棉花种质种仁含油量测定及其遗传多样性分析
2015-03-15刘剑光吴巧娟许剑文肖松华
赵 君, 刘剑光, 吴巧娟, 赵 亮, 许剑文, 肖松华
(江苏省农业科学院经济作物研究所/农业部长江下游棉花油菜重点实验室,江苏 南京 210014)
棉花是中国重要的经济作物,在国民经济发展中具有举足轻重的作用[1-3]。棉花不仅能够提供重要的天然纺织原料,而且棉子中含有丰富的蛋白质、脂肪、棉子糖、维生素等,棉仁中含有30.00%以上的脂肪,其中亚油酸含量高达55.60%,为加工高级保健油的原料,对预防和治疗冠心病、动脉硬化、高血压和高血脂等心脑血管疾病具有显著疗效[4]。然而,长期以来限制棉子高效利用的根本原因在于其中含有1%~2%棉酚及其衍生物。棉酚对人和单胃动物有毒害,食用后胃粘膜组织易被破坏,引起消化功能紊乱,因此棉子这种食物资源未能被充分开发利用[5]。随着中国内地棉花种植面积的不断减少以及植棉经济效益的降低,棉花育种家越来越重视特种棉花品种的选育,其中低酚棉的种仁棉酚含量低于国家食用标准(<0.02%)和国际食用标准(<0.04%),具有重要利用价值,所以选育低酚棉将是提高棉花综合利用价值的一个重要方向。
棉子含油量是受多基因控制的数量性状,而且不同研究者所采用的材料和方法不同,获得的研究结果也存在差别,在不同的研究中主要存在上位性、加性和显性效应以及母体影响[6-8]。另外研究发现,棉子含油量也受环境的影响[9-10]。棉子油的主要成分是脂肪酸,而棉子中脂肪酸的代谢途径受到多个酶的调节,其中柠檬酸合成酶和乙酰CoA羧化酶是2个最关键的限速酶。油酰卵磷脂基因(FAD2-1)是决定脂肪酸不同组分含量的关键酶。磷酸烯醇式丙酮酸羧化酶的活性可以控制棉子蛋白质和油脂的比例,预测通过调节磷酸烯醇式丙酮酸羧化酶基因的表达改变棉子油分与蛋白质比例,可以提高棉子中的脂肪酸含量,创造高含油量的棉花材料[11]。
为了提高棉花的综合利用价值,育种目标既要重视皮棉产量、纤维品质、抗病性和抗虫性的改良,又要兼顾棉子种仁含油量的提高。本研究以高含油量和低酚棉种质系为试验材料,对32份棉花种质系进行遗传多样性分析,期望为高油、低酚棉花新品种选育奠定理论基础。
1 材料与方法
1.1 供试材料
