三疣梭子蟹ftz-f1基因的克隆及相关核受体基因在蜕皮中的功能分析*
2015-04-10张龙涛吕建建高保全
张龙涛 吕建建 高保全 刘 萍① 付 萍
(1.中国水产科学研究院黄海水产研究所 青岛 266071;2.上海海洋大学水产与生命学院 上海 201306;3.青岛海洋科学与技术国家实验室海洋渔业科学与食物产出过程功能实验室 青岛 266235)
核受体(nuclear receptor)是多细胞生物中含量最丰富的几大类转录因子超家族之一,机体的生长发育、细胞分化以及许多生理、代谢过程都可归于核受体与相应配体及其它调节因子的相互作用(Boulanger et al,2011)。在哺乳类中,ftz-f1可归于核受体第5家族的甾类激素受体,这一家族也包括重要的甾类(类固醇)合成调控因子SF-1。ftz-f1首先是作为转录因子激活果蝇(Drosophila)早期胚胎形成阶段分化基因fushi tarazu的同源异性盒(homeobox)而被发现的(Kuroiwa et al,1984)。随后在其它许多物种中都发现了其同源类似物,如小鼠(Mus musculus)、家蚕(Bombyx mori)、烟草天蛾(Manduca sexta)和半滑舌鳎(Cynoglossus semilaevis)等(Chai et al,2000;Choi et al,2005;Cao et al,2012;Shafi et al,2013)。它们在DNA结合结构域和ftz-f1 box区域都有相对的高度保守性,且在其它区域保守性较弱。蜕皮激素(20E)与核受体EcR以及核受体家族的类视黄酸受体 RXR形成异二聚体复合物(配体-受体)复合物是引发 20E初级应答基因表达的关键环节(Riddiford et al,2000)。20E-EcR-USP与共激活因子TAIMAN复合物结合于启动子后,启动早期的核受体应答基因的表达,如fzt-f1、HR3等(Crossgrove et al,2008)。通过这些早期应答的核受体基因进一步调控下游基因的表达,如天冬氨酸凋亡酶编码基因、死亡激活因子等,导致蜕皮和变态过程中的细胞自噬和凋亡,从而调控蜕皮、变态等生理过程(Liu et al,2009;Spindler et al,2009)。此外,也有研究证明 ftz-f1基因在调控表皮基因的表达以及表皮蛋白的沉积上具有不可替代的作用(Yamada et al,2000)。然而有关ftz-f1基因在甲壳动物蜕皮中的功能研究尚无报道。
通过对本实验室三疣梭子蟹(Portunus trituberculatus)转录组数据库的筛选,获得一个在三疣梭子蟹不同蜕皮时期显著差异表达的核受体超家族基因ftz-f1的EST序列,克隆出该基因的全长cDNA序列并命名为PTNHr。采用生物信息学软件分析该基因及其翻译的氨基酸序列;借助Blast程序对与其相近物种的ftz-f1氨基酸序列同源性进行建树分析。采用实时荧光定量技术对核受体基因ftz-f1、EcR和RXR在蜕皮周期中的表达情况进行分析,对 ftz-f1基因在去除单侧眼柄后的表达情况进行分析,初步的阐明这些基因与蜕皮激素在蜕皮过程中的相互联系。本研究结果将有助于补充ftz-f1、EcR和RXR基因在三疣梭子蟹中与蜕皮激素相互作用与调控关系,进一步的丰富三疣梭子蟹蜕皮过程分子调控机制的研究。
1 材料与方法
1.1 实验动物
实验所用三疣梭子蟹取自黄海水产研究所实验基地日照开航水产养殖有限公司,体重为(32±5)g。暂养于室内圆柱形水泥池内,水池底面积为20m2,高度1.5m,池水深保持为20—30cm,水温25°C左右,pH 8.7,溶解氧5.5mg/L。暂养3d,每日8: 00定时换掉1/2的水,16: 00投喂蛤类或者野杂鱼(投喂量为蟹重的1/10)。
1.2 总RNA的提取及cDNA第一链的合成
采用 Trizol法分别提取三疣梭子蟹各个组织的总RNA,用1.0%的琼脂糖凝胶电泳检验总RNA的质量和完整性。取等量的眼柄、肝胰腺和肌肉组织的总 RNA混匀,严格按照SMARTTMRACE Amplification Kit试剂盒的说明方法来合成3′和5′RACE的cDNA第一链。
