高原鼠兔干扰对达乌里秦艽 繁殖特征的影响
2015-04-08贾婷婷庞晓攀金少红陈传武郭正刚
王 倩,贾婷婷,庞晓攀,金少红,陈传武,郭正刚
(兰州大学草地农业科技学院草地农业生态系统国家重点实验室,甘肃 兰州730020)
达乌里秦艽(Gentiana dahurica)是青藏高原高山嵩草(Kobresia p yg maea)草甸的主要伴生种,也是我国大宗的中药材植物之一[1]。其种群密度变化必然引起高山嵩草草甸内其他植物成分的联动变化[2]。达乌里秦艽种群往往受人为樵采和环境适宜性的影响,其中樵采是可控制因素[3],而环境适宜性中的多种外界干扰因素人为控制相对困难[4]。若外界因素干扰后有利于达乌里秦艽繁殖,则会增加其适应性,若外界因素干扰不利于达乌里秦艽繁殖,则会降低其适应性。
高原鼠兔(Ochotona cur zoniae)是青藏高原高山嵩草草甸生态系统中的关键种[5],其通过啃食和挖掘等干扰行为改变着高山嵩草草甸的生境,从而影响植物种群的正常生长发育和生理特性[6-8]。庞晓攀等[2]的研究表明,高原鼠兔干扰明显改变了高山嵩草草甸内植物种群的空间分布格局,贾婷婷等[9]发现,高山嵩草草甸内不同植物的生态位宽度随着高原鼠兔干扰程度增加而呈现不同的变化趋势,说明高山嵩草草甸内不同植物种群对高原鼠兔干扰的响应存在一定的分异特征,有些植物种群遭受高原鼠兔干扰时会释放其生长潜势[8],而有些植物种群的生长潜势则会受到高原鼠兔干扰的抑制[6]。目前已经证实随高原鼠兔干扰水平增加,达乌里秦艽的高度、盖度和生物量均表现为增加的变化态势,这说明高原鼠兔干扰有利于达乌里秦艽的生长[2],但高原鼠兔干扰对达乌里秦艽植株的繁殖特征是否存在影响,需要科学研究提供证据。
繁殖器官是达乌里秦艽体现植物繁殖特征的重要指标[10]。研究达乌里秦艽繁殖器官对高原鼠兔干扰的响应,能够解析高原鼠兔干扰对达乌里秦艽种群繁殖特征的影响,从而在一定程度上能够揭示达乌里秦艽种群对高原鼠兔的响应及其适应变化后生境的能力。因此,本研究通过实地调查方法,分析不同高原鼠兔干扰水平下达乌里秦艽繁殖器官的变化特征,以阐明高山嵩草草甸伴生种达乌里秦艽繁殖特征对高原鼠兔干扰的响应,为揭示高原鼠兔干扰对达乌里秦艽生长的影响提供基础资料。
1 研究地区和方法
1.1 研究地区概况
研究地区位于青藏高原东部甘肃省玛曲县,地理坐标100°40′-102°29′E,33°06′-34°30′N。海拔3 300-4 806 m。气候为高寒湿润气候,无绝对无霜期。年均温度1.2 ℃,1月份平均温度为-10℃,7月份平均气温为11.7 ℃。年均降水量为564 mm,主要集中于5-9 月,年蒸发量1 000~1 500 mm。土壤为亚高山草甸土,表层有5~10 c m 草皮层,有机质含量高达10%~15%。植被类型以高寒草甸为主,优势种为高山嵩草,主要伴生种为达乌里秦艽,垂穗披碱草(El y mus nutans),小花草玉梅(Anemone rivul aris var.f lore-minore),矮 藨 草(Scir pus pu mil us),羊茅(Festuca ovina),米口袋(Guel denstaedtia ver na)等。