本研究所用材料包含17个低酚棉品种(系),其中9份由中国农业科学院棉花研究所提供,分别是冀棉19、冀棉21、冀棉27、湘棉18号、辽棉11号、中棉所18、中棉所20、汾低99和海1,引入后严格自交繁殖保纯;8份由江苏省农业科学院选育,分别是苏研 602、苏研603、苏研604、苏研605、苏研606、苏研607、苏研608和苏研609,这些材料低酚性状纯合。12个种仁高含油量品种(系),分别为荆55263、绵阳73-39、RNT78、紫色美棉、澳 siv-2、澳 C、澳L23、苏联棉91系、库车T94-4、远93抗354、邯郸长绒和M11。另外3个品种为苏棉8号、陆地棉遗传标准系TM-1和泗抗1号,由江苏省农业科学院经济作物研究所保存。
1.2 SSR 分析
取各品种(系)幼嫩叶片,DNA提取方法参照Paterson等[12]的CTAB法。根据南京农业大学公布的遗传图谱[13]均匀选取分布于棉花26条染色体上的370对SSR引物。所有引物信息可从http://www.genome.clemson.edu/projects/cotton 网站下载。分子标记名称中的大写字母,如 BNL,JESPR等,代表引物的来源。Taq酶、dNTPs和PCR反应的其他试剂均购自北京天为时代公司。PCR扩增与染色参照张军等[14]的方法。
1.3 棉花种仁含油量的测定以及脂肪酸组分分析
棉花种仁含油量测定采用国家标准GB/T 10359-1999测定方法。收取成熟期相近,成熟好的棉桃,经浓硫酸脱绒。精选200粒饱满种子,使用工具钳将种壳夹出裂口,除去种壳后获得种仁,利用研钵分别对每份种仁进行充分研磨变成粉末状。将滤纸放于105℃烘箱内烘干2.5 h,置于干燥器内冷却至室温,称质量(M1)。将1 g左右棉仁粉放入滤纸包内,105℃烘箱内烘干2.5 h,置于干燥器内冷却至室温,称质量 (M2)。将称质量过后的装有棉仁粉的纸包放入抽提管内,使用无水乙醚浸泡16 h,再使用无水乙醚抽提8 h,控制抽提速度,保证1 h内抽提6~8次。抽提后,取出纸包,静止过夜,待残留乙醚完全散发后,在105℃烘箱内烘干2.5 h,称质量(M3)。棉仁含油量计算公式为:M=(M2-M3)/(M2-M1)×100%。每份样品含油量数据均测定3次,取平均值,并保留两位有效小数。
脂肪酸组成分析参考高建芹等[15]方法。
1.4 数据统计
对凝胶上清晰可见且有差异的条带进行记录,按分子量从大到小的顺序计带,有带赋值为“1”,无带赋值为“0”,将电泳图谱数字化,运用统计分析软件NTSYSpc-2.10分析数据。利用非加权类平均法(Un-weighted pair group mathematics average,UPGMA)进行聚类分析,并绘成树状图。计算位点多态信息含量
