水分胁迫对香樟和黑壳楠幼苗光合特性的影响
2015-03-23张佑麟徐振东费永俊
丁 琼 张佑麟 刘 刚 徐振东 费永俊
(长江大学楠木种质资源评价与创新中心,湖北 荆州 434025)
水分胁迫对香樟和黑壳楠幼苗光合特性的影响
丁 琼 张佑麟 刘 刚 徐振东 费永俊
(长江大学楠木种质资源评价与创新中心,湖北 荆州 434025)
测定2年生香樟和黑壳楠实生苗在不同水分环境下的光合指标变化,分析其光合生理响应机制。结果表明:淹水和干旱胁迫下,黑壳楠和香樟幼苗的叶绿素(Chl)、净光合速率(Pn)均呈现先升高后降低的趋势;蒸腾速率(Tr)随胁迫时间延长呈下降趋势;气孔导度(Gs)呈现逐步上升趋势,且在胁迫后期气孔导度的增幅较大。与香樟相比,黑壳楠对水淹和干旱胁迫的反应更灵敏,且耐受时间更持久,表现出较强的耐涝性和抗旱性。
水淹胁迫;干旱胁迫;香樟;黑壳楠;光合特性
黑壳楠(Linderamegaphylla)别名八角香,樟科(Lauraceae)山胡椒属常绿乔木,起源于晚白垩纪,原产于中国云、贵、川和两湖、两广等地,现分散到很多地区[1]。其叶大色亮、树形优美,是集观赏、生态和经济于一体的庭院观赏和园林绿化树种[2]。香樟(Cinnamomumcamphora)别名猴樟,樟科樟属常绿乔木,分布于湖南、湖北、云南、广东、广西、台湾、福建、江西等省区[3]。其具有很强的吸烟滞尘、涵养水源、固土防沙和美化环境的能力,此外,抗海潮风及耐烟尘和抗有毒气体能力也较强,并能吸收多种有毒气体,较能适应城市环境,是优良的园林绿化树种。
国内学者对黑壳楠和香樟已开展了大量研究工作。黑壳楠方面的研究包括生理特性[4]、光合特性[5]等;香樟方面的研究包括抗寒特征[6]、生物学特征[7]、生理特性[8]等,但对于黑壳楠和香樟的耐淹性和抗旱性,以及其对水淹和干旱的响应机制研究还较少。目前,在植物的水分胁迫研究中,水涝和干旱各有侧重,北方主要偏重植物耐旱机制的研究,而南方及沿海地区主要偏重耐水涝机制的研究,但对从干旱到水涝不同水分状况下植物的生理生态反应的系统研究较少。另外,河道树种因季节变化和水位消长,遭受水涝和干旱双重胁迫。因此,本研究以2种樟科植物(黑壳楠和香樟)的2年生幼苗为研究材料,通过双套盆控水法研究其幼苗在受到淹水胁迫及其在持续干旱条件下的生理变化规律, 并且对黑壳楠和香樟幼苗在淹水和干旱胁迫下的抗性指标进行分析比较,为筛选抗涝和抗旱型绿化植被及其广泛应用提供依据。
1 试验地概况
