多棘海盘车对魁蚶摄食量、选择性及昼夜摄食差异的初步研究❋
2015-03-31张天文刘广斌刘恩孚郑永允邱兆星
张天文, 刘广斌, 刘恩孚, 郑永允, 邱兆星
(山东省海洋生物研究院, 山东 青岛 266002)
多棘海盘车对魁蚶摄食量、选择性及昼夜摄食差异的初步研究❋
张天文, 刘广斌, 刘恩孚, 郑永允, 邱兆星❋❋
(山东省海洋生物研究院, 山东 青岛 266002)
在水温16~20 ℃、盐度28.2±2.1、溶解氧(7.5±0.7)mg/L、pH8.0±0.3的条件下,研究了3种规格多棘海盘车(Asteriasamurensis)对4种规格魁蚶(Scapharcabroughtonii)的摄食量、摄食选择性和摄食昼夜差异。研究表明:多棘海盘车对不同规格魁蚶摄食数量均随魁蚶底播密度的增加而上升;不同规格多棘海盘车对同一规格魁蚶摄食数量无显著差异(P>0.05),但对不同规格魁蚶摄食数量存在一定差异,对最小壳长组魁蚶的摄食数量最大;多棘海盘车对不同规格魁蚶软体部摄食量与魁蚶规格呈显著正相关;多棘海盘车对不同规格魁蚶摄食数量均存在一定的昼夜差异且夜间的摄食数量大于昼间,魁蚶规格越大,多棘海盘车的最大摄食数量的时间出现越晚。结果表明:魁蚶适宜底播密度在30 ind/m2左右,底播规格越大越能有效防御敌害。根据多棘海盘车对贝类捕食特点,可采用浅水低值贝类作为诱捕生物对多棘海盘车进行清除。
多棘海盘车;魁蚶;摄食量;摄食选择性;昼夜差异
魁蚶(Scapharcabroughtonii)俗称赤贝、血贝,属于瓣鳃纲蚶科(Arcidae),是一种营埋栖生活的大型双壳贝类,肉味鲜美、营养丰富,有较高的经济价值,一直是黄渤海区渔民的兼捕对象。由于长期过量捕捞,近十余年来其自然资源极度萎缩[1],天然采捕产量已远远不能满足国内外市场的需求,发展增养殖和资源修复势在必行[2]。2010年以来,魁蚶增养殖已成为继海参养殖之后,开发利用海洋浅海资源的又一热点[3]。影响魁蚶增养殖效果的因素有很多,底播后最主要的影响因素是敌害生物海星的侵噬[4]。
海星是潮间带及潮下带生态系统中主要的捕食者之一[5-6],其摄食作用可以直接或间接影响底栖生物的分布及丰度[7-10]。山东沿海分布的海星主要为多棘海盘车(Asteriasamurensis)[11],其属于海星纲海盘车属(Asterias)。多棘海盘车主要摄食贝类、甲壳类、多毛类甚至小型鱼类[12],特别喜食双壳类,而且与底栖生物竞争饵料[13-14]。自然环境中多棘海盘车食性虽广,但对饵料生物具有一定的选择性,这种选择性摄食不仅给自身带来更多的生存机会,也具有调节饵料生物种群数量的重要作用[15]。在贝类养殖区,多棘海盘车可以依据贝类气味精确定位埋栖型贝类并挖掘摄食[16-17]。1954年在东京湾仅由多棘海盘车摄食所引起的养殖贝类直接损失就高达4亿多日元[18],2007年青岛沿海多棘海盘车爆发,导致红岛地区底播贝类损失严重[19]。本文通过研究不同规格多棘海盘车对不同规格、不同密度魁蚶的摄食选择性及昼夜摄食差异,为了解多棘海盘车对魁蚶的摄食生态学提供基础数据,以期有效防止主要敌害生物影响,提高魁蚶的存活率,更好的开展魁蚶底播增殖工作。