2 结果与分析
2.1 种仁含油量及不同棉种脂肪酸含量分析
在本研究中,我们选择了其中24个棉花品种(系),测定了其种仁含油量和脂肪酸组分。结果显示,24个品种(系)中包括常规陆地棉栽培品种(系)12个,分别是苏研602、苏研603、苏研604、苏研605、苏研606、苏研607、苏研608、苏研609、冀棉19、冀棉21、湘棉18号和泗抗1号,它们的含油量为28.26% ~32.14%,相互之间不存在显著差异;但与其他12个高油品系相比,存在显著差异。脂肪酸组分方差分析结果显示,不同品种(系)之间脂肪酸组分存在显著差异(表1),但是在高含油量品种和低含油量品种之间没有明显差异,即脂肪酸不同组分的含量与含油量之间没有显著相关性(表2)。
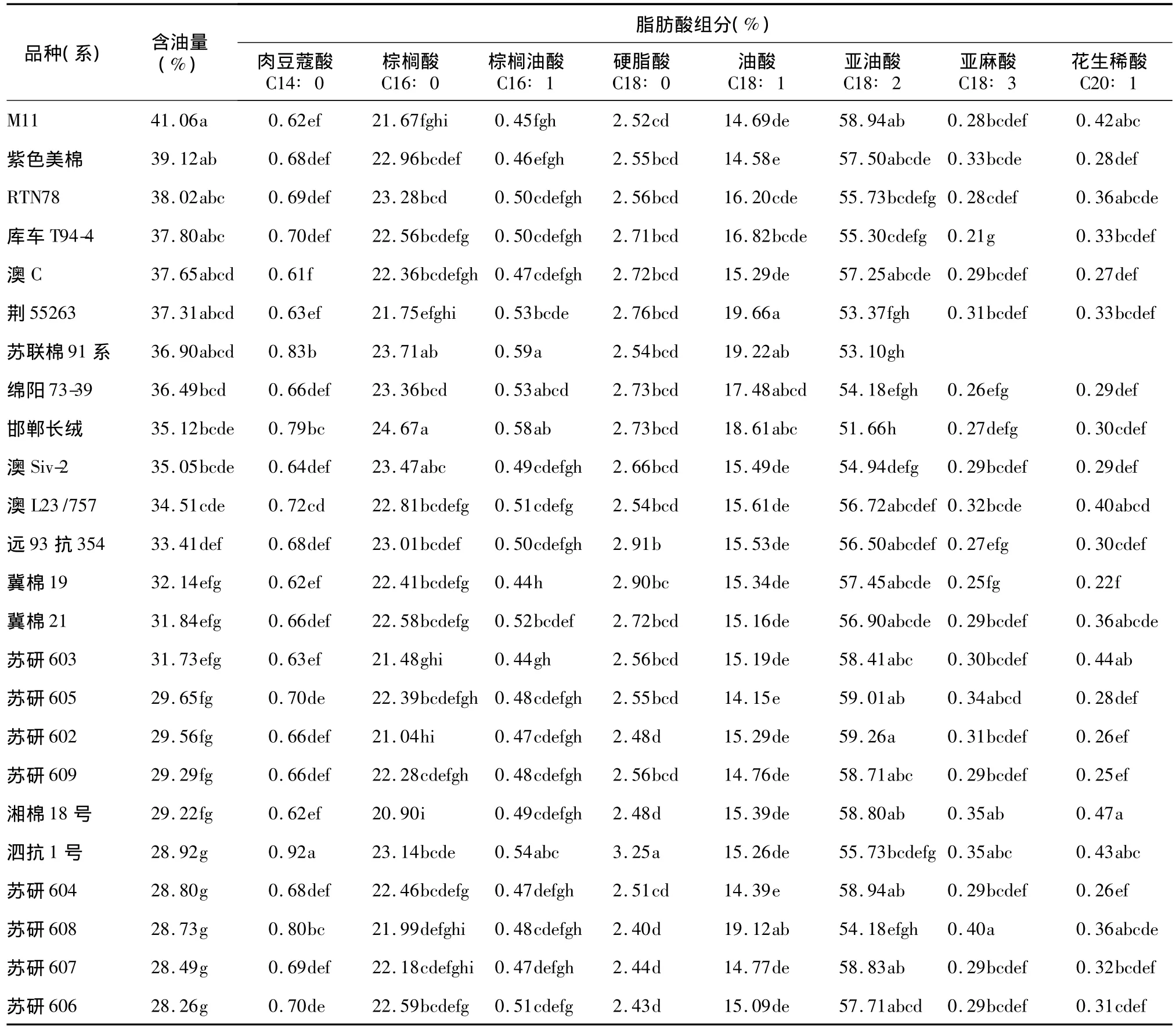
表1 不同棉花品种(系)种仁含油量和脂肪酸组分Table 1 The oil content and percentage of fatty acids in different cotton varieties
根据脂肪酸组分测定结果,我们分析了不同品种间饱和脂肪酸、单不饱和脂肪酸和多不饱和脂肪酸的含量。其中,饱和脂肪酸包括肉豆蔻酸、棕榈酸和硬脂酸;单不饱和脂肪酸包括棕榈油酸、油酸和花生烯酸;多不饱和脂肪酸包括亚油酸和亚麻酸。结果显示,饱和脂肪酸和不饱和脂肪酸的含量与含油量之间不存在显著相关性。饱和脂肪酸含量变幅为24.00% ~28.20%,其中含量最高的是邯郸长绒,最低的是湘棉18号。单不饱和脂肪酸含量变幅为15.12% ~20.52%,其中含量最高的是荆55263,最低的是苏研604。多不饱和脂肪酸含量变幅为51.93% ~59.57%,其中含量最高的是苏研602,最低的是邯郸长绒(表3)。
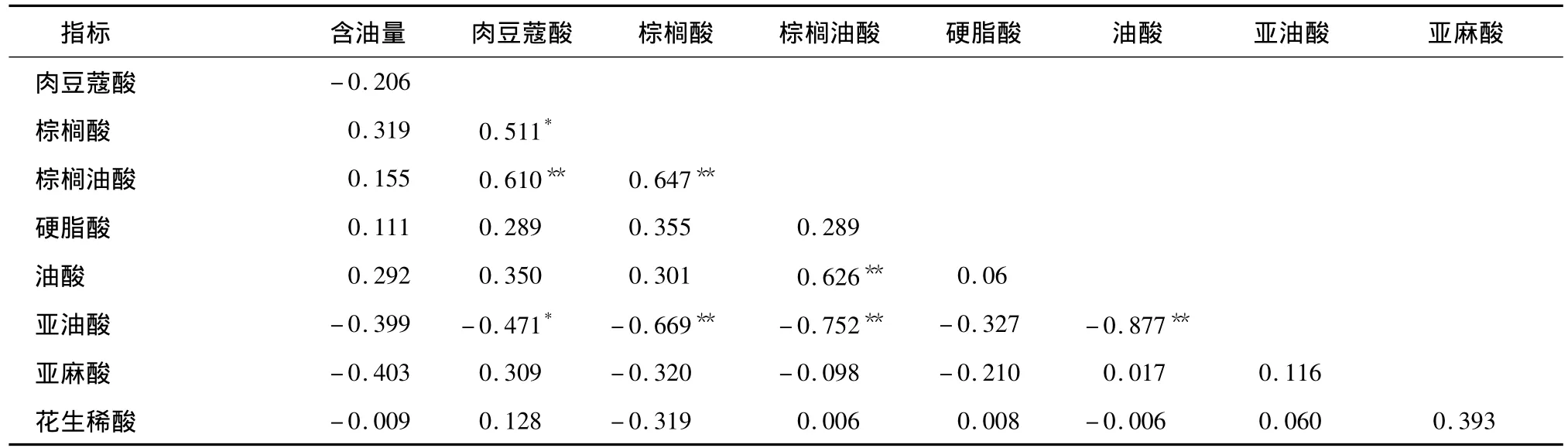
表2 含油量与脂肪酸组分间的相关系数Table 2 Correlation coefficients between oil content and percentage of fatty acid

表3 不同棉花品种(系)间饱和脂肪酸和不饱和脂肪酸含量Table 3 The contents of saturated and unsaturated fatty acids in cotton varieties
