猪H-FABP基因的克隆及真核表达载体的构建
2015-03-30戚顺民吴明明孙金海
戚顺民,马 康,吴明明,孙金海*
(1.青岛农业大学动物科技学院,山东青岛266109;2.中国农业大学,北京100193)
猪H-FABP基因的克隆及真核表达载体的构建
戚顺民1,马 康1,吴明明2,孙金海1*
(1.青岛农业大学动物科技学院,山东青岛266109;2.中国农业大学,北京100193)
心肌脂肪酸结合蛋白基因(H-FABP)是影响家猪肌内脂肪含量的重要候选基因之一。本研究以长白猪为实验材料,采用RT-PCR方法克隆H-FABP基因全长编码序列,并构建pEGFP-N1-H-FABP融合表达载体,通过脂质体介导转染PK15细胞,48 h后进行荧光表达检测。获得了猪H-FABP基因去终止密码子全长编码序列399 bp,成功构建pEGFP-N1-H-FABP融合表达载体,瞬时转染后在PK15细胞中顺利表达。本实验为进一步从细胞水平上解析及验证该基因的生物学功能奠定了基础。
H-FABP;真核表达;融合表达;长白猪
肌内脂肪(IMF),俗称大理石花纹,是影响猪肉质评定的一个重要指标[1]。尽管大多数肉质性状的遗传力较低,但IMF含量却有着较高的遗传力[2],表明其可以通过育种获得较大的遗传改良。研究认为,IMF含量在2.5%~3%的范围内猪肉品质较好,低于1.5%会明显降低猪肉品质[3]。但目前对高瘦肉率和低背膘厚猪种的遗传选育,使肌肉中IMF含量已降低至1%~1.5%。因此,如何在保证一定瘦肉率和低背膘厚的前提下,提高IMF含量成为目前商品猪品种改良的新目标。
脂肪酸结合蛋白(FABPs)家族是一组广泛存在于许多细胞类型胞质内的小分子蛋白,其中与猪肉质密切相关的FABPs有2种类型:脂肪型脂肪酸结合蛋白 (A-FABP)和心脏型脂肪酸结合蛋白 (HFABP)。H-FABP主要通过参与细胞脂质转运过程来实现其生物学功能,其能与胞质内的脂肪酸结合,进而促进细胞对脂肪酸的吸收[4-6]。Gerbens等[7]研究表明,H-FABP基因的mRNA表达水平与猪IMF含量之间存在显著正相关关系。近年来,诸多研究表明H-FABP基因多态性对猪IMF含量影响显著[8-10]。
本研究以长白猪为试验材料,采用基因克隆并构建真核表达载体的方法,对长白猪H-FABP基因编码序列进行分析并构建H-FABP基因融合表达载体,为进一步研究猪H-FABP基因的生物学功能及改善传统瘦肉型猪的肉质奠定理论基础。
1 材料与方法
1.1 材料 长白猪耳组织取自青岛农业大学试验基地,大肠杆菌DH5α、载体pEGFP-N1以及PK15细胞由本实验室保存。
1.2 所需主要仪器设备 PCR仪,电泳仪,凝胶自动成像系统,超净工作台,CO2培养箱,倒置荧光显微镜,水浴恒温振荡器,常规手术器械及玻璃器皿。
1.3 主要试剂 反转录试剂盒购于美国Thermo公司;RNAiso Plus、DNase I、Ex Tap DNA聚合酶、DNA限制性内切酶Hind III和BamH I、T4 DNA连接酶和凝胶回收试剂盒购于宝生物(大连)公司;无内毒素质粒大量抽提试剂盒购于天根生化科技(北京)公司;胎牛血清、DMEM细胞培养液购于美国Hyclone公司;卡那霉素、青霉素购于德国SIGMA公司;脂质体2000购于Invitrogen公司。
1.4 H-FABP基因的克隆
1.4.1 引物设计与合成 参照GenBank数据库中已有的猪H-FABP基因编码区序列 (登录号: NM_001099931.1),应用Primer 5.0软件设计1对扩增H-FABP基因cDNA完整编码序列的引物,保留H-FABP基因cDNA的起始密码子,去掉终止密码子,并引入酶切位点(正向引物:5′-CCCAAGCTTATGGTG GACGCCTTCGC-3′,下划线序列为Hind III酶切位点;反向引物:5′-CGGGATCCTGCCTCTTTCTCGTAA GTGCG-3′,下划线部分为BamH I酶切位点),扩增目的片段大小为399 bp,引物由上海生工生物工程有限公司合成。