1.3 转录组测序
选取不同蜕皮时期三疣梭子蟹眼柄组织 RNA。构建cDNA文库,送交诺禾致源公司的Illumia Hiseq 2000平台进行测序。从29889个unigene中总共筛选出 1389个蜕皮相关的功能基因,挑选出三疣梭子蟹ftz-f1基因序列做进一步的研究。
1.4 引物的设计
根据得到的三疣梭子蟹ftz-f1基因的EST序列,利用Primer Premier 5.0软件基于RACE原理设计3′和5′末端特异性引物。根据 NCBI上已公布的三疣梭子蟹蜕皮激素受体基因(EcR JQ250795)和类视黄酸受体基因(RXR KF061043.1)设计荧光定量引物。实验中所用到的引物由上海生工生物有限公司合成(表1)。
1.5 三疣梭子蟹ftz-f1基因全长cDNA的克隆及验证
利用Advantage 2 PCR Kit试剂进行末端扩增,3′用通用引物 UPM和 NUP分别与相应的特异性引物NHr-F1和NHr-F2进行巢式PCR扩增(反应程序94°C 30s,68°C 30s,72°C 3min,27 个循环);同样的程序以及相应的特异性引物进行5′端的扩增。
将 3′和 5′RACE 的扩增产物经琼脂糖凝胶电泳,切胶纯化并且连接到PMD18-T载体,转化入大肠杆菌Top ten感受态细胞中,在培养基中37°C过夜培养后,挑取阳性菌落继续培养。进行菌落PCR后,挑选含有目的条带的菌液送交上海桑尼公司测序。在该基因的两端设计正反向引物,进行全长cDNA的验证。

表1 实验中所用到的引物Tab.1 The Sequences of the primer used in this experiment
1.6 三疣梭子蟹ftz-f1基因的生物信息学分析
利用NCBI中的BLAST主页程序进行主要的核苷酸序列以及氨基酸序列的比对。利用 Vector NTI 11.0 进行核苷酸序列的拼接比对、冗余序列的去除、开放阅读框的预测和氨基酸的翻译。利用DNAMAN进行氨基酸序列的比对。利用SMRAT和InterProScan进行蛋白质跨膜区的分析。利用ProParam tool进行氨基酸残基分析及蛋白质等电点、分子量等预测。利用MEGA 6.0 构建系统进化树。
1.7 蜕皮过程的分期及去除单侧眼柄实验
采用解剖镜观察三疣梭子蟹游泳足表皮生长状况的方法来区分不同蜕皮时期(沈洁等,2011)。取处于蜕皮间期、蜕皮前期、蜕皮后期的蟹各6只,分别取其眼柄、肌肉、肝胰腺、心脏、血液、鳃,每两只螃蟹的组织放入一个 EP管中,存放于液氮中保存,用以之后的提取RNA和定量分析使用。
采用烫伤法去除三疣梭子蟹的单侧眼柄。将烧红的镊子夹住蟹眼后红色膨大部分基部(此处为眼柄窦宪复合体)1—2s,破坏单侧眼柄的分泌及调控功能。用土霉素按照50—100mg/L的浓度进行全池的消毒。暂养7d后挑取处于蜕皮间期、前期、后期的蟹,取眼柄、肌肉、肝胰腺、心脏、血液、鳃组织,存放于液氮中保存。
1.8 三疣梭子蟹 ftz-f1、EcR和 RXR基因的表达定量分析
用研钵将采集的组织样品在液氮中研磨,每个 EP管单独研磨,然后在每 1mL Trizol试剂中加入 50—100mg的组织粉末,震荡混匀分开标记后放入–80°C中保存。按照Trizol法提取RNA的步骤提取各个组织的总RNA,利用核酸定量仪和琼脂糖凝胶电泳检测RNA的质量和完整性,使用 Transcriptor First Strand cDNA Synthesis Kit (ROX)试剂盒将RNA反转录为cDNA。