1 材料和方法
1.1 实验围格及管理
2013年3—6月,在日照市水产研究所,选取泥沙底质池塘开展实验,采用围格实验生态学方法研究多棘海盘车对魁蚶稚贝的摄食能力及昼夜摄食差异。实验在2 m×2 m的围隔中开展,以30目筛绢做围隔幔,围隔幔下部埋入池塘底泥0.6 m,整个围隔以木桩和青竹为支架架设于池塘中。在实验开始前2 d进水,实验时间为96 h,实验过程中池水深度稳定在(1.0±0.1) m,结束后排空池水进行统计。实验时水温为16~20 ℃,盐度为28.2±2.0,溶解氧为(7.5±0.7)mg/L,pH为8.0±0.3。
1.2 实验材料
实验用魁蚶稚贝来自山东省日照市山海天旅游度假区两城河口海域,为2012年经过越冬培育的苗种。运到实验场地后在暂养池中用循环水暂养14d,期间每天06:00、12:00、18:00、24:00分4次投喂金藻和硅藻。
实验用多棘海盘车采自山东省日照市东港区海域,运到实验场地后,在暂养池中用循环水暂养3d,暂养期间不投喂任何饵料。
1.3 实验设计
1.3.1 多棘海盘车对不同密度、不同规格魁蚶稚贝的摄食量 选取壳长分别为(1.00±0.09)、(1.50±0.13)、(2.00±0.12)和(2.50±0.15)cm 4种规格魁蚶稚贝作为实验对象,记做1.0、1.5、2.0和2.5 cm组,测量4种规格魁蚶稚贝软体部重量,以10、30和50 ind/m23个密度作为实验密度;采用腕足长度在8~10 cm,密度为1 ind/m2的多棘海盘车进行实验,分析多棘海盘车对不同密度不同规格魁蚶的摄食能力,实验时间为96 h。每组实验设置4个平行,同时每组实验设置4个无多棘海盘车的空白对照组。
1.3.2 不同规格的多棘海盘车对不同规格魁蚶稚贝的摄食量 选取腕长分别为5~7、8~10和11~13 cm的3种规格多棘海盘车,记做小型、中型和大型多棘海盘车。魁蚶密度为30 ind/m2,多棘海盘车密度为1 ind/m2。分析3种规格的多棘海盘车对4种规格的魁蚶摄食能力差异,实验时间为96 h。每组实验设置4个平行,同时每组实验设置4个无多棘海盘车的空白对照组。
1.3.3 多棘海盘车对不同规格魁蚶稚贝摄食的昼夜差异 多棘海盘车对4种规格魁蚶摄食昼夜差异实验中,魁蚶密度为30 ind/m2,采用规格为腕足长8~10 cm、密度为1 ind/m2的多棘海盘车进行实验,实验时间为96 h。实验于上午8:00开始,按照昼夜变化规律选取实验开始后1、2、8、12、24、30、46、54、60、72和96 h作为观测时间。每组实验设置4个平行,同时每组实验设置4个无多棘海盘车的空白对照组。
