Rho 型GTPase 激活蛋白MoBem2 在稻瘟病菌分生孢子形态建成中的功能
2015-04-02齐中强刘永锋
齐中强, 杜 艳, 刘永锋
(江苏省农业科学院植物保护研究所,江苏 南京210014)
稻瘟病已经成为危害世界粮食安全生产的一种重要病害[1-4]。由于经济重要性和遗传转化的易操作性以及全序列的公布,稻瘟病菌-植物互作模式已经成为研究的热点课题[5]。稻瘟病菌分生孢子在其侵染水稻的过程中扮演着重要角色[6]。分生孢子首先在其顶端产生粘性物质,随后便萌发产生芽管,最后分化形成附着胞。附着胞的分化主要受细胞周期和程序性死亡等因素调控。附着胞内部通过甘油的积累可以产生大约8 MPa 的膨压,从而产生侵入钉,穿破寄主表皮进入细胞内。侵入后,侵入钉会分化成侵染菌丝,进而在植物细胞内定殖,随着时间的延长,就会形成可见的典型稻瘟病斑。当外界条件(温度、湿度)适宜时,病组织上会重新长出孢子梗,进而产生分生孢子,继续进行再侵染[7]。Rho 蛋白是Ras 超家族中最早被克隆出来的一类蛋白质,它们是一组相对分子质量约为2.0×104~2.5×104的三磷酸鸟苷结合蛋白,具有GTP 酶活性,主要调控细胞骨架的形成及细胞形态建成[8]。Rho 蛋白和其他类型的G 蛋白具有相似的功能,作为一种分子开关,具有GTP 结合的活性状态和GDP 结合的失活状态。激活蛋白Bem2主要行使GTPase 激活功能,促使GTP 向GDP 的转换,加速Rho 蛋白的失活。为了明确Rho 蛋白GTPase 激活蛋白在稻瘟病菌中的功能,本研究通过基因敲除的方法,对稻瘟病菌中Rho 型GTPase 激活蛋白MoBem2 功能进行解析。
1 材料与方法
1.1 供试材料
稻瘟病菌(Magnaporthe oryzae)Guy11 和水稻品种CO39(感病品种)由南京农业大学植物保护学院真菌与卵菌实验室提供。完全培养基CM、产孢培养基SDC 的配制方法参照文献[9]。
1.2 试验方法
1.2.1 敲除载体构建及稻瘟病菌敲除转化 通过稻瘟病菌全基因组序列公布网站,下载预测到的MoBEM2 基因序列。将稻瘟病菌基因组数据库中MoBEM2 基因上、下游各1 kb 左右的DNA 序列作为同源重组的上、下两臂,构建基因敲除载体。以野生型Guy11 基因组DNA 为模板,分别用引物bem2-p1(F)/bem2-p2(R)和bem2-p3(F)/bem2-p4 (R)扩增上、下臂片段,经电泳、切胶回收将PCR 产物纯化。再以这两臂片段为模板,用引物bem2-p1(F)/bem2-p4(R)进行Over-lap PCR 扩增。PCR 产物切胶回收后连接到pMD19-T simple vector(TaKaRa,Dalian,China)得到质粒pMD∶∶BEM2,并送交上海Invitrogen 公司测序,其中引物bem2-p2 和bem2-p3序列中设有EcoRV 酶切位点。以质粒pCB1003 为模板,用引物FL1111 (F)/FL1112 (R)高保真PCR扩增抗性筛选基因—潮霉素磷酸转移酶基因(HPH),所用的高保真酶为Primer STAR(TaKaRa,Dalian,China),反应程序为:98 ℃10 s,56 ℃15 s,72 ℃1 min,30 个循环。