1.2 研究方法
1.2.1 试验设计 本研究采用高原鼠兔有效鼠洞口数密度代替其干扰强度的方法[11]研究高原鼠兔干扰对达乌里秦艽繁殖特征的影响。2012年5 月在玛曲县阿孜站的冷季型草地上随机选定了地势相对一致的36个25 m×25 m 样地[11-13],调查每个样地的有效洞口数、洞口裸斑面积和草丛高度,以这3个因子为变量将36个样地聚成4个类群,每个类群的有效洞口数密度分别为(160±48)、(240±32)、(336±48)、(496±64)个·h m-2,每个梯度选择3个样地作为重复,然后对样地进行固定监测。在植物生长季对样地进行围栏保护,非生长季11月到次年2月进行放牧,放牧强度一致。2014年8月重新调查每个样地的高原鼠兔有效洞口数密度,样地内的密度虽有变化,但梯度依旧明显,分别为(112±32)、(192±48)、(352±64)和(608±48)个·h m-2,分别标记为Ⅰ、Ⅱ、Ⅲ、Ⅳ4 个干扰水平。随着高原鼠兔干扰水平的增大,高山嵩草草甸植物群落的高度和盖度呈降低趋势[2]。取样时间为2014年8月,即有效洞口数密度调查完毕后即可取样。沿样地对角线每间隔7 m 处为圆点,半径为3 m 的圆形区为采样区,每个采样区在中心点附近选定达乌里秦艽1株,测定自然状态下的株高,然后采集。每个样地内共采集达乌里秦艽9株,每个干扰水平下采集达乌里秦艽27株,共采集108株。采集样品后,将每株达乌里秦艽进行茎叶分离,然后将枝条分为营养枝和生殖枝,查数每枝条上的花序数,若有成熟种子时,则收获种子,从而获取每株的营养枝数、生殖枝数、花序数和种子重量。然后将各部分分别装入信封中,置于60 ℃烘箱中烘至恒重,分别称量茎、叶、营养枝、生殖枝、花序和种子的干重。地上生物量即为各部分干重的总和。
1.2.2 数据分析 利用SPSS 16.0统计软件进行数据处理。用One-Way ANOVA 进行单因素方差分析,若有显著性差异,使用Duncan法对平均值进行多重比较。用Partial Correlation进行偏相关分析。用Excel 2010制图。
2 结果与分析
2.1 高原鼠兔干扰对达乌里秦艽株高和地上生物量的影响
高原鼠兔干扰明显影响了达乌里秦艽的株高和地上生物量(图1),表现为随着高原鼠兔干扰水平的增加,达乌里秦艽的株高和地上生物量均呈增加的态势,其中干扰水平为Ⅲ和Ⅳ时,其株高显著高于干扰水平为Ⅰ和Ⅱ时的株高(F(3,104)=6.551,P<0.05);地上生物量在干扰水平Ⅱ和Ⅲ间差异不显著,但干扰水平为Ⅱ和Ⅲ时的地上生物量显著高于干扰水平为Ⅰ时的地上生物量,且显著低于干扰水平为Ⅳ时的地上生物量(F(3,104)=8.680,P<0.05)。
2.2 高原鼠兔干扰对达乌里秦艽茎叶比的影响
高原鼠兔干扰水平变化对达乌里秦艽茎叶比具有明显影响。随高原鼠兔干扰水平的增加,达乌里秦艽的茎叶比表现为增加的变化趋势(图2),干扰水平为Ⅲ和Ⅳ时的茎叶比显著高于干扰水平为Ⅱ时的茎叶比,后者又显著大于干扰水平为Ⅰ时的茎叶比(F(3,104)=17.969,P<0.05)。
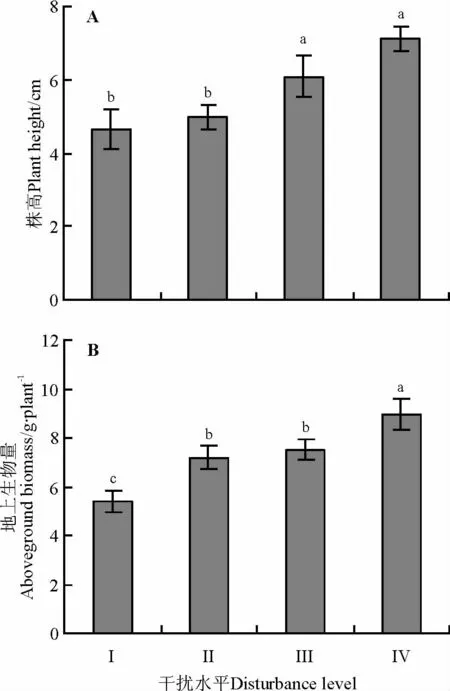