1.4 实验指标测定
不同规格多棘海盘车对不同规格、不同密度魁蚶摄食实验中,实验结束后清点每个围格中的魁蚶空壳数E,记录魁蚶苗种的自然死亡数D。
多棘海盘车对不同规格魁蚶稚贝摄食的昼夜差异实验中,按照选取的观测时间,分别记录实验围格中魁蚶空壳数Et和空白组魁蚶空壳数Dt。
1.5 数据处理及分析
不同规格的多棘海盘车对不同规格魁蚶的摄食量以被摄食魁蚶的空壳数来表示。
K=E-D。
式中:K为被摄食魁蚶的空壳数量(ind);E为实验围格中魁蚶总的空壳数(ind);D为无多棘海盘车的空白对照组中魁蚶的总空壳数(ind)。
多棘海盘车对不同规格魁蚶摄食昼夜差异以不同观测时间段魁蚶空壳数占总的空壳数百分比来表示。
Ht=(Et-Dt)/(E-D)。
式中:t为观测时间;Ht为截止该观测时间被摄食魁蚶占实验期间所有被摄食魁蚶的百分比;Et为该时间实验围格中魁蚶空壳数(ind);Dt为该时间无多棘海盘车的空白对照组中魁蚶的空壳数(ind)。
实验数据采用统计软件SPSS17.0进行方差分析,利用Duncan法进行多重比较,如P>0.05则认为差异不显著,如P<0.05则认为差异显著,结果用小写字母表示,有相同字母的数据表示相互差异不显著(P>0.05)。实验过程中魁蚶壳长记做L,魁蚶肉重记做W。
2 实验结果
2.1 多棘海盘车对不同密度、不同规格魁蚶的摄食数量分析
由图1可以看到,多棘海盘车对不同规格的魁蚶摄食数量随魁蚶密度的增加而升高。当魁蚶密度为30 ind/m2时,多棘海盘车对魁蚶摄食数量虽所提高,但与魁蚶密度在10 ind/m2时差异不显著(P>0.05)。当魁蚶密度为50 ind/m2,多棘海盘车对4种规格的魁蚶摄食数量显著增加。在同一密度时,多棘海盘车对魁蚶的摄食数量随魁蚶壳长的增加而减少,其中2.5 cm组在各个密度下摄食数量都是最小的。多棘海盘车对不同规格的魁蚶摄食数量与魁蚶规格呈成显著负相关(r=-0.9845,P<0.01),当L>0时,两者之间拟合公式为:K=35.543e-0.457L(R2=0.9793)。

(图中标有相同字母的数据表示相互差异不显著(P>0.05)。Data with the same superscript are not significantly different(P>0.05).)
图1 多棘海盘车对不同密度、不同规格魁蚶摄食数量分析
Fig.1 The amount of food intake ofAsteriasamurensison different kinds and density ofScapharcabroughtonii
