紫背天葵(Begonia fimbristipula Hance)叶片形态和生理生态特征对不同光强的响应
2015-03-12汪越刘楠任海邵玲陈雄伟陈刚王俊
汪越,刘楠,任海,邵玲,陈雄伟,陈刚,王俊*
1. 中国科学院华南植物园,广东 广州 510650;2. 中国科学院大学,北京 100049;3. 肇庆学院生命科学学院,广东 肇庆 526061
紫背天葵(Begonia fimbristipula Hance)叶片形态和生理生态特征对不同光强的响应
汪越1, 2,刘楠1,任海1,邵玲3,陈雄伟3,陈刚3,王俊1*
1. 中国科学院华南植物园,广东 广州 510650;2. 中国科学院大学,北京 100049;3. 肇庆学院生命科学学院,广东 肇庆 526061
摘要:紫背天葵(Begonia fimbristipula Hance)是我国特有的珍稀濒危药用植物。以紫背天葵组培球茎为材料,通过遮阴网设置3种光照强度,分析紫背天葵叶片形态、生理、营养元素及次生代谢产物含量对光照的响应特征,以期为其人工栽培提供理论指导。结果表明,紫背天葵的比叶面积和表观量子效率随光照强度的增大而降低,而上表皮厚度随光照强度的增大而增加。尽管高光照(80%自然光照)处理下的紫背天葵叶片具有更高的最大光合速率、光补偿点、光饱和点、瞬时水分及光能利用效率,但对照(50%自然光照)下的紫背天葵叶片具有更丰富的总花青素含量(0.4 μmol·g-1)。与对照相比,低光照下(20%自然光照)的紫背天葵叶片最大光和速率、暗呼吸速率、光补偿点、光饱和点及瞬时水分利用效率显著降低,且总花青素含量无显著差异。上述结果表明,紫背天葵可通过调节自身形态生理特征来适应较高或较低的光照条件,对不同的光环境有较好的适应能力。但从叶片药用成分保持的角度来看,紫背天葵在接近原生境光照条件的50%自然光照下具有更佳的光合作用和花青素合成能力,可作为其人工栽培的参考光照条件。
关键词:紫背天葵;光照;形态解剖;光合生理;次生代谢产物
引用格式:汪越,刘楠,任海,邵玲,陈雄伟,陈刚,王俊. 紫背天葵(Begonia fimbristipula Hance)叶片形态和生理生态特征对不同光强的响应[J]. 生态环境学报, 2015, 24(6): 957-964.
WANG Yue, LIU Nan, REN Hai, SHAO Ling, CHEN Xiongwei, CHEN Gang, WANG Jun. Responses of Leaf Morphological and Physiological Characteristics of Begonia fimbristipula Hance to Light Intensity [J]. Ecology and Environmental Sciences, 2015, 24(6): 957-964.
紫背天葵(Begonia fimbristipula Hance),属秋海棠科秋海棠属多年生无茎草本植物,为我国特有种,性喜温暖湿润的环境,一般生长在悬崖峭壁的荫蔽石壁上(古粹芝,1999)。自然条件下主要有红葵和绿葵(也称青葵),其中红葵叶片内具有丰富的红色素,已有的研究表明红色素主要成分为花青素,且为其主要药用成分(段志芳等,2006;谭雄斯等,2012;邵玲等,2014)。近十余年,由于过度的人为采摘及自身生长环境脆弱,其自然种群急剧减少,2005年紫背天葵已被《中国珍稀植物》评估为濒危物种的低危等级(邢福武,2005)。目前对紫背天葵的研究主要集中在组培、叶片内化学成分、种群特性调查及小规模的人工栽培试验(张兰英和李耿光,1986;许旋等,2000;邵玲和梁霞,2012)。关于环境条件对紫背天葵形态、生理及次生代谢产物的影响尚未见报道,未能确定其高产优质栽培的光照条件。
光是影响植物生长发育的重要环境因子,光照过量和不足对植物都有一定的影响(Dai等,2009;Deng等,2012)。研究表明高光照条件下植物叶面积减小、叶厚度增大、栅栏组织发达、光饱和点和补偿点较高(覃凤飞等,2012)。低光条件下植物叶片通常较大且薄,光补偿点和饱和点较低,栅栏组织退化而海绵组织发达,在适应较低光照条件的同时最大限度的提高自身的光能利用率(Gratani等,2006)。此外,有关植物体内次生代谢产物与光照环境关系研究表明,植物在适应光照环境时光合能力与体内次生代谢产物积累有一定关系。Croteau等(1972)认为,植物光合作用的增加能提供较多的次生代谢前体,同时抑制了次生代谢产物的分解;光强对阴生植物人参体内次生代谢产物的影响研究表明光强过大时人参皂苷的含量下降(张治安等,1994);王洋等(2004)对喜树次生代谢产物喜树碱对光强的响应研究表明,在严重遮阴的情况下喜树碱含量降低。因此,光强对次生代谢产物的影响与植物本身生长的特性及环境适应能力有很大关系。次生代谢产物多作为药用植物的药用成分,研究光照对药用植物形态、生理及次生代谢产物含量的影响对阐明栽培过程中道地性的维持机理具有重要意义。