采用 FastStart Universal SYBR Green Master试剂,在 ABI 7500 Real Time PCR仪对各个组织的基因表达情况进行测定。反应体系为 10μL,包括 5μL的FastStart Universal SYBR Green Master (ROX)、0.1μL的正反向引物、3.8μL的PCR反应水、1μL的cDNA。反应程序为 95°C 10min,95°C 10s,60°C 34s,40 个循环;95°C 15s,60°C 1min,95°C 15s。为了得到更为可靠的定量分析结果,采用 geNorm软件对β-actin和18S rRNA这两个内参基因进行分析,结果显示β-actin在蜕皮周期更为稳定。以β-actin基因作为内参,样本和内参均设置3个重复,每个样本为单个EP管中组织合成的模版。采用2–△△Ct方法计算各个基因的相对表达量,使用SPSS 19.0统计软件进行单因素方差分析其显著性,P<0.05认为差异显著。
2 结果
2.1 总RNA提取
采用Trizol法提取的各个组织的RNA,经 1.0%琼脂糖凝胶电泳检测后,呈现3条清晰的条带,说明RNA完整性较好;用核酸定量仪检测 RNA,其OD260/OD280均在1.9—2.0之间,说明RNA纯度较高,可用于后续实验。
2.2 三疣梭子蟹ftz-f1基因全长cDNA的克隆及验证
该基因的EST序列长1165bp,用blastx对序列进行在线比对,结果显示: 该序列与其它物种的ftz-f1的相似度在65%以上。3′RACE扩增结果得到大小为666bp的cDNA片段,5′RACE扩增结果得到大小为539bp的cDNA片段。将这两个片段与已知EST序列进行拼接,去除冗余序列得到三疣梭子蟹 ftz-f1基因的全长cDNA序列,命名为PTNHr。对该基因全长 cDNA进行测序验证,验证出覆盖开放阅读框的1585bp序列准确、可靠。该cDNA序列全长为1763bp(GenBank登录号KP299258),其中包括588bp的开放阅读框(ORF),141bp的5’端非编码区(UTR),1034bp的3’端非编码区(UTR)(图1)。
2.3 三疣梭子蟹ftz-f1基因序列的生物学分析
DNAstar软件分析表明,三疣梭子蟹 ftz-f1基因编码一个由 195个氨基酸组成的蛋白质,分子量为22.8kDa,理论等电点为 6.35。该蛋白具有核受体家族标志的 DNA 结合域(DBD)和配体结合域(LBD)。ProtParam tool软件分析表明,ftz-f1蛋白带有负电的氨基酸残基为26个(Asp和Glu),带有正电的氨基酸残基为 23个(Arg和 Lys),不稳定系数为27.7,亲水性平均数为–0.291,属于稳定蛋白。使用 SMRAT和InterProScan在线软件分析,ftz-f1基因翻译的蛋白质结构中无跨膜结构域。
2.4 三疣梭子蟹ftz-f1基因的同源性分析
BLAST同源性分析三疣梭子蟹ftz-f1基因的氨基酸序列,发现与刀额新对虾(Metapenaeus ensis)同源性达到75.4%,与切叶蚁(Acromyrmex echinatior)、德国小蠊(Blattella germanica)、黑腹果蝇(Drosophila melanogaster)、烟草天蛾(Manduca sexta)、赤拟谷盗(Tribolium castaneum)的同源性分别为66.0%、67.5%、61.9%、61.9%和 66.0%(图2)。

图1 三疣梭子蟹ftz-f1基因cDNA全长及其编码的氨基酸序列Fig.1 ftz-f1 nucleotide sequence and deduced amino acid sequence of P.trituberculatus

图2 三疣梭子蟹ftz-f1氨基酸序列与其它物种的ftz-f1氨基酸序列比对Fig.2 The amino acid sequence of P.trituberculatus ftz-f1 alignment with other species
利用 MEGA6.0软件进行系统进化分析,构建系统进化树(图3)。结果显示13个物种中,三疣梭子蟹与刀额新对虾紧密聚为一支,之后与赤拟谷盗、德国小蠊亲缘关系较近。在昆虫中,黑腹果蝇与嗜凤梨果蝇紧密聚为一支,家蚕与烟草天蛾等聚为一支。
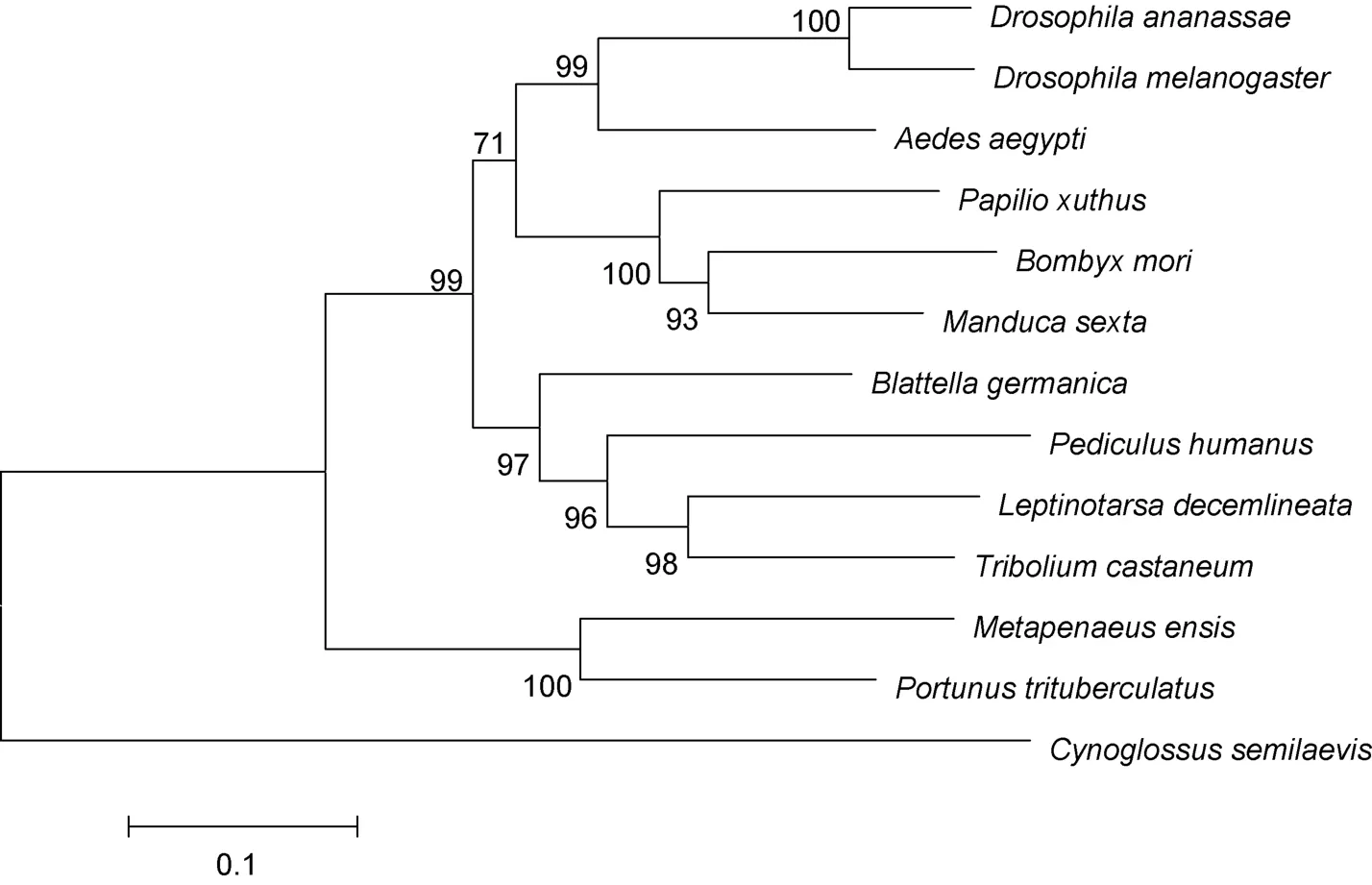