对24个品种(系)的含油量和脂肪酸组分的遗传多样性分析结果显示,在供试的24个品种(系)中,不同品种(系)的含油量和脂肪酸组分均呈正态或近似正态分布;脂肪酸组分中,肉豆蔻酸、硬脂酸和亚麻酸在不同品种(系)中分布比较集中,遗传多样性较差。棉花种仁含油量在不同品种之间差异很大,变 幅 为 28.26%~41.06%,变 异 系 数 为12.17%。其中含油量最高的是 M11,达到41.06%;最低的是苏研606,含油量为28.26%。在供试的24个品种(系)中,棉花种仁脂肪酸组分在不同品种(系)之间存在非常大的差异,其中变异最大的是花生烯酸,变异系数达到20.30%;肉豆蔻酸含量变幅为0.61% ~0.92%,其中含量最高的是泗抗1号,最低的是澳 C;棕榈酸含量变幅为20.90% ~24.67%,其中含量最高的是邯郸长绒,最低的是湘棉18号;棕榈油酸含量变幅为0.44%~0.59%,其中含量最高的是苏联棉91系,最低的是冀棉19;硬脂酸含量变幅为2.40% ~3.25%,其中含量最高的是泗抗1号,最低的是苏研608;油酸含量变幅为14.15% ~19.66%,其中含量最高的是荆55263,最低的是苏研 605;亚油酸含量变幅为51.66% ~59.26%,其中含量最高的是苏研602,最低的是邯郸长绒;亚麻酸含量变幅为 0.21%~0.40%,其中含量最高的是苏研608,最低的是库车T94-4;花生稀酸含量变幅为0.22% ~0.47%,其中含量最高的是湘棉18号,最低的是冀棉19(表4)。
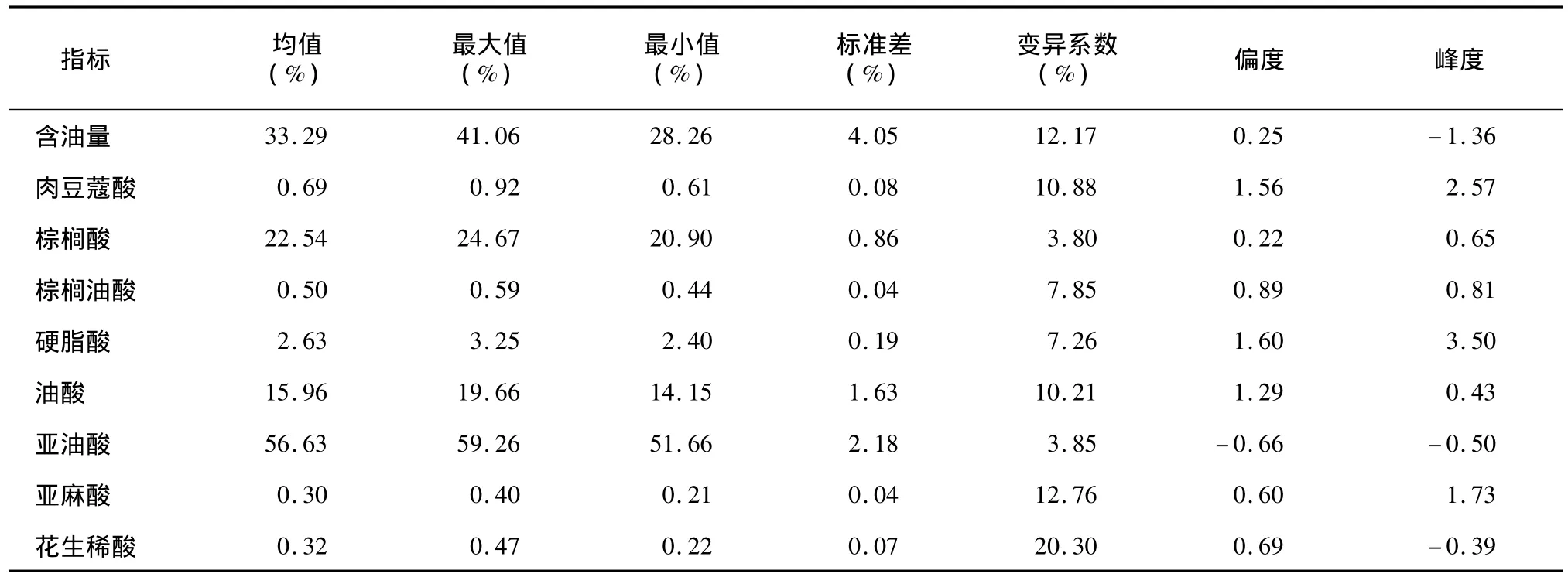
表4 种仁含油量及脂肪酸组分的统计分析Table 4 Statistical analysis of oil content and percentage of fatty acid in 24 cotton varieties
2.2 SSR引物筛选及标记信息分析
依据南京农业大学构建的遗传图谱[13],在各连锁群每隔10 cM选取1个SSR标记位点,共选用SSR标记370对。对供试的32份材料进行多态性初筛,其中190对引物扩增稳定,并表现出多态性,多态性比率为51.35%(表5)。