试验地位于湖北省荆州市长江大学内,地处东经111°15′~114°05′,北纬29°26′~31°31′。属亚热带季风湿润气候区,年辐射总量4 366.8~4 576.2 MJ/m2,年日照时数1 823~1 978 h,日照率为41%~44%。年均气温16.2~16.6 ℃,无霜期250~267 d,年降水量1 100~1 300 mm。本试验在长江大学园林园艺学院实习基地的塑料大棚中进行,其中,2011年11月至2012年1月大棚内温度为20~28 ℃,空气湿度50%~65%。
2 材料与方法
2.1 试验材料与处理
将2年生黑壳楠、香樟实生苗栽植于直径25 cm,高21 cm的瓦盆中,每盆1株,基质为沙子、锯沫、陶粒、珍珠岩按照比例4∶2∶2∶1进行配置的混合基质。将长势均匀的黑壳楠、香樟2年生盆栽幼苗各100株移进塑料大棚中,并进行常规管理。挑选长势均匀一致的黑壳楠、香樟2年生幼苗90株进行胁迫处理,采用完全随机设计,每树种每处理设3个重复。每处理各15盆,将处理分为5个梯度水平,分别为全淹、2/3淹、1/3淹、对照(CK)和持续干旱。
1) 全淹水平:土壤含水量饱和,采用双套盆法,保持水面与瓦盆水平,保持土壤含水量处于完全饱和状态,模拟自然状态下的水渍危害。
2) 2/3淹水平:水面处于盆子的2/3处,及水面在土壤表面的14 cm,模拟地下水位过高,为轻度淹水胁迫。
3) 1/3淹水平:含水量处于盆子的1/3处。
4) 对照水平:土壤含水量为最大田间持水量的75%左右,这是木本植物生长的最佳含水量,在本试验中作为对照。
5) 持续干旱水平:40%~45%饱和田间持水量,为中度干旱胁迫。其中每天查看1次盆中水分情况,用量筒及时补充水分到指定胁迫程度。于11月18日、11月28日、12月8日、12月18日采取叶片进行光合指标的测定。
2.2 测定指标与方法
2.2.1 光合特性的测定 对不同水分处理的黑壳楠和香樟进行测定。其净光合速率(Pn)、气孔导度(Gs)和蒸腾速率(Tr)选择无云天气,在9:00—11:30,每株测定2片叶子,每次测定叶片尽量保持同一位置,测定叶片为黑壳楠与香樟幼苗顶端两片对称叶。用美国Li-6400便携式光合分析仪测定。
2.2.2 叶绿素含量的测定 测定2树种叶片的叶绿素a(Chla)、叶绿素b(Chlb)和叶绿素总含量(Chla+b),选取用于测定光合速率的叶片,采用丙酮-乙醇法[9]测定。
2.2.3 模糊隶属值计算 评价方法参照侯建华等[10]的方法进行。
2.3 数据分析
采用Excel 2003进行数据整理和绘图,采用Duncan多重比较检验每个生理指标处理间的差异显著性。
3 结果与分析