图3 MEGA 6.0软件采用邻接法构建的三疣梭子蟹与其它物种的ftz-f1氨基酸的系统进化树Fig.3 NJ phylogenetic tree of amino acid sequence of P.trituberculatus ftz-f1 and other species made by MEGA 6.0
2.5 三疣梭子蟹三个核受体基因的表达情况分析
2.5.1 三疣梭子蟹不同蜕皮时期,各个组织中ftz-f1、EcR和RXR基因的表达情况 采用实时荧光定量PCR方法,分析三疣梭子蟹各个组织中ftz-f1基因在不同的蜕皮时期的表达特征。结果表明,眼柄中ftz-f1基因在蜕皮后期的表达量明显高于蜕皮间期和蜕皮前期;但是肝胰腺、心脏和鳃中ftz-f1基因在蜕皮后期却明显低于蜕皮间期和蜕皮前期;血细胞和肌肉中该基因的表达量在蜕皮各个时期的变化不明显(图4)。
三疣梭子蟹各个组织中EcR基因在不同的蜕皮时期的表达情况: 在眼柄、心脏和鳃中,三疣梭子蟹 EcR基因的表达量均为蜕皮前期高于蜕皮间期和蜕皮后期;在血细胞、肌肉和肝胰腺中,EcR基因的表达量为蜕皮后期高于蜕皮间期和蜕皮前期(图5)。
三疣梭子蟹各个组织中 RXR基因在不同的蜕皮时期的表达情况: 眼柄中 RXR基因在蜕皮后期表达量低于蜕皮间期和蜕皮前期;血细胞、肝胰腺、心脏和鳃中 RXR基因在蜕皮后期表达量明显高于蜕皮间期和蜕皮前期;肌肉中 RXR基因表达量在蜕皮各个时期中变化不明显(图6)。
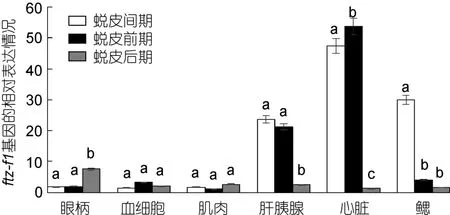
图4 三疣梭子蟹ftz-f1基因在不同蜕皮时期各个组织中的表达分布情况Fig.4 Distribution of ftz-f1 gene expression in different tissues in different molt stages
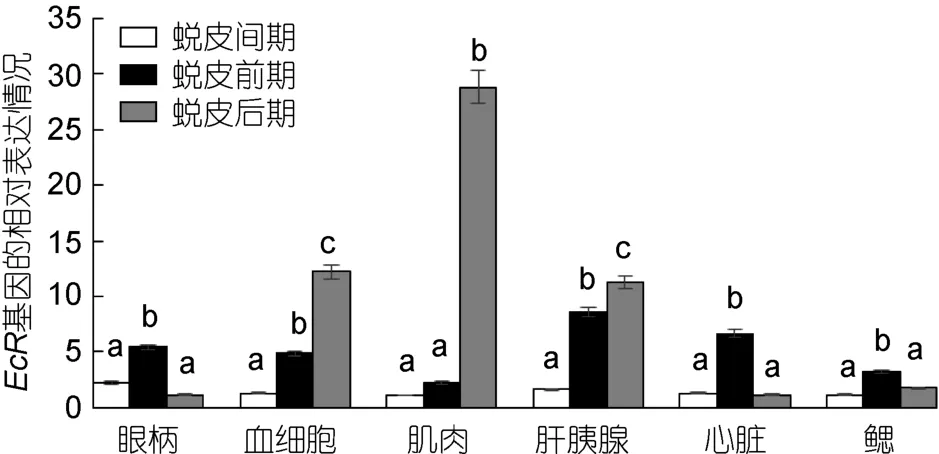
图5 三疣梭子蟹EcR基因在不同蜕皮时期各个组织中的表达分布情况Fig.5 Distribution of EcR gene expression in different tissues in different molt stages
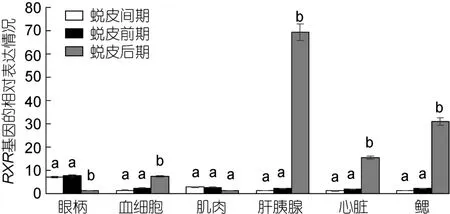
图6 三疣梭子蟹RXR基因在不同蜕皮时期各个组织中的表达分布情况Fig.6 Distribution of RXR gene expression in different tissues in different molt stages
2.5.2 三疣梭子蟹 ftz-f1基因在不同蜕皮时期,去除单侧眼柄与正常相对比的表达情况 对去除单侧眼柄的三疣梭子蟹在不同蜕皮时期,与相应的正常对照组进行实时荧光定量。结果显示: 在蜕皮前期与蜕皮间期,去除单侧眼柄后 ftz-f1基因的表达量在整体上明显低于相应的正常对照组,并且表达差异较大(图7、图8)。但是在蜕皮后期,去除单侧眼柄后ftz-f1基因的表达量在肝胰腺、心脏和鳃中的表达量却是高于正常对照组(图9)。
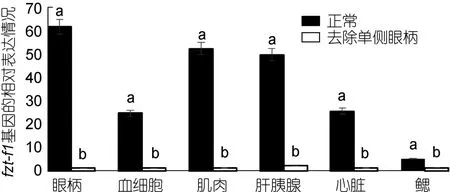
图7 去除单侧眼柄后,ftz-f1基因在三疣梭子蟹蜕皮前期与正常对照各个组织中的相对表达情况Fig.7 After single eye- ablation,distribution of ftz-f1 gene expression in different tissues of pre-molt stage compare to those in normal condition
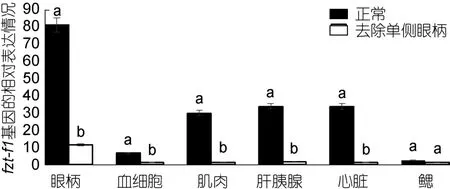
图8 去除单侧眼柄后,ftz-f1基因在三疣梭子蟹蜕皮间期与正常对照各个组织中的相对表达情况Fig.8 After single eye- ablation,distribution of ftz-f1 gene expression in different tissues in inter-molt stage compare to that in normal condition