其中A亚组多态性位点百分率为59.16%,D亚组为43.46%。在190个多态性位点上共检测到609个等位变异,各位点检测到的等位变异数不同,变幅为2~9个,平均为3.09个;只检测到2个等位变异的位点有78个,占总多态位点的41.05%;检测到3个等位变异的位点50个,占26.31%;检测到4个等位变异的位点33个,占17.37%;检测到5个等位变异的位点17个,占8.95%;检测到6个及6个以上等位变异的位点仅12个,占6.32%。检测到2~3个等位变异的位点数占总位点数的67.38%,说明试验选择的资源中等位变异数普遍较少,遗传多样性较低。190个位点多态信息含量(PIC)变幅为0.07~0.81,平均为0.40;多样性指数变幅为0.15~0.83,平均为0.46;其中检测到最多等位变异数的标记是BNL3383,位于D9染色体,检测到的等位变异数是9个,PIC值为0.81。A、D亚组分析结果显示,A亚组平均等位变异数为3.25,多样性指数为0.49;D亚组平均等位变异数为3.06,多样性指数为0.47,说明陆地棉A亚组遗传多样性略高于D亚组。D亚组可能携带更多的控制重要育种目标性状的基因,在长期的品种改良中承受着更高的选择压力,从而造成更多、更强的选择牵连效应(表6)。
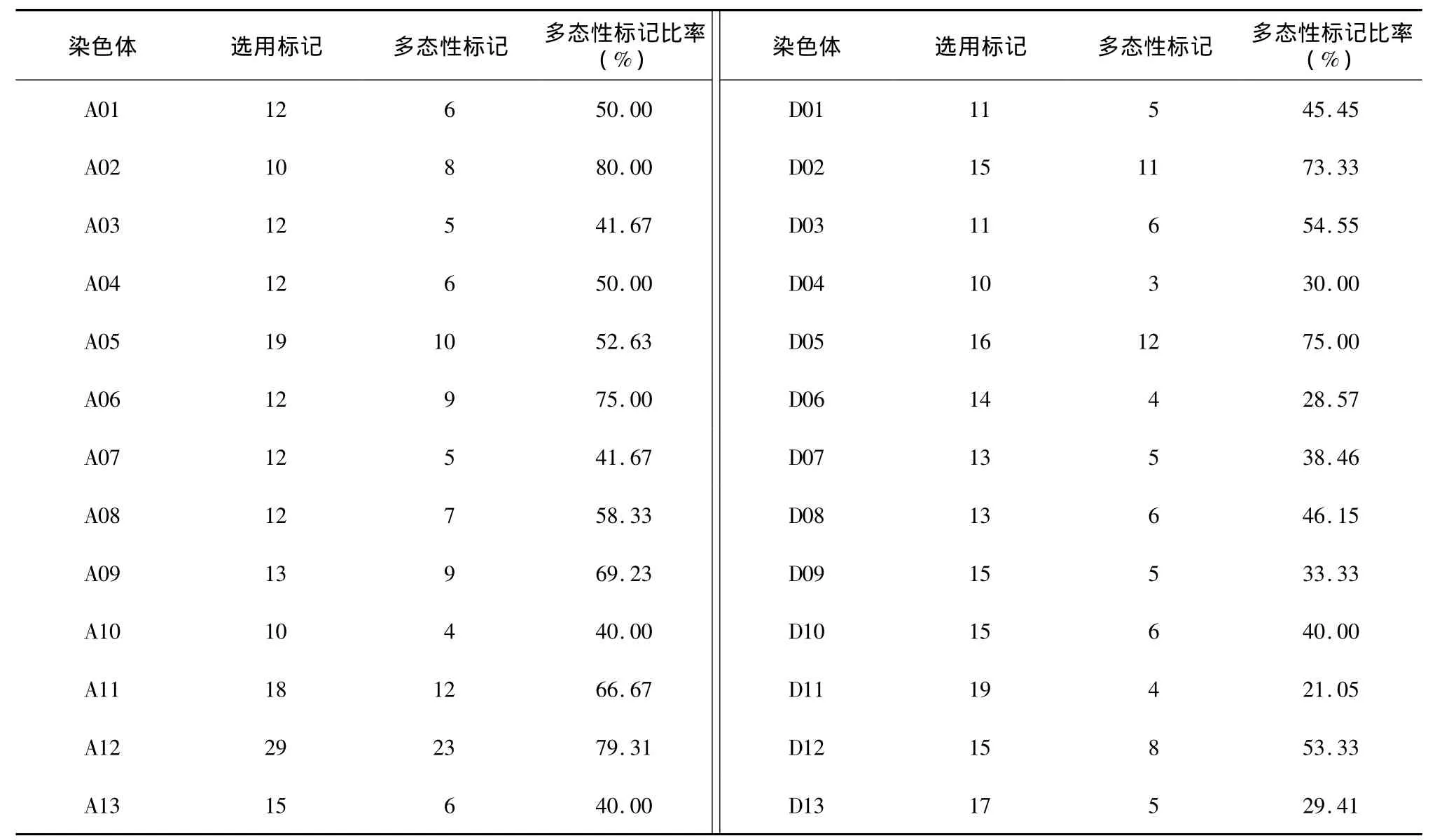
表5 基因组多态性标记分布Table 5 Polymorphic marker distribution on 26 chromosomes of 32 cotton varieties
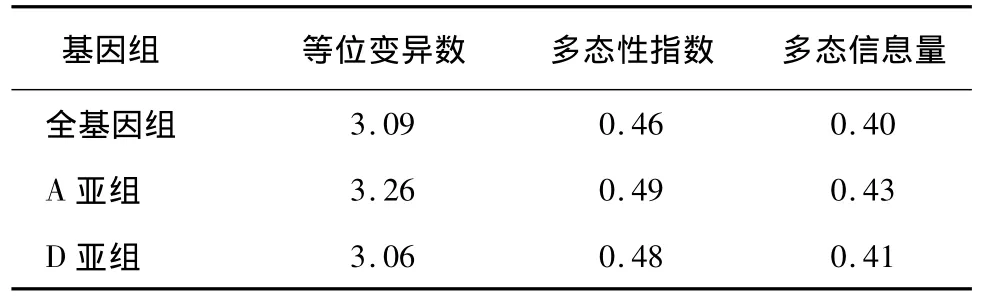
表6 190对SSR位点多态性信息统计Table 6 Information of 190 polymorphic SSR loci
2.3 遗传多样性和系谱分析
明确品种间亲缘关系是育种中亲本组配的前提。运用统计分析软件NTSYSpc-2.10分析32份材料的遗传多样性。利用非加权类平均法(Un-weighted pair group mathematics average,UPGMA)进行聚类分析,并绘成树状图(图1)。32份材料被分成5个类群,在第一类群中包含本研究所有的19个陆地棉低含油量品种(系),分别是苏研系列品种(系)、冀棉19、冀棉21、冀棉27、辽棉11号、汾低99、泗抗1号、中棉所18、中棉所20、TM-1以及苏棉8号。第二类群和第五类群只包含1个品种,分别是海1和紫色美棉。