由图2可以看到多棘海盘车对不同密度魁蚶软体部摄食量随魁蚶密度的增大而增加。4种不同规格的魁蚶在密度为50 ind/m2时被摄食的软体部重量均为最大,在密度为10和30 ind/m2时被摄食的软体部重量差异不显著;多棘海盘车对不同规格魁蚶软体部摄食量也呈现随魁蚶规格的增大而增加的趋势,在3种密度下1 cm实验组软体部被摄食量均为最小,2.5 cm实验组软体部被摄食量均为最大,但2 cm实验组和2.5 cm实验组之间没有显著差异。多棘海盘车对不同规格的魁蚶软体部摄食量与魁蚶规格呈成显著正相关(r=0.9873,P<0.01),两者之间拟合公式为:W=1.4862L(R2=0.972 8)。

(图中标有相同字母的数据表示相互差异不显著(P>0.05)。Data with the same superscript are not significantly different(P>0.05).)
图2 多棘海盘车对不同密度、不同规格魁蚶软体部摄食量分析
Fig.2 The weight of food intake ofAsteriasamurensison different kinds and density ofScapharcabroughtonii
2.2 不同规格多棘海盘车对不同规格魁蚶的摄食数量分析
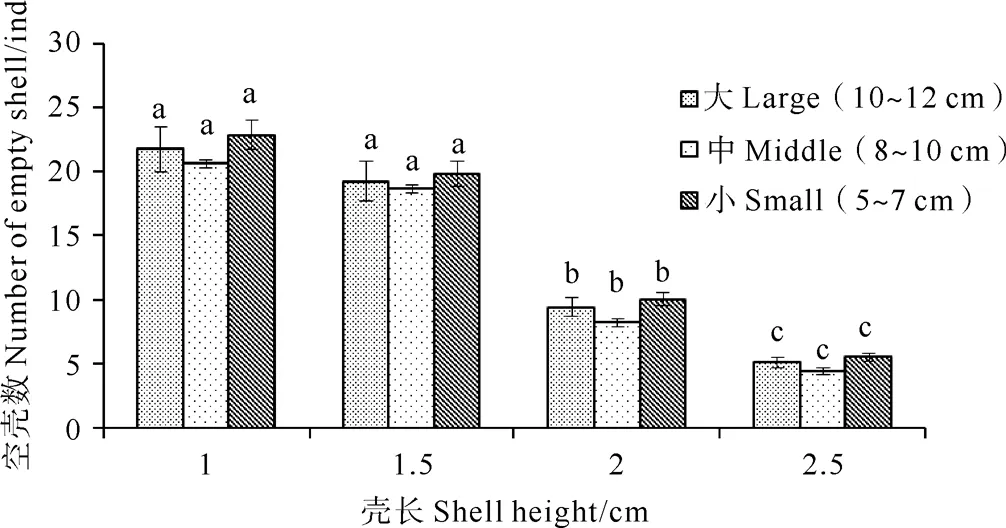
由图3可以看到,不同规格的多棘海盘车对同一规格魁蚶的摄食数量无显著差异(P>0.05),但同一规格的多棘海盘车对不同规格魁蚶的摄食数量存在一定的差异。其中1 cm组和1.5 cm组被摄食数量差异不显著,随着实验组魁蚶规格的增大,被摄食数量逐渐降低,3种规格的多棘海盘车对2.5 cm组摄食数量均为最低(平均为5 ind)。在4个实验组中,腕足长度在5~7 cm的多棘海盘车对魁蚶摄食数量略高于腕足长度8~10 cm和10~12 cm组,但差异不显著(P>0.05)。
2.3 多棘海盘车对不同规格魁蚶的摄食数量昼夜差异
多棘海盘车对4种规格魁蚶摄食数量均存在一定的昼夜差异(见图4),摄食时间主要集中在晚上时段(20:00—8:00)。8:00开始实验,4种规格魁蚶在12 h时(第一天20:00)均未被摄食;到24 h观测时(第二天8:00),除了2.5 cm组之外,其他3种规格魁蚶被多棘海盘车摄食,其中2 cm组被摄食比例最大均为14.29%;至30 h观测时(第二天14:00),除了1 cm组魁蚶被少量摄食之外,其他3个规格实验组均未见多棘海盘车摄食魁蚶;至48 h观测时(第三天8:00),4种规格魁蚶均大量被多棘海盘车摄食,所占比例为所有观测时间中最高的;至54 h观测时(第三天14:00),只有1 cm组有少量魁蚶被摄食,其他3个实验组未发现有新增空壳;至60 h观测时(第三天20:00),除2.5 cm组外,其他3个规格实验组均有少量魁蚶被摄食;至72 h(第四天8:00),4种规格实验组魁蚶均有被摄食现象,但是摄食比例显著低于48h观测时;至96 h(第五天8:00),4种规格实验组魁蚶均有被摄食现象,摄食所比例最低。