扩增得到的平末端PCR 产物切胶回收后插入EcoR V 酶切过的质粒pMD∶∶BEM2 中,得到敲除载体pMD∶∶BEM2∶∶HPH。以该质粒为模板,用引物bem2-p1(F)/bem2-p4(R)扩增得到约3.4 kb 的敲除片段用于稻瘟病菌的原生质体转化,原生质体转化方法参照文献[10]。
1.2.2 转化子验证及Southern 杂交 提取转化得到的转化子基因组DNA(CTAB 法),用引物bem2-p5(F)/bem2-p6(R)进行验证,随后将验证得到的候选突变体进行Southern 杂交。以稻瘟病菌野生型Guy11 基因组DNA 为模板,用引物bem2-p5(F)/bem2-p6(R)扩增出MoBEM2 基因内部889 bp 片段,以此片段作为探针,并用地高辛标记。Guy11 菌丝基因组DNA 经EcoR I 酶切于1.0%琼脂糖凝胶中过夜电泳,充分分离酶解片段,然后转移至带正电的尼龙膜(HybondTM-N+,Amersham,Biosciences UK Limited)上,与地高辛标记的探针于58 ℃杂交过夜。Southern 杂交过程参照Digoxigenin high-prime DNA labeling and the detection starter kit 1(Roche,Germany)的操作手册。以上引物序列见表1。
1.2.3 分生孢子的诱导产生 将野生型Guy11 及敲除突变体接种在SDC 培养基上,28 ℃黑暗培养7 d 左右,待菌丝体长满平板后,用手术刀将表面气生菌丝刮掉,于黑光灯下照射3 d,诱导分生孢子产生。收集孢子时,向培养基内加入3 ml 无菌水,轻轻用1.5 ml EP 管底部将表面气生菌丝和孢子刷下,再经过4 层擦镜纸过滤收集孢子。
1.2.4 附着胞形成的观测 将盖玻片(Fisherbrand,12-540-A 18 ×18-2)放置在载玻片上(下面滴加无菌水),取40 μl 浓度为1 ml 5 ×104个的分生孢子液,滴加于盖玻片中央,随后将载玻片放入培养皿中28 ℃黑暗保湿培养2 h、4 h、6 h、8 h、24 h 和48 h后分别制片观测附着胞形成率。设置3 个重复。
1.2.5 水稻喷雾接种及大麦离体致病性测定 将刷下的分生孢子液浓度调至1 ml 5 ×104个,加入0.25%明胶,喷雾接种生长14 d 的水稻,28 ℃黑暗培养24 h,随后16 h 光照,8 h 黑暗,处理期间需要保持高温高湿状态。接种5 ~7 d 后,观察结果。大麦离体致病性测定:剪取生长14 d 的大麦叶片铺于含有保湿滤纸的培养皿中,将孢子液浓度调至1 ml 5 ×104个,加入0.25%明胶,每片叶片滴3 滴孢子液,以清水做对照,置于28 ℃黑暗培养24 h,随后16 h 光照,8 h 黑暗处理。接种5 ~7 d 后,观察结果。试验重复3 次。

表1 本研究中使用的引物Table 1 Primers used in this study