第三类群包含高含油量品种(系)绵阳73-39和荆55263。第四类群包含剩余的高油品种(系),分别是 RNT78、澳 siv-2、澳 C、澳 L23、苏联棉91系、库车 T94-4、远93抗354、邯郸长绒和 M11。由分类结果可知,本研究所涉及的32个品种(系)中的高含油量品种(系)与低含油量品种(系)的遗传差异比较大,遗传距离远。高含油量品种(系)分布于3个类群,表明高含油量种质资源遗传基础比较丰富。
2.4 标记与棉花种仁含油量的关联分析
为减少群体结构和个体间亲缘关系对关联分析结果的影响,本研究分别利用TASSEL 2.1软件[17]的一般线性模型(General linear model,GLM;Q)和混合线性模型(Mixed linear model,MLM;Q+K)程序,将196个位点的等位变异分别与种仁含油量进行关联分析。
与种仁含油量关联的位点见表7。采用一般线性模型检测到与种仁含油量性状显著关联的位点11个,分布在9条染色体上,其中A亚组7个,D亚组4个;混合线性模型检测到与种仁含油量性状显著关联的位点9个,分布在7条染色体上,其中A亚组6个,D亚组3个,与标记NAU2325和NAU5428连锁的位点没有检测到。A12和D3染色体上检测到2个位点,A12染色体上的位点分别是标记BNL598和标记NAU3294,D3染色体上的位点分别是标记NAU2325和NAU4052。一般线性模型检测到的位点解释变异 0.056 0~0.144 9,其中达到极显著水平的3个,分别是A12、D2和A2染色体上的标记BNL598、CIR246和NAU5384;采用混合线性模型检测到的位点解释变异 0.043 6~0.132 3,其中达到极显著水平的位点3个,与一般线性模型检测到的极显著位点相同(表7)。

图1 32份材料的聚类分析图Fig.1 Clustering analysis of 32 cotton germplasm
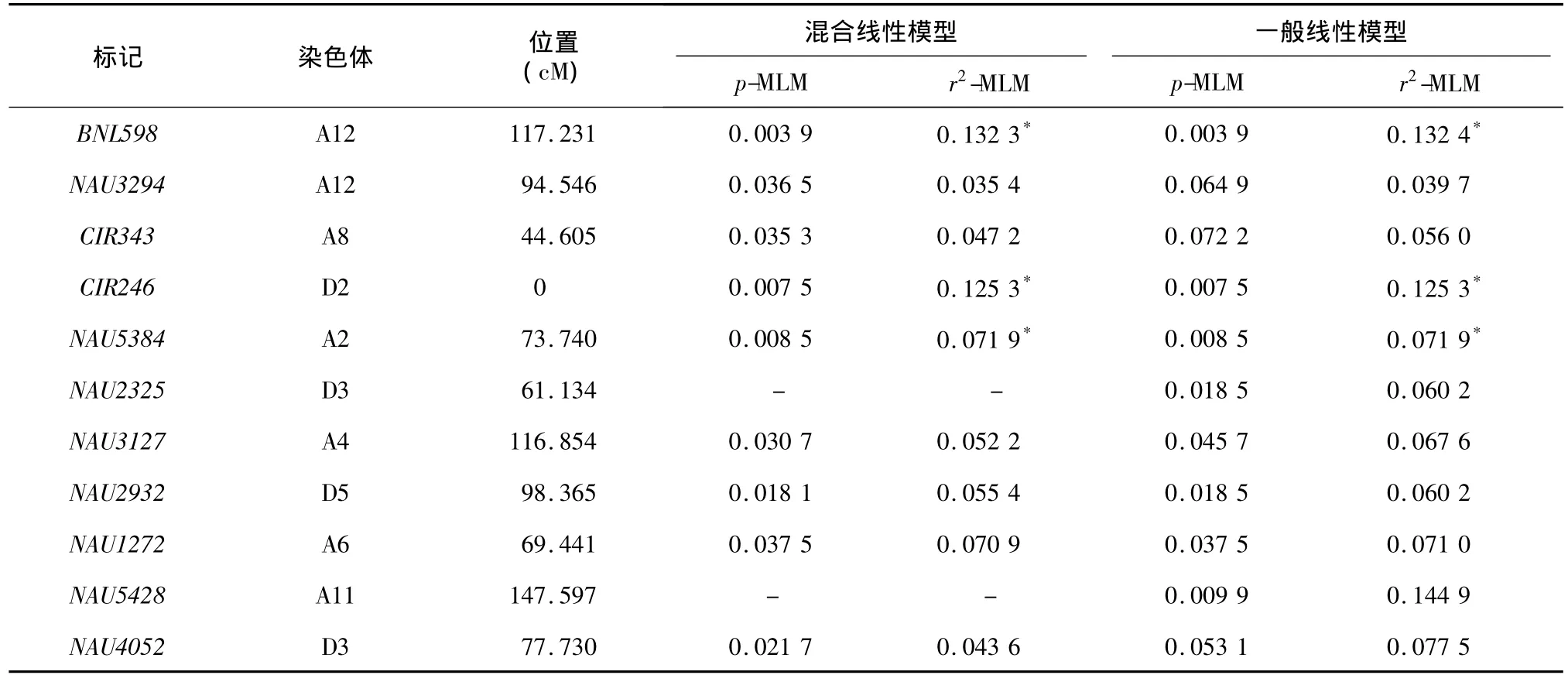
表7 与棉仁含油量性状显著相关的标记位点及其对表型变异的解释率Table 7 Marker loci associated with oil content trait and their explained phenotypic variations
3 讨论