(图中标有相同字母的数据表示相互差异不显著(P>0.05)。Data with the same superscript are not significantly different (P>0.05).)
图3 不同规格多棘海盘车对不同规格魁蚶的摄食数量分析
Fig.3 Feeding intensity of different kindsAsteriasamurensison different kindsScapharcabroughtonii
3 讨论
3.1 不同规格多棘海盘车对不同密度、不同规格魁蚶的摄食分析
多棘海盘车食性广泛,处于生物群落食物链顶端[20],摄食对象包括环境中各种可利用的饵料生物,并有一定的选择性[21]。这种选择性是依据多棘海盘车对饵料生物的喜好以及环境中这种饵料生物的多寡、可获性等来决定。国外关于海星捕食关系及摄食选择性的研究起步较早,如太平洋地区多棘海盘车、福氏海盘车等对扇贝、贻贝规格选择性及功能反应(Functional response)等相关研究[22-25],而关于多棘海盘车对魁蚶摄食选择性的研究还未见报道。
根据最适食谱模型,当食物的数量达到一定密度后,捕食者会拒绝吃有利性较小的猎物[26]。在本实验中也发现多棘海盘车对魁蚶软体部摄食量随魁蚶规格和密度的增加而增大。当底播密度为50 ind/m2时,多棘海盘车对4种规格魁蚶软体部摄食量都是最大的。当底播密度为10和30 ind/m2时,4种规格魁蚶被多棘海盘车摄食量无显著差异(P>0.05)。
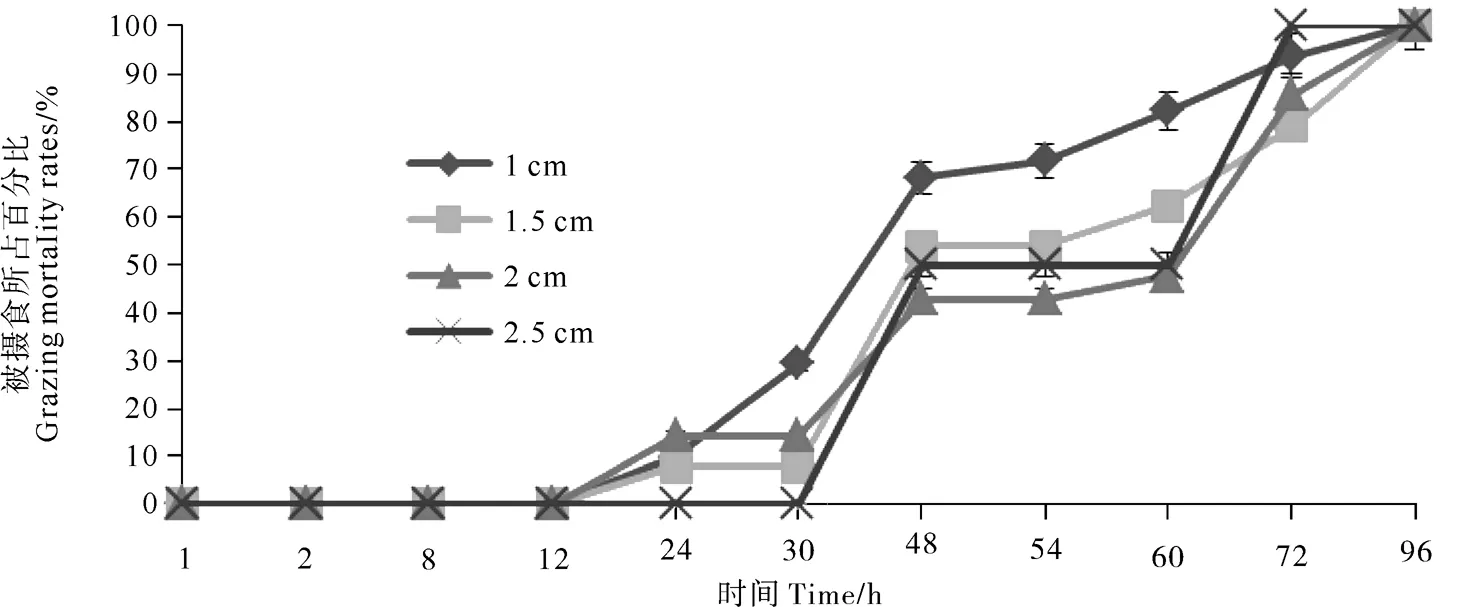
图4 多棘海盘车对不同规格魁蚶稚贝摄食数量昼夜差异
在魁蚶底播生产中,如果选择密度为10 ind/m2,虽然被敌害生物摄食少,但因底播密度低,其产量和经济效益欠佳;如果选择密度为50 ind/m2,又会有大量的魁蚶被敌害生物摄食,造成不必要的经济损失。因此建议魁蚶底播时密度为30 ind/m2左右,既保证单位面积魁蚶数量,又可以避免多棘海盘车对魁蚶摄食造成过多损失,有助于取得较好的增养殖效果,这也与于瑞海等[3]研究认为魁蚶底播密度一般选择在15~30 ind/m2之间相近。
海洋生态系统中摄食者的食物选择性由食物颗粒的大小决定[27],多棘海盘车对贝类的摄食与贝类大小有显著关系。根据Kim Y[18]等的研究成果,多棘海盘车对各种壳长贝类都具有摄食能力,对适宜壳长的贝类摄食率最大。在本实验中,3种规格的多棘海盘车虽对同一规格魁蚶摄食数量无显著差异,但对不同规格魁蚶的摄食数量之间存在差异,其中对1和1.5 cm组魁蚶摄食数量最多,2.5 cm实验组摄食数量最少,这与刘佳[28]研究认为多棘海盘车对于最小壳长组摄食量最大的结果相同。