图1 高原鼠兔干扰对达乌里秦艽株高和地上生物量的影响Fig.1 Effects of plateau pika distur bance levels on plant height and aboveground biomass of Gentiana dahurica
2.3 高原鼠兔干扰对达乌里秦艽营养枝数和生殖枝数的影响
达乌里秦艽营养枝数和生殖枝数随着高原鼠兔干扰水平增加均表现为增加的态势(图3),干扰水平为Ⅲ和Ⅳ时的达乌里秦艽营养枝数和生殖枝数均显著大于干扰水平为Ⅰ和Ⅱ时的营养枝数和生殖枝数(F(3,104)=29.799,P<0.05;F(3,104)=9.653,P<0.05),但生殖枝数在干扰水平Ⅲ和Ⅳ间差异不显著,而干扰水平为Ⅳ的营养枝数显著大于干扰水平为Ⅲ时的营养枝数。
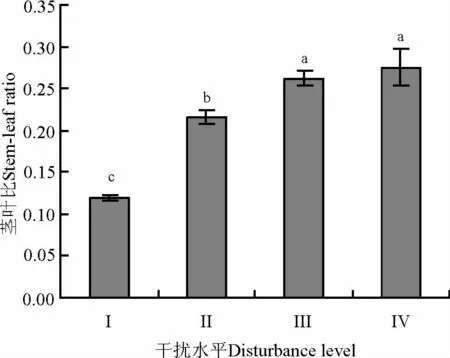
图2 高原鼠兔干扰对达乌里秦艽茎叶比的影响Fig.2 Effects of plateau pika disturbance levels on stem-leaf radio of Gentiana dahurica

图3 高原鼠兔干扰对达乌里秦艽营养枝数和生殖枝数的影响Fig.3 Effects of plateau pika distur bance levels on numbers of vegetative shoot and reproductive shoot of Gentiana dahurica
2.4 高原鼠兔干扰对达乌里秦艽花序数和花序生物量的影响
当高原鼠兔干扰水平从Ⅰ增加至Ⅳ的过程中,达乌里秦艽的花序数和花序生物量均呈现增加的趋势(图4)。干扰水平为Ⅳ时的花序数显著大于干扰水平为Ⅲ时的花序数,而后者又显著大于干扰水平为Ⅰ和Ⅱ时的花序数(F(3,104)=7.37,P<0.05),但干扰水平Ⅰ和Ⅱ间的花序数差异不显著。干扰水平为Ⅳ时的花序生物量显著高于干扰水平为Ⅱ和Ⅰ时的花序生物量(F(3,104)=9.131,P<0.05),但花序生物量在干扰水平Ⅱ和Ⅲ、Ⅲ和Ⅳ间差异不显著(P>0.05)。

图4 高原鼠兔干扰对达乌里秦艽花序数(A)和花序生物量(B)的影响Fig.4 Effects of plateau pika distur bance levels on inflorescence numbers(A)and inflorescence biomass(B)of Gentiana dahurica
2.5 高原鼠兔干扰对达乌里秦艽单株种子产量的影响
达乌里秦艽单株种子产量对高原鼠兔干扰水平变化的响应表现为,随高原鼠兔干扰水平增加,达乌里秦艽单株种子产量呈增加的趋势(图5),干扰水平为Ⅳ时的单株种子产量显著高于干扰水平为Ⅲ时的单株种子产量(F(3,104)=55.408,P<0.05),而后者与干扰水平为Ⅱ时的单株种子产量差异不显著,但显著大于干扰水平为Ⅰ时的单株种子产量。
3 讨论
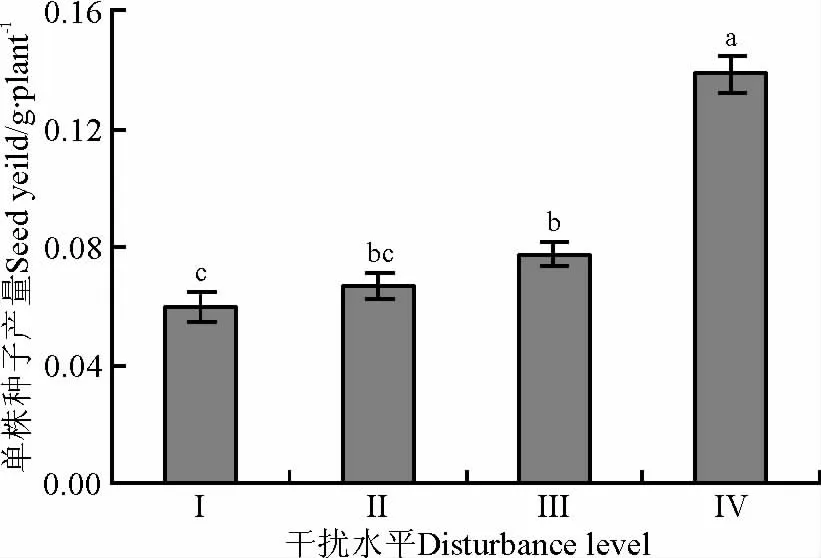