棉花是一种重要的经济作物,它不仅给人类提供了纤维原料,而且种子含有18% ~20%的油脂,棉花种仁直接榨油可生产高质量的食用油,由于棉酚存在,极大地限制棉花种仁的利用。低酚棉种仁榨油无需精炼脱毒,可直接食用,所以选育显性低酚高含油量棉花种质对今后提高棉花整体利用价值具有重要意义[4]。本研究利用32份棉子材料其中包含17个低酚棉品种(系)和12份高油材料,测定其种仁含油量及脂肪酸组分,结果显示,通过杂交选育的低酚棉品种(系)种仁含油量显著低于高油材料,这与前人报道的含油量的遗传偏向低油亲本一致,即通过多代杂交选育,棉花种仁含油量将会不断降低[18]。同时也有研究结果表明,棉花的母体效应对子代含油量存在影响,其效应要高于遗传影响,这意味着在品种选育过程中选择高含油量材料或低含油量材料做母本,对杂交后代种仁含油量将会有显著影响[6,19]。
利用分子标记研究不同棉花品种(系)的遗传多样性,分析其遗传背景的差异,可以为今后通过配制棉花杂交组合选育高油低酚棉花种质提供参考。本研究将32份材料通过聚类发现,低含油量品种(系)和高含油量品种(系)分为2大类,而且这2类群之间亲缘关系比较远。前人的研究结果表明,种仁含油量与棉花产量之间没有相关性,与棉酚含量呈正相关[20-21]。说明在棉花产量及相关性状的改善和提高时不会降低棉花种仁含油量。出现现有品种(系)的含油量显著低于高油种质的含油量的现象,主要有两方面的原因:(一)棉花种仁含油量的遗传模式可能会使杂交后代含油量降低;(二)棉花种仁含油量不是育种家在棉花品种选育过程中关注的主要指标,导致现有品种(系)含油量的降低。
棉花种仁含油量是数量性状,受多基因控制。Yu等[22]利用陆地棉与海岛棉回交群体共检测到17个与油份含量相关的QTLs,分布在12条染色体上,其中位于A12染色体上检测到2个QTLs,其中1个解释表型变异23.64%。刘小芳等[23]以陆地棉重组自交系群体为材料共检测到8个与含油量相关的QTLs,其中位于D7染色体上的1个QTL在2个环境中能同时检测到。李晓娜等以89份半野生棉为材料,应用关联分析,在3个年份环境下重复检测出12个与棉仁含油量显著相关的位点[24]。本研究检测到的与含油量相关的位点与前人研究部分相同,这与研究材料不同有关系。另外本研究获得的与已经报道的不同的与棉仁含油量性状显著相关的标记位点,可能为棉仁含油量性状的分子标记辅助选择提供依据。
[1] 吴巧娟,刘剑光,赵摇君,等.棉花耐盐碱性状的QTL定位[J].江苏农业学报,2014,30(5):966-971.