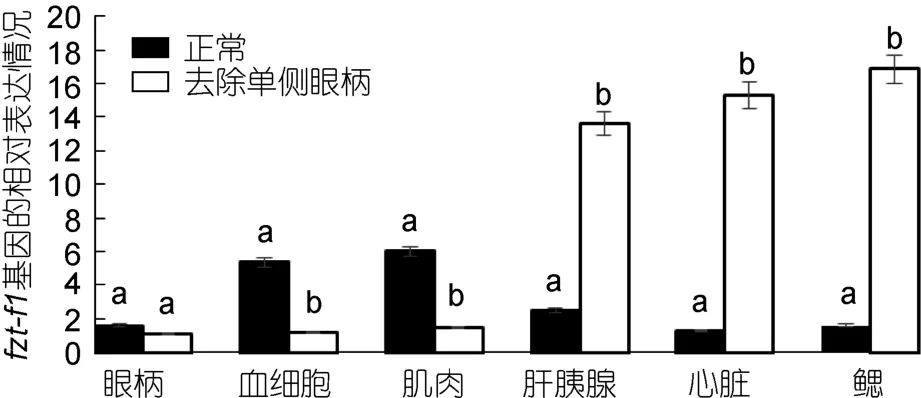
图9 去除单侧眼柄后,ftz-f1基因在三疣梭子蟹蜕皮后期与正常对照各个组织中的相对表达情况Fig.9 After single eye- ablation,distribution of ftz-f1 gene expression in different tissues of post-molt stage compare to normal condition
3 讨论
核受体是配体依赖性转录因子超家族,能够直接与 DNA结合,调节下游靶基因的表达,与机体的代谢、生殖和发育等多种生理过程密切相关(Mckenna et al,2002)。fzt-f1基因属于核受体第5家族A组,该组内的核受体基因是重要的甾类激素合成调控因子(Lu et al,2001)。fzt-f1基因在鱼类中研究较为深入,但是在甲壳类中却很少有报道(Liu et al,1997;Masuda et al,1998;Lin et al,2000)。本实验克隆得到三疣梭子蟹 fzt-f1基因,该基因具有核受体基因特有的DNA结合结构域(DBD)和配体结合区(LBD)。该基因序列与刀额新对虾克隆出的fzt-f1蛋白同源性达到75.4%,系统进化分析中也与刀额新对虾聚为一支。
蜕皮激素对于甲壳类的蜕皮过程起着至关重要的作用。已有的研究证明,蜕皮激素在血液中是以20-羟蜕皮酮(20E)形式存在。20E在血液内的浓度在蜕皮前期逐渐升高,临近蜕皮时,形成一个峰值,然后浓度再迅速下降(Wang et al,2000;Watson et al,2001)。整个调控过程是通过20E-EcR-RXR配体-受体复合物引发的应答基因级联放大反应完成,20E与蜕皮激素受体EcR和类视黄酸受体(RXR)异二聚体复合物在细胞核内形成转录复合体,进而引发20E初级应答基因(如 fzt-f1、HR3等)表达(Durica et al,1999;LeBlanc,2007)。初级反应基因编码的转录因子诱导表达的20E次级应答基因级联放大20E信号,进而调控下游基因表达。
实时定量分析ftz-f1基因在不同蜕皮时期各个组织中的相对表达情况,结果表明: 眼柄中 ftz-f1基因的表达量为蜕皮后期高于蜕皮前期和蜕皮间期,而在肝胰腺、心脏和鳃组织中ftz-f1基因的表达量为蜕皮后期低于蜕皮前期和蜕皮间期。推测在眼柄中,该基因与20E呈负调控关系,而在肝胰腺、心脏和鳃中该基因与20E呈正调控关系。由于眼柄中X器-窦腺复合体合成蜕皮抑制激素(MIH)(Nakatsuji et al,2004),因此该基因在眼柄中的表达可能与 MIH更为相关。对核受体基因EcR在蜕皮周期中的定量分析结果显示: 眼柄、心脏和鳃组织中,EcR基因在蜕皮前期表达量高于其它两蜕皮时期,表达模式与20E在蜕皮时期的浓度变化相似,说明了该基因与20E的协同调节作用,这与日本囊对虾(Marsupenaeus japonicus)的研究结果一致(Asazuma et al,2007)。在血细胞、肝胰腺和肌肉中,EcR基因的表达模式为蜕皮后期高于蜕皮前期高于蜕皮间期,出现了与20E浓度不协调的现象。也有学者发现,在蓝蟹(Callinectes sapidus)体内 EcR基因的表达量与20E在血淋巴中的浓度并不协同,关于这一现象的解释还有待于进一步的研究(Techa et al,2013)。核受体基因RXR在蜕皮周期的定量结果显示: RXR与ftz-f1基因在肝胰腺、心脏和鳃组织中的表达模式上呈现出完全相反的趋势,这预示着RXR和fzt-f1这两个基因之间可能存在典型的抑制调控作用。而RXR与EcR基因在整体上呈现相似的表达模式,这与中国对虾(Fenneropenaeus chinensis)和中华绒螯蟹(Eriocheir sinensis)中的研究相一致(Priya et al,2009;王瑶等,2013)。由于RXR蛋白和EcR蛋白是作为20E异二聚体复合物的组成部分,因此这两个基因在三疣梭子蟹的蜕皮功能上也更趋向于协同调节作用。