3.1 水分胁迫对黑壳楠和香樟幼苗光合特性的影响
3.1.1 净光合速率 淹水和干旱处理后黑壳楠和香樟净光合速率的变化情况见图1。
水分胁迫能极其显著地影响黑壳楠和香樟幼苗的净光合速率。在淹水和干旱胁迫后,随着处理时间的延长,黑壳楠和香樟的净光合速率总体均呈先升高后下降的趋势,但香樟下降的时间更为提前,干旱胁迫后,净光合速率始终呈下降趋势(图1)。多重比较显示,淹水胁迫下黑壳楠净光合速率显著低于香樟,全淹处理、2/3水淹处理的净光合速率显著低于1/3水淹处理, 1/3水淹处理又显著低于对照。说明全淹和2/3水淹对2个树种的净光合速率影响均较大。随着胁迫程度的增大,净光合速率降低的幅度也越来越大,在同一淹水胁迫程度下,黑壳楠净光合速率受影响要比香樟大(表1)。持续干旱胁迫下,黑壳楠净光合速率显著低于香樟,干旱处理也显著低于对照,随着胁迫时间的延长,净光合速率有明显的下降趋势,说明黑壳楠在受干旱胁迫后光合作用可能受胁迫影响的程度更大(表2)。
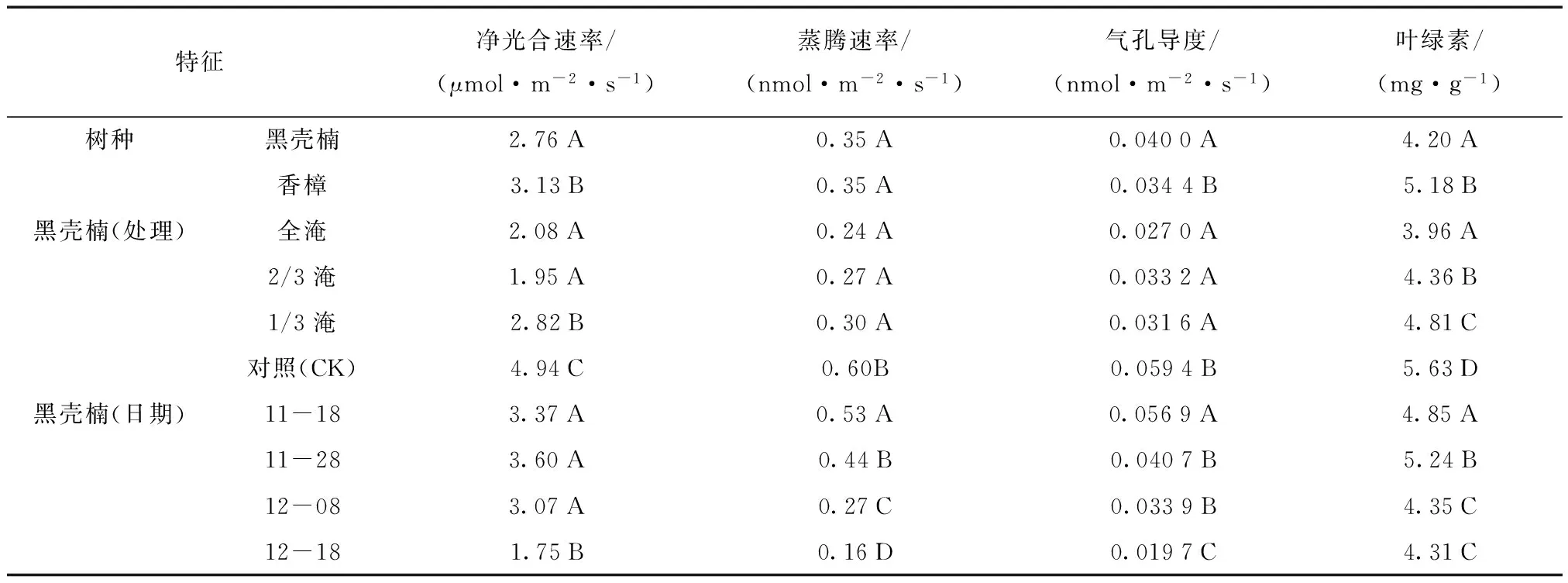
表1 淹水胁迫下光合指标的多重比较Tab.1 Multiple comparison of photosynthetic characteristics under water logging stress
注:同一列中含有不同字母表示差异显著,相同字母表示差异不显著(P<0.05)。
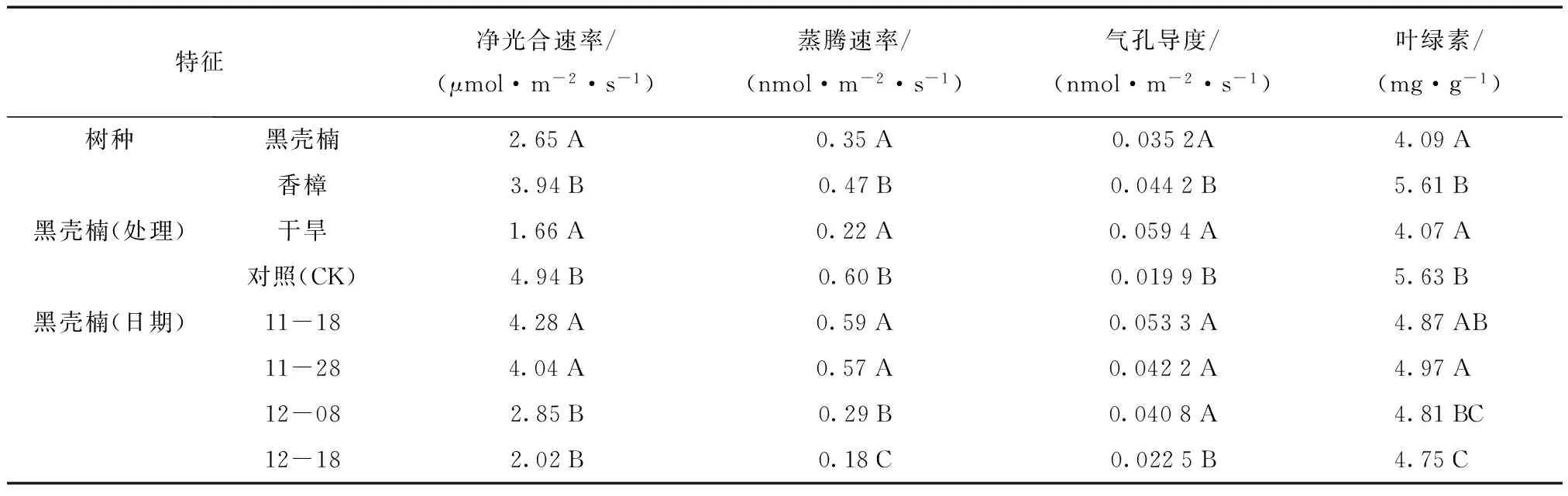
表2 干旱胁迫下光合指标的多重比较Tab.2 Multiple comparison of photosynthetic characteristics indices under drought stress
注:同一列中含有不同字母表示差异显著,相同字母表示差异不显著(P<0.05)。
3.1.2 蒸腾速率 水分胁迫也能极其显著地影响黑壳楠和香樟幼苗的蒸腾速率。随胁迫时间延长,黑壳楠和香樟的蒸腾速率均呈现下降趋势,其中黑壳楠在受到干旱胁迫后蒸腾速率呈先上升后下降的趋势,这可能是黑壳楠在受到干旱胁迫后有一定自我保护机制(图2)。淹水胁迫下黑壳楠与香樟的蒸腾速率差异不显著,全淹处理、2/3水淹、1/3水淹处理之间未形成显著差异,但比对照(CK)低,说明淹水后对2个树种均产生影响。随着胁迫程度的增大,黑壳楠蒸腾速率降低的幅度不大(表1);在同一淹水胁迫程度下,香樟蒸腾速率受影响程度更大(图2)。持续干旱胁迫下,黑壳楠蒸腾速率显著低于香樟,处理蒸腾速率比对照(CK)显著低,说明持续干旱胁迫对两个树种的蒸腾速率均产生较大影响。随着胁迫时间的延长,蒸腾速率呈显著下降的趋势(表2)。