图5 高原鼠兔干扰对达乌里秦艽种子产量的影响Fig.5 Effects of plateau pika disturbance levels on seed yield per plant of Gentiana dahurica
高原鼠兔对高山嵩草草甸内不同种群的影响具有综合性特征,不仅受高原鼠兔选择性采食的影响,而且受生境变化后不同植物适应性的影响。高原鼠兔干扰水平增加意味着其选择性采食和挖掘行为的加强[2],其对高山嵩草草甸原有生境的改变愈加广泛,从而对不同植物种群的生长和繁殖产生愈为强烈的影响[6,8],迫使不同植物种群通过改变生态策略而适应变化后的生境[14-15],其中繁殖对策是植物种群适应变化后生境的主要策略之一。生物量作为生态系统植物有机物总量,是草地生态系统生产力的重要体现,也是整个生态系统运行的能量基础和营养物质来源[16]。本研究结果表明,达乌里秦艽的株高和地上生物量均随高原鼠兔干扰水平增加而具有增加态势,主要原因是随高原鼠兔干扰水平逐渐增大,单位面积内的裸斑数增加、生境逐渐旱化[17],选择性采食行为加强,这一方面为以种子繁殖为主的中旱生达乌里秦艽创造了适宜的条件[18-19],另一方面抑制了以无性繁殖为主的湿中生植物的生长[9],减轻了达乌里秦艽的竞争压力,从而促进了达乌里秦艽的株高生长,增加了生物量。
不同繁殖器官间生物量的分配指一株植物生长发育过程中所同化的资源用于茎、叶、花、果各器官的比例,它控制着植物终生生殖与生存的平衡[16]。当高原鼠兔干扰水平较低时,达乌里秦艽作为伴生种主要受优势种高山嵩草较强竞争力的抑制,利用资源的能力有限,在群落内处于相对弱势地位,植株将所同化的资源主要分配于同化器官叶,以提高同化速率,保证物种在生境内的生存[20-21]。当高原鼠兔干扰强度增大时,高山嵩草草甸生境的异质性增加,为原来处于劣势竞争条件下的伴生种达乌里秦艽提供了竞争资源的机会[9],其茎叶比增大,说明此时植株将同化产物分配至同化器官的比例减少,而将更多的同化产物分配和积累在茎器官中,促进茎器官转化成枝条,从而增加了营养枝和生殖枝的数量。随着干扰程度的增加,同化产物分配至茎器官、转变为枝条的趋向愈加明显和强烈,促进更多的枝条进入生殖生长,生殖枝数量不断增加[22],生殖枝上的花序数和花序生物量亦显著增加。随着花序数和生物量的增加,花序内的种子产量也在逐渐增加[23],从而提高了植株个体的繁殖能力。
高原鼠兔干扰增加了高山嵩草草甸伴生种达乌里秦艽的繁殖能力,从而提高了其在群落内与其他种群的竞争能力,这点从达乌里秦艽的生态位宽度随着高原鼠兔干扰程度的增加而增加中得到佐证[9],说明达乌里秦艽利用资源的能力随高原鼠兔干扰程度的增加而增加,这在一定程度上揭示了高原鼠兔过度干扰劣化高山嵩草草甸品质的途径,即高原鼠兔过度干扰提高了以种子繁殖为主的家畜不可食植物的繁殖能力,增加了其在群落内的比例,从而整体上劣化了高山嵩草草甸的放牧品质。
[1] 郭伟娜,魏朔南.秦艽的生物学研究[J].中国野生植物资源,2008,27(4):1-10.
[2] 庞晓攀,贾婷婷,李倩倩,雒明伟,肖玉,赵旭,郭正刚.高原鼠兔有效洞穴密度对高山嵩草群落及其主要种群空间分布特征的影响[J].生态学报,2015,35(3):872-884.