甲壳类蜕皮激素是由 Y器官合成与分泌,而 Y器官则受 MIH的调控(罗荣生等,1990;Gong et al,2015)。眼柄中 X器-窦腺复合体是 MIH制造与分泌的器官,去除单侧眼柄将会造成 20E浓度的上升(Devaraj et al,2006;Wang et al,2013)。去除单侧眼柄后对 fzt-f1基因在各个组织中的表达量进行分析,发现在去除单侧眼柄后该基因的表达量在各个蜕皮时期组织中出现明显下降;说明 20E浓度的升高对ftz-f1基因的表达起到了明显的抑制作用。由此可以推测出,ftz-f1基因在蜕皮过程中具有重要的作用,与20E具有紧密的联系,且更可能为抑制调控关系。
4 结论
本研究成功克隆出了三疣梭子蟹ftz-f1基因全长cDNA序列,并进行了生物信息学的分析。对三疣梭子蟹 ftz-f1基因以及相关的核受体基因EcR和 RXR,在蜕皮周期各个组织中的表达情况进行了分析并预测之间可能存在的联系。本实验证明了ftz-f1基因在蜕皮过程中的调控作用,并对与三个重要的蜕皮功能相关核受体基因之间的联系进行了初步的探讨。有关 ftz-f1基因以及核受体基因在蜕皮中的功能,还需要更多的研究。关于核受体基因之间的相互作用与联系也需要更为深入的生物学技术来进行研究。
王 瑶,杨志刚,郭子好等,2013.中华绒螯蟹RXR基因全长cDNA克隆及表达分析.水产学报,37(12): 1761— 1769
沈 洁,朱冬发,胡则辉等,2011.三疣梭子蟹蜕皮周期的分期.水产学报,35(10): 1481—1487
罗荣生,王幽兰,曹梅讯等,1990.中华绒螯蟹血淋巴20-羟蜕皮酮诱发蜕皮和卵巢发育的作用.动物学报,36(2):157—164
Asazuma H,Nagata S,Kono M et al,2007.Molecular cloning and expression analysis of ecdysone receptor and retinoid X receptor from the kuruma prawn,Marsupenaeus japonicus.Comp Biochem Physiol B Biochem Mol Biol,148(2):139—150
Boulanger A,Clouet-Redt C,Farge M et al,2011.Ftz-f1 and Hr39 opposing roles on EcR expression during Drosophila mushroom body neuron remodeling.Nat Neurosci,14(1):37—44
Cao J L,Chen J J,Jiang Z L et al,2012.Molecular cloning and expression analysis of FTZ-F1 in the GIFT tilapia,Oreochromis niloticus.Zool Res,33(E3-4): E40—E46
Chai C,Chan W K,2000.Developmental expression of a novel Ftz-F1 homologue,ff1b (NR5A4),in the zebrafish Danio rerio.Mech Dev,91(1—2): 421—426
Gong J,Ye H H,Xie Y J et al,2015.Ecdysone receptor in the mud crab Scylla paramamosain: a possible role in promoting ovarian development.J Endocrinol,224(3): 273—287
Choi C Y,Habibi H R,2005.Molecular cloning and tissue distribution of SF-1-related orphan receptors during sexual maturation in female goldfish.Biotechnol Lett,27(17):1283—1290
Crossgrove K,Maina C V,Robinson-Rechavic M et al,2008.Orthologues of the Drosophila melanogaster E75 molting control gene in the filarial parasites Brugia malayi and Dirofilaria immitis.Mol Biochem Parasitol,157(1): 92—97
Durica D S,Chung A C K,Hopkings P M et al,1999.Characterization of EcR and RXR gene homologs and receptor expression during the molt cycle in the crab,Uca pugilator.Amer Zool,39(4): 758—773
Devaraj H,Natarajan A,2006.Molecular mechanisms regulating molting in a crustacean.FEBS J,273(4): 839—846
LeBlanc G A,2007.Crustacean endocrine toxicology: a review.Ecotoxicology,16(1): 61—81
Kuroiwa A,Hafen E,Gehring W J,1984.Cloning and transcriptional analysis of the segmentation gene fushi tarazu of Drosophila.Cell,37(3): 825—831
Wang C J,Zhu D F,Qi Y Z et al,2013.Molt-inhibiting hormone levels and ecdysteroid titer during a molt cycle of Portunus trituberculatus.Acta Hydrobiol Sin,37(1): 22—28
Lin W W,Wang H W,Sum C et al,2000.Zebrafish ftz-f1 gene has two promoters,is alternatively spliced,and is expressed in digestive organs.Biochem J,348(2): 439—446
Liu D,Le Drean Y,Ekker M et al,1997.Teleost FTZ-F1 homolog and its splicing variant determine the expression of the salmon gonadotropin IIbeta subunit gene.Mol Endocrinol,11(7): 877—890
Liu Y,Sheng Z T,Liu H H et al,2009.Juvenile hormone counteracts the bHLH-PAS transcription factors MET and GCE to prevent caspase-dependent programmed cell death in Drosophila.Development,136(12): 2015—2025
Lu T T,Repa J J,Mangelsdorf D J,2001.Orphan nuclear receptors as eLiXiRs and FiXeRs of sterol metabolism.J Biol Chem,276(41): 37735—37738
Ito M,Masuda A,Yumoto K et al,1998.cDNA cloning of a new member of the FTZ-F1 subfamily from a rainbow trout.Biochim Biophys Acta,1395(3): 271—274