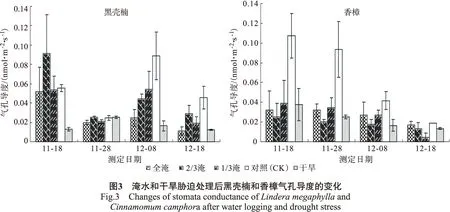
3.1.3 气孔导度 水分胁迫也能极其显著地影响黑壳楠和香樟幼苗的气孔导度。淹水和干旱胁迫下,黑壳楠气孔导度随着时间变化,呈先下降后上升又下降的趋势,而香樟气孔导度总体上呈下降趋势(图3)。黑壳楠在淹水胁迫下气孔导度显著高于香樟,全淹处理、2/3水淹处理、1/3水淹处理之间未形成显著差异,但是比对照(CK)低,说明淹水后对2个树种的气孔导度均产生影响,香樟受到的影响程度更大(表1)。在受干旱胁迫影响后,黑壳楠的气孔导度显著低于香樟,干旱处理比对照(CK)显著高,说明持续干旱胁迫下黑壳楠幼苗呈现一定的应对机制。随着胁迫时间的延长,气孔导度降低的幅度不大(表2)。
3.1.4 叶绿素含量 黑壳楠和香樟幼苗在叶绿素(Chl)含量上仍然受到水分胁迫的显著影响。淹水和干旱胁迫下,黑壳楠叶绿素含量表现出随时间变化上升—下降的趋势;同一时间段,淹水处理后植株叶绿素含量表现出对照(CK)≥1/3水淹≥2/3水淹≥全淹;干旱胁迫下,叶绿素含量对照(CK)≥干旱,这可能反射出黑壳楠在受淹水和干旱胁迫后,由于其叶绿素含量的下降导致其光合作用能力的下降。在淹水处理下,香樟叶绿素含量随时间延长表现出先上升后下降的趋势,且随着淹水时间的延长,叶绿素下降幅度增大;除11月28日外,其余测定时间香樟叶绿素含量均表现为对照组(CK)≥1/3水淹≥2/3水淹≥全淹,说明淹水处理对香樟叶绿素含量影响较大。持续干旱胁迫下叶绿素含量<对照(图4)。黑壳楠在受到淹水胁迫后,其叶绿素含量显著低于香樟;各淹水处理间表现出1/3水淹≥2/3水淹≥全淹,且差异显著,均显著低于对照。说明淹水胁迫对黑壳楠和香樟均有较大影响。随着胁迫时间的延长,黑壳楠叶绿素含量表现出先上升后下降的趋势,这可能与植物自我调节机制有关(表1)。在受到持续干旱胁迫后,黑壳楠叶绿素含量显著低于香樟,干旱处理也显著低于对照(CK),说明在受到淹水与干旱胁迫后,2树种的叶绿素含量随时间的变化呈现出升降趋势(表2)。

3.2 黑壳楠和香樟幼苗抗性能力综合评价
抗性综合评价就是按树种对逆境抵抗能力的大小进行筛选、评价和归类的过程,通过抗性鉴定,能很好地掌握树种特性,从而应用于生产。由于植物对水分胁迫的抗性机制比较复杂,单个生理指标很难评价其抗性能力的强弱,因此,必须综合分析植物光合作用的变化,才能科学评价其抗涝性和抗旱性的强弱。