从本研究结果来看,壳长在2.0~2.5 cm的魁蚶个体作为底播苗种的标准较为适合,这也与唐启升等[9]研究结论相近。在开展底播增殖过程中,魁蚶规格越大越能有效防御敌害,因此日本放流魁蚶规格多为2~3 cm甚至5~6 cm[29]。但过大的规格必然延长中间培育时间,增加成本,并有可能因春季缺氧、夏季高温等不可控环境因素导致稚贝幼蚶大量死亡[9]。因此,魁蚶底播规格需兼顾如潜沙、敌害、成本等多方面因素来确定。
3.2多棘海盘车对魁蚶摄食活动的昼夜差异
多棘海盘车对不同规格魁蚶摄食数量均存在昼夜差异,这种与光照有关的昼夜摄食差异广泛存在于水生动物中[30]。多棘海盘车属于低等棘皮动物,视觉感觉器官不发达,仅在上皮间散布着许多菱形的有触觉器和化学感觉器功能的神经感觉细胞。在多棘海盘车管足处吸盘数目较多,主要起到嗅觉功能[27]。虽然多棘海盘车的视觉和嗅觉感觉器官系统并不发达,但在长期的演化过程中,逐渐对光照、饵料等生态因子适应,从而形成了昼夜摄食差异[19]。在本实验中,实验开始前24 h多棘海盘车对4种规格的魁蚶摄食数量较低,主要原因可能是多棘海盘车和其他水生生物一样需要一定时间来适应周围环境的改变。在实验24~30 h(第二日白天)和48~60 h(第三日白天),仅观察到多棘海盘车对1 cm魁蚶有少量摄食,其他规格魁蚶没有明显摄食现象。在实验30~48 h(第二日夜晚),1和1.5 cm实验组大量被摄食,空壳所占百分比分别由29.39%和7.69%上升至68.18%和53.85%;而2和2.5 cm实验组则在60~72 h(第三日夜晚)摄食数量较大,空壳所占百分比分别由47.62%和50%上升至85.71%和100%。从本实验结果看,在魁蚶底播增殖中,如底播规格较小的苗种(不大于1.5 cm),在底播后第二日夜晚多棘海盘车对魁蚶摄食数量较大;如底播规格稍大的苗种(1.5~2.5 cm),在底播后第三日夜晚多棘海盘车对魁蚶摄食数量较大。多棘海盘车对不同规格魁蚶摄食数量均存在一定的昼夜差异,夜间的摄食数量大于白天,魁蚶规格越大,多棘海盘车对魁蚶最大摄食数量时间出现的越晚。
3.3 魁蚶底播增殖中多棘海盘车防治策略
多棘海盘车是口外食者,可以依据气味对贝类进行精确定位。根据搜寻理论,捕食者通常会选择那些高收益性的饵料生物,而往往忽略低收益性的饵料生物[31]。Tamura认为浅水区贝类的闭壳肌力量比深水区的力量大[32],从饵料生物可获性来说,魁蚶具有比浅水贝类具有更强大的闭壳肌,打开它们需要耗费更多的能量。因此可以在魁蚶底播增殖区附近选取浅水低值贝类作为诱捕生物对多棘海盘车进行清除,从而减少多棘海盘车对魁蚶的摄食量,以获得较好的经济收益。
[1] 王兴章,邢信泽,张启刚. 几种理化因子对魁蚶人工育苗的影响[J]. 齐鲁渔业, 2003, 20(2): 17.
[2] 孙振兴. 魁蚶的增养殖[J]. 科学养鱼, 1990, 4: 166-169.