McKenna N J,O'Malley B W,2002.Combinatorial control of gene expression by nuclear receptors and coregulators.Cell,108(4): 465—474
Wang W L,Spaziani E,Huang H Z et al,2000.Ecdysteroid hormones and metabolites of the stone crab,Menippe mercenaria.J Exp Zool,286(7): 725—735
Nakatsuji T,Sonobe H,2004.Regulation of ecdysteroid secretion from the Y-organ by molt-inhibiting hormone in the American crayfish,Procambarus clarkii.Gen Comp Endocrinol,135(3): 358—364
Priya T A J,Li F H,Zhang J Q et al,2009.Molecular characterization and effect of RNA interference of retinoid X receptor (RXR)on E75 and chitinase gene expression in Chinese shrimp Fenneropenaeus chinensis.Comp Biochem Physiol B Biochem Mol Biol,153(1): 121—129
Riddiford L M,Cherbas P,Truman J W,2000.Ecdysone receptors and their biological actions.Vitam Horm,60(1):1—73
Shafi M,Wang Y N,Zhou X S et al,2013.Isolation and expression analysis of FTZ-F1 encoding gene of black rock fish (Sebastes schlegelii).J Ocean Univ China,12(1):183—189
Spindler K D,Hönl C,Tremmel C H et al,2009.Ecdysteroid hormone action.Cell Mol Life Sci,66(24): 3837—3850
Techa S,Chung J S,2013.Ecdysone and retinoid-X receptors of the blue crab,Callinectes sapidus: Cloning and their expression patterns in eyestalks and Y-organs during the molt cycle.Gene,527(1): 139—153
Watson R D,Lee K J,Qiu S H et al,2001.Molecular cloning,expression,and tissue distribution of crustacean moltinhibiting hormone.Am Zool,39(5): 407—417
Yamada M,Murata T,Hirose S et al,2000.Temporally restricted expression of transcription factor betaFTZ-F1: significance for embryogenesis,molting and metamorphosis in Drosophila melanogaster.Development,127(23): 5083—5092