1.4.2 H-FABP基因的扩增与克隆 采用Trizol法抽提获得长白猪耳组织总RNA,反转录获得cDNA, RT-RCR方法扩增得到H-FABP基因全长编码序列并用1.5%琼脂糖凝胶电泳进行检测。回收纯化后与pMD19-T载体连接并转化大肠杆菌DH5,在含氨苄抗性的固体培养基上37℃恒温培养12~16 h,挑取单克隆于液体培养基中过夜培养,采用菌液PCR和双酶切进行插入片段的鉴定,阳性克隆送上海生工生物工程有限公司测序。
1.4.3 H-FABP基因序列测定及分析 在NCBI网站上利用在线工具BLAST对克隆得到的H-FABP基因编码序列与GenBank上的6种动物:人(NM_004102.3)、大 鼠 (NM_024162.1)、鸡 (NM_001030889.1)、牛(NM_174313.2)、山 羊 (NM_001285701.1) 及 驴(GQ244482.1)的相应编码区序列和氨基酸序列进行同源性比对。
1.5 融合表达载体构建、转染、筛选及鉴定
1.5.1 融合表达载体构建 将测序鉴定正确的pMD19-T-H-FABP质粒和pEGFP-N1质粒同时进行Hind III和BamH I双酶切,胶回收H-FABP基因和pEGFP-N1载体片段,用T4连接酶16℃连接6 h,转化DH5,在含kan+抗性的固体培养基上37℃恒温培养12~16 h,挑取阳性克隆进行菌液PCR、双酶切鉴定,将鉴定正确的重组质粒命名为pEGFP-N1-HFABP,并送上海生工生物工程有限公司测序。
1.5.2 融合表达载体瞬时转染PK15细胞 采用脂质体介导将鉴定正确的H-FABP基因真核表达载体pEGFP-N1-H-FABP瞬时转染PK15细胞。试验组:转染重组质粒pEGFP-N1-H-FABP;阴性对照组:转染空载体质粒pEGFP-N1;空白对照组:未转染质粒。转染48 h之后于荧光倒置显微镜下观察,200倍放大倍数下每组随机选取10个视野的转染的细胞数和细胞总数分别进行计数,以10个视野的平均百分比表示每组PK15细胞的平均转染效率。
1.5.3 统计分析 用于分析统计的软件为SPSS V19.0,选用卡方检验进行统计学处理。计量数据均以平均值±标准差表示。
2 实验结果
2.1 长白猪耳组织总RNA提取 Trizol法提取长白猪耳组织总RNA,2%琼脂糖凝胶电泳检测RNA完整性,显示有清晰的28 S和18 S条带(图1)。说明提取的总RNA完整度较高,可进行后续反转录。
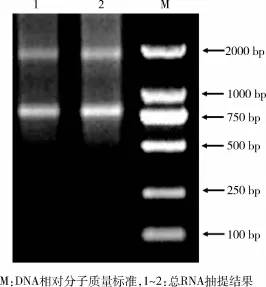
图1 总RNA琼脂糖凝胶电泳检测
2.2 长白猪H-FABP基因克隆及测序 通过反转录PCR扩增得到目的基因H-FABP,1.5%琼脂糖凝胶电泳显示在399 bp处得到1条清晰的条带(图2)。测序结果表明所获得的猪H-FABP与GenBank(登录号: NM_001099931.1)中的相应基因序列完全一致,证明RT-PCR扩增获得的目的基因为实验所需目的基因。将目的片段回收与pMD19-T载体连接得到质粒命名为pMD19-T-H-FABP。质粒经Hind III和BamH I双酶切8 h后,在399 bp和2 700 bp处显示有条清晰的条带(图3),其中399 bp为目的基因片段大小,2 700 bp为载体长度大小,与预期结果相符。
2.3 H-FABP基因生物信息学分析 将测序获得的长白猪H-FABP基因编码区序列与GenBank中收录的人(NM_004102.3)、大鼠(NM_024162.1)、鸡(NM_00 1030889.1)、牛(NM_174313.2)、山羊(NM_001285701.1)及驴(GQ244482.1)相应编码区序列和氨基酸序列进行同源性比对(表1)。发现在哺乳动物中,该基因的同源性达86%~94%,氨基酸的同源性达88%~95%,可知H-FABP基因在不同物种之间具有较高的保守性,推断该基因在物种遗传进化过程中具有重要作用。
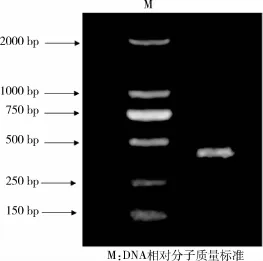
图2 H-FABP基因克隆琼脂糖凝胶电泳结果
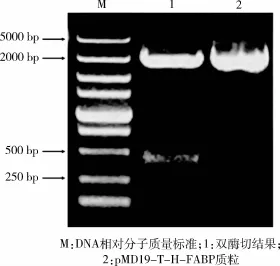
图3 pMD19-T-H-FABP质粒双酶切结果