1.2.6 水稻叶鞘侵染 取生长28 d 的水稻植株,剥去外面叶片,留存倒数第2 片叶鞘和新叶,将新叶慢慢取出,保留叶鞘部分。收集野生型和突变体分生孢子,调配孢子液浓度为1 ml 1 ×105个,用1 ml注射器将孢子液注入叶鞘内,随后放入保湿培养皿中保湿培养,水稻根部覆盖脱脂棉保湿。48 h 后,用手术刀将叶鞘切断,选取距根部较近部位用眼科镊撕取叶鞘内表皮,随后制片观察并拍照。每个菌株处理10 株水稻叶鞘。
1.2.7 分生孢子荧光增白剂(CFW)染色 将10 mg/ml的CFW 染色液稀释1 000倍,加入准备好的孢子悬浮液中,避光染色5 min,随后用无菌水冲洗数次,置于荧光显微镜下观察。
2 结果与分析
2.1 稻瘟病菌MoBEM2 基因敲除突变体的获得
利用同源重组原理对稻瘟病菌中的MoBEM2基因进行了定向敲除(图1),经初步验证获得#23和#32 2 个转化子。Southern 杂交结果(图2)显示,两个突变体用基因探针没有杂交出4.7 Kb 条带,说明已被成功敲除,进一步用潮霉素探针杂交出了约6.7 Kb 条带,证明MoBEM2 基因已被潮霉素基因成功替换。
2.2 MoBEM2 基因敲除突变体的孢子形态
将稻瘟病菌野生型Guy11 和#23 突变体及互补转化子分别接种到SDC 培养基中,随后诱导产孢。发现突变体分生孢子产量显著降低(下降40%左右)(图3),孢子形态发生改变(图4),分生孢子只形成一个隔膜(约占40%左右),而野生型中超过90%均为正常孢子(图5),说明MoBem2 参与了稻瘟病菌分生孢子的形态建成。

图1 MoBEM2 基因替换策略简图Fig.1 Schematic illustration of MoBEM2 targeted gene replacement
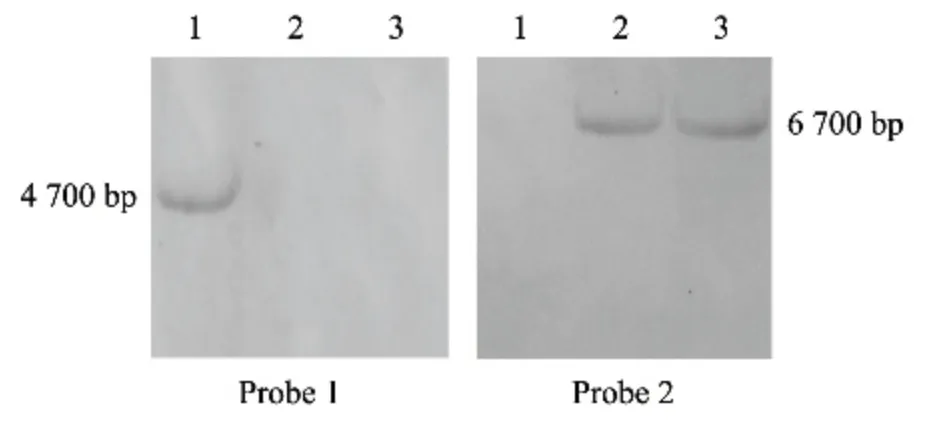
图2 MoBEM2 敲除突变体Southern 杂交验证Fig.2 Southern blot of MoBEM2 deletion mutant
2.3 MoBEM2 基因敲除突变体附着胞的形成率
稻瘟病菌附着胞的形成对其致病性是必须的[11],因此检测了MoBEM2 敲除突变体附着胞形成情况。结果(图6)表明,突变体分生孢子在培养8 h时附着胞形成率显著低于野生型和互补转化子,但随着时间延长,该缺陷恢复,至24 h,突变体附着胞形成率与野生型没有差异。说明MoBEM2 基因敲除延缓了稻瘟病菌附着胞的形成。
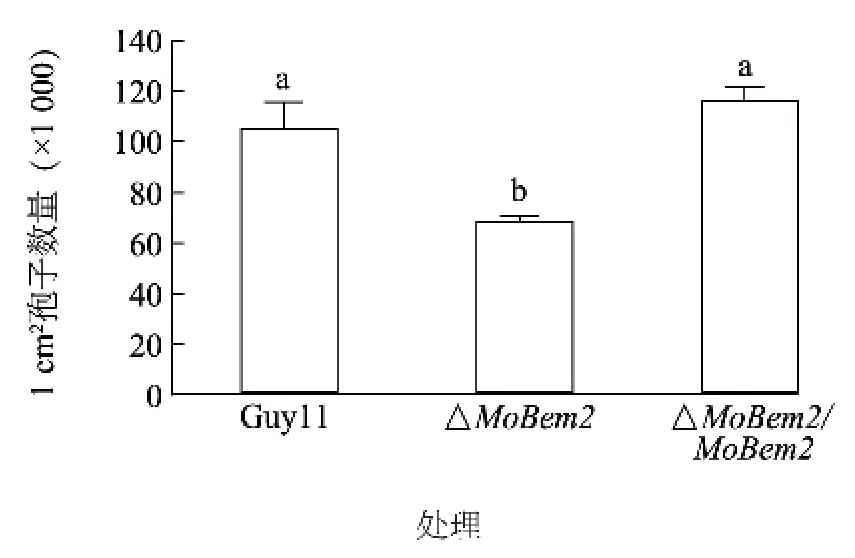
图3 MoBEM2 敲除突变体分生孢子产量Fig.3 Conidial production of MoBEM2 deletion mutant
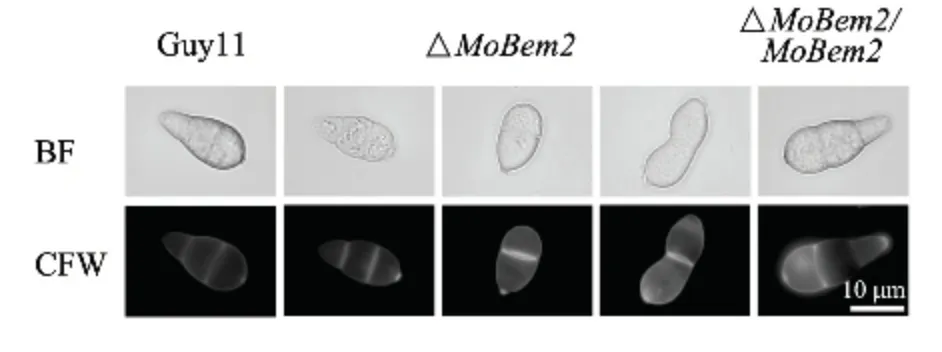
图4 MoBEM2 敲除突变体分生孢子形态Fig.4 Conidial morphology of MoBEM2 deletion mutant