对各项指标的隶属度值和综合评定结果表明:在全淹、2/3水淹、1/3水淹处理水平下,黑壳楠抗涝能力强于香樟;在干旱处理水平下,黑壳楠的抗旱能力也强于香樟(表3~4)。这个结果与试验过程中所观察的结果大致吻合,黑壳楠在受到淹水胁迫后表现出生长加快,但香樟则基本上保持不变,甚至到胁迫后期株型矮小。

表3 黑壳楠和香樟抗涝性综合评价Tab.3 The comprehensive appraisal on ability to water logging-resistant of Lindera megaphylla and Cinnamomum camphora
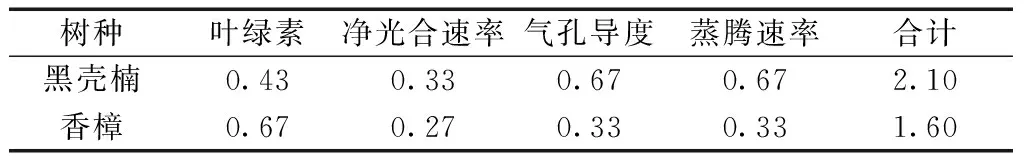
表4 黑壳楠和香樟抗旱性综合评价Tab.4 The comprehensive appraisal on ability to drought-resistant of Lindera megaphylla and Cinnamomum camphora
4 结论与讨论
在不同水分胁迫条件下,植物的净光合速率可以作为检验应试树种对水分逆境条件光合生理生态响应的一个重要核心表征因子[11-13],许多植物在遭受水淹后会伴随着净光合速率的下降[14],部分研究表明,植物在遭受水淹的几小时到几天内光合作用会明显降低[15],但随着时间的延长,耐淹植物会产生一定的适应性,逐渐恢复到正常水平或达到相对稳定的状态[16],但这种恢复能力也随着植物种类和水淹时间的变化而有所不同[17]。在本试验中,黑壳楠与香樟幼苗的净光合速率随着胁迫时间延长呈现出先上升后下降的趋势,这与衣英华等对枫杨(Pterocaryastenoptera)幼苗进行水分胁迫的研究结果相似[18]。在同一时间内,净光合速率表现出对照(CK)≥1/3水淹≥2/3水淹≥全淹≥干旱,净光合速率在全淹和持续干旱处理下受到的影响最大。
蒸腾作用对植物体具有十分重要的意义。在诸多环境因素中,基质含水量对植物蒸腾速率的影响远比气候因素直接。在本试验中,黑壳楠和香樟的蒸腾速率随胁迫时间延长均呈现下降趋势,但黑壳楠的蒸腾速率随着时间延长表现出先上升后下降的趋势,这说明淹水和干旱胁迫对黑壳楠和香樟幼苗均有较大影响,其中,香樟受影响程度更为明显。
已有研究表明,植物和土壤之间的水力导度下降将与根系缺氧一样,引起植物净光合速率和气孔导度降低[19-20]。在整个试验过程中,黑壳楠和香樟幼苗叶片气孔导度呈现出逐步上升趋势,并且在胁迫后期气孔导度的增幅较大,说明水淹和干旱处理对气孔导度的影响较为突出。