表1 H-FABP基因在长白猪和其他物种之间的同源性比对
2.4 pEGFP-N1-H-FABP融合表达载体的构建与鉴定 将构建好的融合表达载体进行转化大肠杆菌DH5,挑取阳性克隆,进行菌液PCR扩增鉴定(图4)。融合表达载体经Hind III和BamH I双酶切8 h后,在399 bp和4 700 bp处显示出2条清晰的条带(图5),其中399 bp为目的基因的片段大小,4 700 bp为载体的长度大小,与预期结果相符,测序结果证实所获得的目的片段与GenBank中的H-FABP基因序列完全一致,说明pEGFP-N1-H-FABP融合表达载体构建正确,融合表达载体图谱见图6。
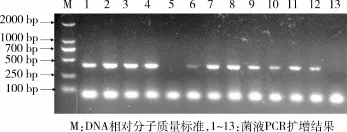
图4 菌液PCR琼脂糖凝胶电泳结果

图5 pEGFP-N1-H-FABP融合表达载体双酶切结果

图6 pEGFP-N1-H-FABP融合表达载体图谱
2.5 pEGFP-N1-H-FABP融合表达载体瞬时转染PK15细胞结果 将pEGFP-N1-H-FABP融合表达载体瞬时转染PK15细胞,48 h后置于荧光显微镜下观察绿色荧光蛋白表达情况,转染pEGFP-N1-HFABP融合表达载体的PK15细胞可以观察到绿色荧光(图7),转染效率平均为21.28%;转染空载体的PK15细胞,绿色荧光蛋白高表达(图8),转染效率平均为40.36%;未转染质粒的细胞未观察到绿色荧光(图略)。
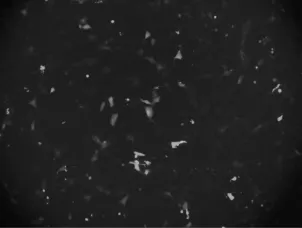
图7 融合表达载体转染PK15细胞荧光结果图

图8 空载体转染PK15细胞荧光结果图
3 讨论
生长性能作为猪育种中最重要的经济性状之一,发现并验证新的候选基因对家猪的遗传改良有着极其重要的意义。相关研究表明,H-FABP基因可作为肌内脂肪含量的重要候选基因[11]。H-FABP基因表达较为广泛,在心脏、骨骼肌、肾脏、肺脏、乳腺、大脑、胎盘以及卵巢等部位均有表达,但表达量差异较大,主要在心肌细胞和骨骼肌细胞内表达。本研究从长白猪耳组织中成功克隆其H-FABP基因编码序列,也从侧面印证了该基因表达较为广泛的特点。通过其与GenBank中的人、大鼠、鸡、牛、山羊及驴进行同源性比较,笔者发现在哺乳动物中,该基因序列的同源性达86%~94%,编码氨基酸的同源性达88%~95%,可见H-FABP基因具有较高的保守性且氨基酸突变类型多属于同义突变,也显示出H-FABP基因在动物遗传进化过程中对于维护机体正常生理机能具有重要意义。
本研究选用的pEGFP-N1载体中含有SV40 ori和PCMV复制原件,可在宿主细胞分裂的过程中,保证目的基因的稳定表达。GFP(绿色荧光蛋白)是一种生物发光物质,由238个氨基酸残基组成,EGFP(增强型绿色荧光蛋白)为GFP的突变体,在细胞内可稳定表达绿色荧光,其荧光强弱取决于蛋白的含量, GFP与外源基因偶联时一般不影响外源蛋白的构象和功能[12-13]。本研究发现,融合表达载体转染PK15细胞的转染效率(21.28%)显著低于空载体转染PK15细胞的转染效率(40.36%)。推测,外源基因与EGFP基因融合表达可能会改变GFP的空间构象,进而影响绿色荧光的表达强度。
目前,国内对于猪H-FABP基因的研究大多停留在该基因的多态性与性状的关联分析方面。本研究以PK15细胞为宿主细胞,通过脂质体介导转染带有目的基因和绿色荧光蛋白的融合表达载体,探讨了H-FABP基因在细胞中的表达,为进一步从细胞水平上解析及验证该基因的生物学功能奠定基础。
[1]Fiedler I,Nürnberg K,Hardge T,et al.Phenotypic variations of muscle fibre and intramuscular fat traits in longissimus muscle of F2 population Duroc×Berlin Miniature Pig and relationships to meat quality[J].Meat Sci,2003,63(1):131-139.