[2] 蔡立旺,陈摇源,王永慧,等.棉花钾素吸收利用效率与产量的关系[J].江苏农业学报,2014,30(5):972-979.
[3] 李建刚,张卫国,李东方,等.不同施肥模式对新疆棉花产量和品质的影响[J].江苏农业科学,2014,42(2):55-58.
[4] 肖松华,吴巧娟,刘剑光,等.显性低酚棉新品系种仁营养品质与利用评价[J].棉花学报,2012,24(2):127-132.
[5] 中国农业科学院棉花研究所.中国棉花遗传育种学[M].济南:山东科学技术出版社,2009.
[6] YUANY L,ZHANGTZ,JINGSR.Studies of the inheritance of seed qualities and the exploitation of F2heterosis in low gossypol strains in upland cotton[J].Acta Genetica Sinica,2001,28(5):471-481.
[7] 王国印,李蒙恩.棉子品质性状的遗传研究[J].华北农学报,1991,6(2):20-25.
[8] 朱乾浩.低酚棉种仁含油量的母体效应和杂种优势分析[J].种子,1994(5):5-8.
[9] 韩智彪.棉籽油份含量近红外测定技术研究[D].武汉:华中农业大学,2012.
[10]李文峰,孟亚利,王友华,等.棉籽蛋白质和油分含量预测的生态模型[J].生态学杂志,2011,30(11):2653-2658.
[11]王彦霞,刘正杰,马峙英,等.RNA干涉技术与棉花高油育种[J].棉花学报,2011,23(2):178-183.
[12] PATERSON A H,BRUBAKER CL,WENDEL JF.A rapid method for extraction of cotton(Gossypium spp.)genomic DNA suitable for RFLPand PCRanalysis[J].Plant MolBiol Rep,1993,11:122-127.
[13] ZHAO L,LV Y D,CAI C P,et al.Toward allotetraploid cotton genome assembly:integration of a high-density molecular genetic linkage map with DNA sequence information[J].BMCGenomics,2012,13:539.
[14]张 军,武耀廷,郭旺珍,等.棉花微卫星标记的PAGE/银染快速检测[J].棉花学报,2000,12(5):267-269.
[15]高建芹,浦惠明,戚存扣,等.应用气相色谱仪分析油菜脂肪酸含量[J].江苏农业学报,2008,24(5):581-585.
[16]张小娟,何团结,陆徐忠,等.陆地棉SSR核心引物筛选及95份骨干种质的遗传多样性分析[J].棉花学报,2011,23(6):529-536.
[17] BRADBURY P,ZHANGZ,KROON D,et al.TASSEL:software for association mapping of complex traits in diverse samples[J].Bioinformatics,2007,23:2633.
[18]廖伯寿,雷 永,王圣玉,等.花生重组近交系群体的遗传变异与高油种质的创新[J].作物学报,2008,34(6):999-1004.
[19] YE Z H,LU Z Z,ZHU J.Genetic analysisfor developmental behavior of some seedquality traits in Upland cotton(Gossypum hirsutum L.)[J].Euphytica,2003,129:183-191.
[20]王国建,朱 军,臧荣春,等.陆地棉种子品质性状与棉花产量性状的遗传相关性分析[J].棉花学报,1996,8(6):295-300.
[21]赵永国,郭瑞星.棉子含油量研究进展与高油棉花育种可行性分析[J].棉花学报,2011,23(2):184-188.
[22] YU JW,YU SX,FAN SL,et al.Mapping quantitativetrait loci for cottonseed oil,protein and gossypol content in a backcross inbred line population[J].Euphytica,2012,187:191-201.
[23]刘小芳,李俊文,余学科,等.利用重组自交系进行陆地棉(Gossypum hirsutum L.)棉籽油分含量和蛋白质含量的QTL定位[J].分子植物育种,2013,11(5):520-528.
[24]李晓娜,蔡小彦,KIFLOM W O,等.半野生棉棉仁含油量与SSR 标记的关联分析[J].棉花学报,2015,27(2):104-110.
