white and black sheep[J]. Genome Res,2008,18(8):1 282-1 293.
Explore of Promoter Activity Area of GoatAgoutiGene
ZHANG Yong-gai1,2,3,GUO Min1,ZHANG Da-shan1,LIU Wei-lan2, LI Xiang-long1,ZHOU Rong-yan2,LI Lan-hui2
(1 Hebei Normal University of Science & Technology,Qinhuangdao Hebei,066004;2 Agricultural University of Hebei;3 Dingzhou Second Middle School;China)
The candidate promoter region (6 450-7 100 bp) of goatAgoutigene was predicted using the sequence joined from JN651404 and EF587236.2. The fragment of 2 537 bp was obtained, in which a typical transcription factor binding sites of TATA box was found, and other transcription factor binding sites (HSF, SYR, GATA-1, ADR1 and Nkx-2) were also predicted. Twelve luciferase reporter gene plasmids were constructed based on two parts of goat genome sequence of AJPT01136617.1 which had higher homology with cattle predicted promoters A and B. Then luciferase reporter gene plasmids were transiently transfected into cells of A375 and 293T to determine whether the fragments hadAgoutigene promoter activity. The results showed that the constructed plasmids had no significant difference compared with negative control, indicating the fragments had no promoter activity.
goat;Agoutigene;promoter
国家自然科学基金资助项目(项目编号:31172196);河北省应用基础研究计划重点基础研究资助项目(项目编号:15962901D)。
,男,博士,教授,博士研究生导师。主要研究方向:动物遗传育种。E-mail:lixianglongcn@yahoo.com。
2015-09-15
10.3969/J.ISSN.1672-7983.2015.03.001
S827.2
A
1672-7983(2015)03-0001-07
张永改(1987-),女,硕士。主要研究方向:动物遗传学。
(责任编辑:朱宝昌)