图5 MoBEM2 敲除突变体不同形态分生孢子百分比Fig.5 Percentage of uniseptate and normal conidial morphology of MoBEM2 deletion mutant

图6 MoBEM2 突变体附着胞形成率Fig.6 Appressorium formation rate of MoBEM2 deletion mutant
2.4 MoBEM2 基因敲除突变体的致病性
将野生型、敲除突变体和互补转化子的分生孢子配制成浓度为1 ml 5.0 ×104个的孢子悬浮液(含0.25%明胶),分别采用水稻植株喷雾和大麦离体叶片点滴的方法进行致病性测定。结果显示,在两种接种条件下,敲除突变体发病情况与野生型、互补转化子均无差异,表明MoBEM2 基因在稻瘟病菌致病过程中不起作用(图7)。

图7 MoBEM2 敲除突变体对水稻和大麦的致病性Fig.7 Pathogenicity to rice (A)and barley (B)of MoBEM2 deletion mutant
3 讨论
Rho 型蛋白通过与特异性的鸟嘌呤核苷酸转换因子(GEF)结合而被激活,与GTP 酶激活蛋白(GAPs)结合而失活[12]。本试验以稻瘟病菌中一个Rho 型蛋白GTP 酶激活蛋白MoBem2 为研究对象,通过基因敲除,发现MoBem2 参与稻瘟病菌分生孢子的形态建成,延缓附着胞的形成。
分生孢子在稻瘟病菌的致病过程中具有重要作用[13]。稻瘟病菌中,MoRac1 主要参与稻瘟病菌分生孢子的形成以及形态建成,并且调控稻瘟病菌对水稻的致病力[14]。本研究中,敲除Rho 型GTP 酶激活蛋白编码基因MoBEM2 后,该突变体出现单隔的分生孢子,分生孢子产量也显著下降,推测该基因参与分生孢子的形成及形态建成。上述结果表明Rho 蛋白主要通过其激活和失活调控稻瘟病菌分生孢子的形态建成,在这个过程中,GTP 酶的作用尤为明显。此外,MoHOX2 作为Homeobox 转录因子家族的成员,敲除后稻瘟病菌丧失产孢能力[15]。2 个SNARE 蛋白MoVam7[16]和MoSec22[17]同样参与了稻瘟病菌的产孢过程,两者的敲除突变体均丧失产孢能力。说明了稻瘟病菌分生孢子的调控涉及多种机制。
附着胞在稻瘟病菌侵染过程中起着至关重要的作用[18]。MoBEM2 敲除突变体分生孢子在诱导界面上能够形成附着胞,但在附着胞形成早期,突变体附着胞形成率显著降低,但在后期与野生型没有差别。说明该基因一定程度上影响稻瘟病菌附着胞早期的形成,但不影响稻瘟病菌的致病性。综上所述,MoBem2 参与了稻瘟病菌分生孢子的形成及形态建成过程,这对认识稻瘟病菌致病过程有重要意义。
[1] WILSON R A,TALBOT N J. Under pressure:investigating the biology of plant infection by Magnaporthe oryzae[J]. Nature Reviews Microbiology,2009,7:185-195.