[3] 于瑞海, 李琪. 无公害魁蚶底播增养殖稳产新技术[J]. 海洋湖沼通报, 2009(3): 87-90.
[4] 付卓,郑国富. 魁蚶育苗与增养殖技术[J]. 中国水产, 2007(11): 49-50.
[5] Sloan N A. Aspects of the feeding biology of asteroids [J]. Oceanogr Mar Biol Ann Rev, 1980,18:57-124.
[6] Menge B A. Effects of feeding on the environment: Asteroidea[M].//Jangoux M, Lawrence J M Eds. Echinodern Nutrition F L. USA: CRC Press, 1982: 524-551.
[7] Brenchley G A. Mechanisms of spatial competitions in marine soft bottom communities [J]. J Exp Mari Biol Ecol, 1982, 60: 7-33.
[8] Barkai A, McQuaid C. Predator-prey role reversal in a marine benthic ecosystem[J]. Science, 1988, 242: 62-64.
[9] 唐启生, 邱显寅, 王俊, 等. 山东近海魁蚶资源增殖的研究[J]. 应用生态学报, 1994, 5(4): 396-402.
[10] 刘佳, 张秀梅. 多棘海盘车对太平洋牡蛎、紫贻贝、菲律宾蛤仔摄食选择性的研究[J]. 中国海洋大学学报(自然科学版), 2012, 42(7-8): 98-105.
[11] 周学家, 刘信艺, 王世信, 等. 山东近海海星种类及分布[J]. 齐鲁渔业, 1996, 13(1): 41-43.
[12] 杨德渐, 孙世春. 海洋无脊椎动物学[M].青岛: 青岛海洋大学出版社, 1999: 486-500.
[13] Hatanaka M, Kosaka M. Biological studies on the population of the starfish, Asterias amurensis, in Sendai Bay[J]. Tohoku Journal of Agricultural Research, 1958, 9(3): 159-177.
[14] Davenport S R. Mcloughlin R. Preliminary assessment of the distribution and potential impact of the introduced starfishAsteriasamurensisin Tasmanian waters: status report to Fisheries Research and Development Corporation[M]. Fisheries Research and Development Corporation(Australia). Hobart: CSIRO Division of Fisherues, 1993: 38.
[15] Barkai A, McQuaid C. Predator-prey role reversal in a marine benthic ecosystem [J].Science, 1988, 242: 62-64.
[16] Hawkes G, Day R. Review of the biology and ecology ofAsteriasamurensis:status report to Fisheries Research and Development Corporation[M].Fisheries Research and Development Corporation(Australia). Hobart: CSIRO Division of Fisherues, 1993: 38.
[17] Thompson M, Drolet D, Himmelman J H. Localization of infaunal prey by the sea starLeptasteriaspolaris[J]. Marine Biology, 2005, 146: 887-894.
[18] Kim Y S. Histological observation of the annual change in the gonad of the starfish,AsteriasamurensisLuken [J]. Bulletin of the Faculty of Fisheries Hokkaido University, 1968, 19(2): 97-108.
[19] 杜美荣, 方建光, 张继红, 等. 多棘海盘车对四种贝类摄食率和选择性的初步研究[J]. 渔业现代化, 2012, 39(2): 25-29.
[20] 李淑芸, 张秀梅, 聂猛, 等. 不同温度下多棘海盘车对菲律宾蛤仔的摄食选择性研究[J]. 水产学报, 2014, 38(7): 981-991.
[21] Himmelman J H, Dutil C, Gaymer C F. Foraging behavior and activity budgets of sea stars on a subtidal sediment bottom [J]. J Exp Mar Biol Ecol, 2005, 322: 153-165.
[22] Barbeau M A, Scheibling R E. Behavioral mechanisms of prey size selection by sea stars (AsteriasvulgarisVerrill) and carbs (CancerirroratusSay) preying on juvenile sea scallops (Placopectenmagellanicus(Gmelin)) [J]. J Exp Mar Biol Ecol, 1994, 180: 103-136.