[3] 唐建宁,杨云飞,李满,许强,王俊.六盘山区秦艽植物资源及其利用研究[J].农业科学研究,2006,27(1):59-62.
[4] 陈士林,索风梅,韩建萍,谢彩香,姚辉,李西文,李滢,魏建和.中国药材生态适宜性分析及生产区划[J].中草药,2007,38(4):481-487.
[5] Smith A T,Foggin J M.The plateau pika(Ochotona cur zoniae)is a keystone species for biodiversity on the Tibetan Plateau[J].Ani mal Conservation,1999,2(4):235-240.
[6] Pech R P,Arthur A D,Zhang Y M,Lin H.Population dynamics and responses to management of plateau pikas(Ochotona curzoniae)[J].Journal of Applied Ecology,2007,44(3):615-624.
[7] Delibes-Mateos M,Smith A T,Slobodchikoff C N,Swenson J E.The paradox of keystone species persecuted as pests:A call for the conservation of abundant s mall mammals in their native range[J].Biological Conservation,2011,144:1335-1346.
[8] Eldridge D J,Whitford W G.Disturbances by desert rodents are more strongly associated with spatial changes in soil texture than woody encroach ment[J].Plant and Soil,2014,381:395-404.
[9] 贾婷婷,毛亮,郭正刚.高原鼠兔有效洞穴密度对青藏高原高寒草甸群落植物生态位的影响[J].生态学报,2014,34(4):1-9.
[10] 何维明,钟章成.植物繁殖对策的概念及其研究内容[J].生态学杂志,1997,14(6):1-3.
[11] Guo Z G,Li X F,Liu X Y,Zhou X R.Response of alpine meadow communities to burrow density changes of plateau pika(Ochotona cur zoniae)in the Qinghai-Tibet Plateau[J].Acta Ecologica Sinica,2012,32:44-49.
[12] 李倩倩,赵旭,郭正刚.高原鼠兔有效洞穴密度对高寒草甸优势植物叶片和土壤氮磷化学计量特征的影响[J].生态学报,2014,34(5):1213-1223.
[13] Guo Z G,Zhou X R,Hou Y.Effect of available burr ow densities of plateau pika(Ochotona cur zoniae)on soil physicochemical property of t he bare land and vegetation land in t he Qinghai-Tibetan Plateau[J].Acta Ecological Sinica,2012,32(2):104-110.
[14] 周雪荣,郭正刚,郭兴华.高原鼠兔和高原鼢鼠在高寒草甸中的作用[J].草业科学,2010,27(5):38-44.
[15] Davidson A D,Lightfoot D C.Burrowing rodents increase landscape heter ogeneity in a desert grassland[J].Jour nal of Arid Environ ments,2008,72(7):1133-1145.
[16] 方炎明.植物生殖生态学[M].济南:山东大学出版社,1996:1-109.
[17] Liu H,Chen Y,Zhou L,Jin Z.The effects of management on population dynamics of plateau pika[J].Mathematical and Co mputer Modelling,2013,57:525-535.
[18] 王文颖,王启基,景增春,李世雄,史惠兰.江河源区高山嵩草草甸覆被变化对植物群落特征及多样性的影响[J].资源科学,2006,28(2):118-124.
[19] Alcántara J M,Rey P J,Sánchez-Laf uente A M,Valera F.Early effects of r odent post-dispersal seed predation on the outco me of the plant-seed disperser interaction[J].Oikos,2000,88(2):362-370.
[20] Weiher E,Vandererf A,Thompson K,Roderick M,Garnier E,Eriksson O.Challenging the ophrastus:A common core list of plant traits for f unctional ecology[A].Proceeding of 41st Sy mposiu m of the International Association for Vegetation Science[C].Uppsala,Sweden:41st Sy mposiu m of t he Inter national Association for Vegetation Science,1998:609-620.
[21] 任海彦,郑淑霞,白永飞.放牧对内蒙古锡林河流域草地群落植物茎叶生物量资源分配的影响[J].植物生态学报,2009,33(6):1065-1074.
[22] 郝虎东.无芒雀麦资源分配及其繁殖数量特征研究[D].呼和浩特:内蒙古农业大学硕士论文,2009.
[23] 刘左军,杜国祯,陈家宽,刘振恒,董高生,马建云.影响黄帚橐吾种子生产的因素Ⅰ.生境和花序结构[J].植物生态学报,2003,27(5):677-683.