[2] 于 涛,张海楼,隽英华,等.施肥模式对水稻稻瘟病抗性的影响[J].江苏农业科学,2014,42(7):113-116.
[3] 张晓娟,张 羽,张辰露,等.分子标记在稻瘟病抗性育种中应用的研究进展[J].江苏农业科学,2013,41(8):73-75.
[4] 唐 成,陈 露,安敏敏,等.稻瘟病诱导水稻幼苗叶片氧化还原系统的特征谱变化[J]. 江苏农业科学,2014,42(12):141-144.
[5] DEAN R A,TALBOT N J,EBBOLE D J,et al. The genome sequence of the rice blast fungus Magnaporthe grisea[J]. Nature,2005,434:980-986.
[6] HAMER J E,HOWARD R J,CHUMLEY F G,et al. A mechanism for surface attachment in spores of a plant pathogenic fungus[J]. Science,1988,239,288-290.
[7] TALBOT N J. On the trail of a cereal killer:Exploring the biology of Magnaporthe grisea[J]. Annu Rev Microbiol,2003,57:177-202.
[8] JOHNSON D I. Cdc42:An essential Rho-type GTPase controlling eukaryotic cell polarity[J]. Microbiol Mol Biol Rev,1999,63:54-105.
[9] ZHANG H,TANG W,LIU K,et al. Eight RGS and RGS-like proteins orchestrate growth,differentiation,and pathogenicity of Magnaporthe oryzae[J]. PLoS Pathogens,2011,7:e1002450.
[10] QI Z,WANG Q,DOU X,et al. MoSwi6,an APSES family transcription factor,interacts with MoMps1 and is required for hyphal and conidial morphogenesis,appressorial function and pathogenicity of Magnaporthe oryzae[J]. Molecular Plant Pathology,2012,13:677-689.
[11] THINES E,WEBER R W,TALBOT N J. MAP kinase and protein kinase A-dependent mobilization of triacylglycerol and glycogen during appressorium turgor generation by Magnaporthe grisea[J]. Plant Cell,2000,12:1703-1718.
[12] JAFFE A B,HALL A. Rho GTPases:biochemistry and biology[J]. Ann Rev Cell Dev Biol,2005,21:247-269.
[13] TALBOT N J,EBBOLE D J,HAMER J E. Identification and characterization of MPG1,a gene involved in pathogenicity from the rice blast fungus Magnaporthe grisea[J]. The Plant cell,1993,5:1575-1590.
[14] CHEN J,ZHENG W,ZHENG S,et al. Rac1 is required for pathogenicity and Chm1-dependent conidiogenesis in rice fungal pathogen Magnaporthe grisea [J]. PLoS Pathogens,2008,4:e1000202.
[15] KIM S,PARK S Y,KIM K S,et al. Homeobox transcription factors are required for conidiation and appressorium development in the rice blast fungus Magnaporthe oryzae[J]. PLoS Genetics,2009,5:e1000757.
[16] DOU X,WANG Q,QI Z,et al. MoVam7,a conserved SNARE involved in vacuole assembly,is required for growth,endocytosis,ROS accumulation,and pathogenesis of Magnaporthe oryzae[J].Plos One,2011,6 :e16439.
[17] SONG W W,DOU X Y,QI Z Q,et al. R-SNARE homolog moSec22 is required for conidiogenesis,cell wall integrity,and pathogenesis of Magnaporthe oryzae[J]. PLoS One,2010,5:e13193.
[18] JEON J,GOH J,YOO S,et al. A putative MAP kinase kinase kinase,MCK1,is required for cell wall integrity and pathogenicity of the rice blast fungus,Magnaporthe oryzae[J]. Mol Plant Microbe Interact,2008,21,525-534.