叶片中光合色素是叶片光合作用的物质基础,Chl含量与光合强度密切相关[21]。Chl之间的比值和总量在一定程度上表示了光系统对光量子的捕获和利用状况,同样也反应了植物处于特定环境下所承受光胁迫的潜在能力[22]。在本试验中,随着淹水处理时间的延长,2个树种的叶绿素总含量基本上表现出先上升后下降的趋势,在干旱胁迫下,2个树种的叶绿素总含量基本上也呈现出先上升后下降的变化趋势,但后期下降的程度较小,与张克清的研究一致[23],在同一时间,各处理水平下2树种的叶绿素总含量基本上低于对照水平,表明不同水分胁迫影响了植物叶片光合色素的合成,淹水胁迫下更为明显。
植物的耐涝性和抗旱性是由多因素控制的数量性状,与多种物质及复杂的生理生化反应有关,因此,单一的生理指标难以全面准确地反应不同树种耐涝性和抗旱性的强弱,应结合与耐涝性和抗旱性有关的多个指标来进行综合性评价。本研究计算了香樟和黑壳楠幼苗在受淹水和干旱胁迫后的平均隶属度值和综合排序值,在不同水分胁迫下,黑壳楠的所有隶属函数值均大于香樟,从而得出黑壳楠的耐涝和抗旱能力强于香樟。因此,黑壳楠更适合大面积地进行推广应用。
[1] 中国科学院植物志编辑委员会.中国植物志[M].北京:科学出版社,1979:8-10.
[2] 江明艳,陈其兵,潘远志.我国樟科植物的园林应用前景[J].西南园艺,2004,32(3):16-18.
[4] 滕开琼,杨秋生,戴钢,等.低温胁迫对黑壳楠幼苗生理效应及脂质过氧化作用的影响[J].河南农业大学学报,2002,36(3):151-154.
[5] 宋国,费永俊.不同配方施肥对黑壳楠生长和光合生理的影响[J].长江大学学报,2013,10(11):16-18.
[6] 尤杨,刘弘,吴荣升,等.低温胁迫对香樟幼树抗寒性的影响[J].广东农业科学,2008(11):23-25.
[7] 李志勇,王彦辉,于彭涛,等.重庆酸雨区马尾松香樟混交林的土壤化学性质和林木生长特征[J].植物生态学报,2010,34(4):387-395.
[8] 王江,张崇邦,柯世省,等.添加污泥对尾矿砂理化性质及香樟生理特性的影响[J].生态学报,2010,30(10):2593-2602.
[9] 张宪政.植物叶绿素含量测定[J].辽宁农业科学,1986(3):26-28.
[10] 侯建华,王茅燕,李明哲.玉米萌发期抗旱鉴定的初步研究[J].内蒙古农牧学院学报,1994,15(3):19-22.
[11] 李昌晓,钟章成,刘芸.模拟三峡库区消落带土壤水分变化对落羽杉幼苗光合特征的影响[J].生态学报,2005,25(8):1953-1959.