[23] Laudien J, Wahl M. Indirect effects of epibiosis on host mortality: seastar predation on differently fouled mussels [J]. Mar Ecol, 1990,20(1): 35-47.
[24] Kim Y S. Selective feeding on the several bivalve mollusks by starfish, Asterias amurensis lüken [J]. Bull Fac Fish Hokkaido univ, 1969, 19(4):244-249.
[25] Penney A J, Griffiths C L. Prey selective and the impact of the starfish Marthasterias glacialis (L.) and other predators on the musselChoromytilusmeridionalis(Krauss) [J]. J Exp Mar Biol Ecol, 1984,75(1): 19-36.
[26] 尚玉昌. 行为生态学[M]. 北京: 北京大学出版社, 1998: 20-22.
[27] Turner E. A northern Pacific seastar,Asteriasamurensisin Tasmania[J]. AMSA Bulletin, 1992, 120(9): 18-19.
[28] 刘佳. 多棘海盘车摄食行为和繁殖生物学的初步研究[D]. 青岛: 中国海洋大学, 2012: 47-50.
[29] 高见东洋. 魁蚶增殖研究[J]. 水产养殖, 1981, 29(1): 38-56.
[30] 张海发, 刘晓春, 林浩然, 等. 斜带石斑鱼仔鱼的摄食节律及日摄食量[J]. 水产学报, 2004, 28(6): 669-674.
[31] Elner R W, Hughes R N. Energy maximization in the diet of the shore crabs,Carcinusmaenas[J]. J Anim Ecol, 1978, 47(1): 103-107.
[32] Tamura T. The power of the adductor muscle of the oyster, Ostrea circunpicta Pils [J]. Sci Rep Tohoku Imp Univ, 1929, 4(4): 259-279.
责任编辑 朱宝象
A Preliminary Study on Food Consumption, Preference and Day-Night Predatory Differential ofAsteriasamurensisonScapharcabroughtonii
ZHANG Tian-Wen, LIU Guang-Bin, LIU En-Fu, ZHENG Yong-Yun, QIU Zhao-Xing
(Marine Biology Institute of Shandong Province,Qingdao 266002,China)
In order to study the food consumption, preference and day-night predatory differential ofAsteriasamurensisonScapharcabroughtonii, the experiments were carried out at temperatures ranging from 16 to 20 ℃, salinity 28.2±2.1, dissolved oxygen (7.5±0.7) mg/L and pH 8.0±0.3.The results provided basic data for promoting steady and healthy development of the multiplication and cultivation ofS.broughtonii. The results of food consumption suggested that the food intake of starfish increased with the density ofS.broughtoniiat all conditions. The food consumption of starfish in different sizes onS.broughtoniiin the same size had no significant difference(P>0.05), but significantly different amongS.broughtoniiin different sizes. The starfish had the highest feeding rate on the smallestS.broughtonii. Our findings also verified the positive relation between the weight of food intake of starfish in different sizes on the density and size ofS.broughtonii. The feeding rhythm of starfish onS.broughtoniiin different sizes showed a certain day-night differential, higher in night than on day. The maximum food consumption occurred when the size ofS.broughtoniibecame lager. The results showed that 30 ind/m2was suitable sea bottom sowing density forS.broughtonii. Our data indicated that the ability to avoid predators and rivals increased as the shell length raise. Based on prey selectivity bivalves that have a lower commercial value may be used to clear awayA.amurensis.
Asteriasamurensis;Scapharcabroughtonii; food consumption; food preference; day-night differential
国家海洋公益性项目(201205023);现代农业产业技术体系建设专项资金项目(CARS-48);山东省博士基金项目(BS2010HZ017); 山东省现代农业产业技术体系建设专项资金项目(SDAIT-19)资助
2014-03-17;
2015-06-08
张天文(1983-),男,博士,主要从事养殖生态学研究。E-mail:ztw236@163.com
❋❋通讯作者: E-mail: zhaoxingqiu@163.com
S912; S917.1
A
1672-5174(2015)12-024-06
10.16441/j.cnki.hdxb.20140089