[2] Suzuki K,Irie M,Kadowaki H,et al.Genetic parameter estimates of meat quality traits in Duroc pigs selected for average daily gain,longissimus muscle area,backfat thickness, and intramuscular fat content[J].J Anim Sci,2005,83(9):2058-2065.
[3] Devol D L,Mckeith F K,Bechtel P J,et al.Variation in composition and palatability traits and relationships between muscle characteristics and palatability in random sample of pork carcasses[J].J Anim Sci,1988,66(2):385-395.
[4]Haunerland N H.Fatty Acid Binding Proteins[M].Encyclopedia of Entomology:Springer Verlag New YorkInc,2008:1416-1418.
[5] 徐海霞,房兴堂,陈宏,等.脂肪酸结合蛋白及其主要家族基因的研究进展[J].畜牧与兽医,2008,40(1):94-97.
[6]陈轶霞,骆学农,王红宁.脂肪酸结合蛋白及其生物学功能[J].生命的化学,2012,32(6):502-506.
[7]Gerbens F,Verburg F J,Van Moerkerk H T,et al.Associations ofheartand adipocyte fatty acid-binding protein gene expression with intramuscular fat content in pigs[J].J Anim Sci, 2001,79(2):347-354.
[8] 呼红梅,王继英,郭建凤,等.莱芜猪和杜洛克猪肌肉HFABP基因表达量与肌内脂肪和脂肪酸含量关联分析[J].华北农学报,2010,25(4):64-68.
[9] Cho K H,Kim M J,Jeon G J,et al.Association of genetic variants for FABP3 gene with back fatthickness and intramuscular fat content in pig[J].Mol Biol Rep,2011,38(3):2161-2166.
[10]Chen J N,Jiang Y Z,Cen W M,et al.Distribution of H-FABP and ACSL4 gene polymorphisms and their associations with intramuscular fat content and backfat thickness in different pig populations[J].Genet Mol Res,2014,13(3):6759-6772.
[11]Arnyasi M,Grindflek E,Jávor A,et al.Investigation of two candidate genes for meat quality traits in a quantitative trait locus region on SSC6:the porcine short heterodimer partner and heart fatty acid binding protein genes[J].J Anim Breed Genet, 2006,123(3):198-203.
[12]程根阳,陈香美,白雪源,等.绿色荧光蛋白与人肾脏NaDC3融合基因的构建及其在肾小管上皮细胞中的定位[J].细胞生物学杂志,2003,25(3):170-173.
[13]Li Y,Li J,Lu C R,et al.The construction and identification of eukaryotic expression vector pEGFP-N1-hTERT[J].Agri Sci Tech, 2008,9(5):50-54.
Cloning and Constructing of Eukaryotic Expression Vector of H-FABP Gene in Pig
QI Shun-min1,MA Kang1,WU Ming-ming2,SUN Jin-hai1*
(1.College of Animal Science and Technology,Qingdao Agricultural University,Shandong Qingdao 266109,China; 2.China Agricultural Uniuersity,Beijing 100193,China)
Heart fatty acid binding protein gene(H-FABP)is one of the most important candidate genes that affect pig’s intramuscular fat content.In this research,the full-length coding sequence of H-FABP gene was cloned from Landrace by RT-PCR method.pEGFP-N1-H-FABP fusion expression vector was constructed and detected by liposome-mediated transfection PK15 cells with fluorescence expression after 48 h.The whole CDS size of H-FABP was 399 bp excluding the termination codon.The pEGFP-N1-H-FABP fusion expression vector was constructed and successfully expressed in PK15 cells.The results of this trial provide a basis for parsing and verifying the biological function of this gene on cellular level.
H-FABP;eukaryotic expression;fusion expression;landrace
S828.2
A文献标识码:0258-7033(2015)11-0069-05
2014-09-29;
2015-01-21
转基因生物新品种培育科技重大专项(2013ZX08006-003);山东省现代农业产业技术体系生猪创新团队建设项目(SDAIT-06-022-01)
戚顺民(1989-),男,在读硕士,E-mail:1030308369@qq. com
*通讯作者:孙金海,博士,教授,主要从事细胞遗传和分子遗传学研究,E-mail:sunjinhai00528@sina.com.