[12] 张习敏,申刚,陈玲,等.茅台酒厂生态功能区4种优势树种光合特性比较[J].西南林业大学学报,2014,34(5):53-58.
[13] 李昌晓,钟章成.模拟三峡库区消落带土壤水分变化条件下对水松幼苗光合生理响应[J].北京林业大学学报,2007,29(3):23-28.
[14] Kozlowaki T T. Responses of woody plants to flooding and salinity [J]. Tree Physiology Monograph,1997,1:1-29.
[15] Regehr D L, Bazzaz F A, Boggess W R.Photosynthesis, transpiration and leaf conductance inPopulusdeltoidesin relation to flooding and drought[J].Photosynthetica, 1975, 9:52-61.
[16] 陈芳清,郭成园,王传华.水淹对秋华柳幼苗生理生态特征的影响[J]. 应用生态学报,2008,19(6):1229-1233.
[17] Li X L, Li N ,Yang J. Morphological and photosynthetic responses of riparian plantDistyliumchinenseseedlings to simulated Autumn and Winter flooding in Three Gorges Reservoir Region of the Yangtze River, China [J]. Acta Ecologic Sinica,2011,31(1):31-39.
[18] 衣英华,樊大勇,谢宗强,等. 模拟水淹对枫杨和栓皮栎气体交换、叶绿素荧光和水势的影响[J].植物生态学报,2006,30(6):960-968.
[19] 陈静,秦景,贺康宁. 水分胁迫对银水牛果生长及光合气体交换参数的影响[J].西北植物学报,2009,29(8):1649-1655.
[20] 李灵玉,朱帆,王俊刚. 水分胁迫下臭柏光合特性和色素组成的季节变化[J].生态学报,2009,29(8):4346-4351.
[21] 何军,许兴,李树华,等.水分胁迫对牛心朴子叶片光合色素及叶绿素荧光的影响[J].西北植物学报,2004,24(9):1594-1598.
[22] 李玉灵,朱帆,王俊刚,等. 水分胁迫下臭柏(Sabinavulgaris)光合特性和色素组成的季节变化[J].生态学报,2009,9(8):4346-4352.
[23] 张克清. 淹水胁迫对5种豆科植物生长及生理性状的影响[D].南京:南京林业大学,2007:30-31.
(责任编辑 赵粉侠)
Effects of Water Stress on Photosynthesis Characteristics ofCinnamomumcamphoraandLinderamegaphyllaSeedlings
DING Qiong,ZHANG You-lin,LIU Gang,XU Zhen-dong,FEI Yong-jun
(Phoebe Germplasm Resources Evaluation and Innovation Center,Yangtze University,Jingzhou Hubei 434025,China)
Based on 2-year-oldCinnamomumcamphoraandLinderamegaphyllaseedlings,the photosynthetic indices ofCinnamomumcamphoraandLinderamegaphyllaunder different water treatments,were measured and then their photosynthetic physiology response mechanisms were analyzed.The results indicated that chlorophyll content and photosynthesis rate aimed to both ofLinderamegaphyllaandCinnamomumcamphoraincreased at first,and then decreased under water logging and drought stress conditions.With the prolong of water stress′s time,transpiration rate dropped gradually,yet the conductivity was increasing gradually,and obviously increased in the later period of stress.Compared toCinnamomumcamphora,Linderamegaphyllawas more sensitive to water stress,and tolerance was enhanced,showing more flooding and drought tolerance.
water logging stress;drought stress;Cinnamomumcamphora;Linderamegaphylla;photosynthetic characteristics
2014-12-29
湖北省科技支撑计划项目(2013BBB24)资助。
费永俊(1965—),男,博士,教授。研究方向:园林植物种质资源研究与评价。Email:411642145@qq.com。
10.11929/j.issn.2095-1914.2015.04.003
S723.1
A
2095-1914(2015)04-0014-07
第1作者:丁琼(1990—),女,硕士生。研究方向:园林植物种质资源研究与评价。Email:514147032@qq.com。