本文通过观测人工设置的3种光照梯度下紫背天葵叶片形态、生理、营养元素及次生代谢产物含量特征,主要探讨以下科学问题:(1)紫背天葵在不同光照强度下,其形态、生理和次生代谢产物含量会发生哪些变化?(2)紫背天葵叶片对光的适应性如何?研究旨在为紫背天葵规模化生产过程中适宜光照条件的选择提供科学依据。
1 材料与方法
1.1自然概况及实验处理
实验样地位于广东省广州市中国科学院华南植物园(23°35' N,112°57'E),园内以低丘陵台地为主。属于南亚热带季风气候,夏季炎热潮湿,秋冬温暖干旱,年平均温度21.8 ℃,最热月(7─8月)平均气温28.3 ℃,最冷月(1月)平均温度13.3 ℃。年降水量为1623.6~1899.8 mm,4─9月为湿季,10月─翌年3月为干季(杨勇等,2010)。
本实验紫背天葵球茎由肇庆学院生命科学学院邵玲教授提供。野外自然条件下紫背天葵生于疏林下石上或悬崖石缝中(古粹芝,1999),不同生境光照条件差异较大,通过实地踏查,野外分布的紫背天葵的生境光照约为50%自然光照(50% NL),因此通过不同透光率的遮荫网设定3个光照梯度分别为80%(80% NL)、50%(50% NL)和20%自然光照(20% NL),其中以最接近野外生长条件的50% NL为对照,80% NL为高光照处理,20% NL为低光照处理。将紫背天葵球茎种于花盆,栽培土壤为泥炭土,再将花盆分别置于3种光照梯度下,每个光照处理下3个区组,每个区组10个球茎,栽培期间每天浇两次水。试验于2013年12月开始持续到2014年4月,待其营养生长达到最大时期(4月),进行相关指标的测定,各项指标分别从每个重复中选取1~3片叶子。
1.2研究方法
1.2.1叶面积和比叶面积
将叶片收获洗净后平铺于标准坐标纸上,用数码相机对叶片进行拍照后用Image-J计算叶片面积(Liu和Guan,2012)。之后将叶片置于65 ℃烘箱内烘干至恒重,称量叶片干重后计算比叶面积(SLA=总叶面积/总叶重),每个处理9个重复。
1.2.2叶片横切面指标测量
采用半薄切片法观察叶片横切面结构,每个处理9个重复。从叶片的中部避开叶脉切取0.5 cm×1 cm小叶片后迅速投入4%戊二醛溶液固定,于4 ℃中存放12 h。后再用2%的锇酸固定4 h,梯度乙醇脱水后丙酮过夜,再用EP812环丙树脂包埋,半薄切片机(LKB118000,LKB,Japan)切片(0.5~2 μm),甲苯胺蓝染色后置于光学显微镜(SZX12,Olympus,Japan)下观察并拍照。测量叶片厚度、栅栏组织厚度和海绵组织厚度等指标。
1.2.3叶片表皮结构
利用扫描电镜观察叶片上、下表皮结构,每个处理9个重复。在叶片中部避开叶脉切去5 mm×5 mm的叶片后立即投入4%的戊二醛溶液中,于4 ℃下保存12 h。用0.2 mol·L-1的磷酸缓冲液漂洗数次后梯度乙醇脱水,之后分别置于3∶1、1∶1、1∶3的乙醇:叔丁醇和纯叔丁醇10 min,取出置于真空喷涂仪内进行真空干燥。粘台后于IB-5型离子镀膜仪上喷涂后扫描电镜(日本电子T-300)下观察拍照。统计气孔密度和上、下表皮毛密度等指标。
1.2.4叶绿体超微结构
前处理与半薄切片相同,ULPRACUT EXINXIN型超薄切片机切片(60 nm)后,经醋酸双氧铀柠檬-柠檬酸铅双染色后置于日立H-800透射电子显微镜下观察并拍照。叶绿体大小、基粒厚度等指标为27个叶绿体测定平均值。
1.2.5光响应曲线测定
于晴朗的天气,用便携式光合作用仪(LI-6400,USA)测定其光响应曲线,设定光合有效辐射梯度为:1400、1200、1000、800、500、300、150、100、50、30、15、0 μmol·m-2·s-1。测定过程中保持环境稳定,CO2为400 μmol·μmol-1,叶温25 ℃,每次测定3个重复。利用非直角双曲模型对数据进行拟合分析,并计算最大光合速率(Amax)、暗呼吸速率(Rd)、表观量子效率(AQY)、光补偿点(LCP)、光饱和点(LSP)、瞬时水分利用效率(IWUE=Pn/Tr)、瞬时光能利用效率(ILUE=Pn/PAR)等指标(Nijs等,1997;Liu和Guan,2012)。
非直角双曲模型公式为:

式中:Pn为净光合速率,I为光照强度,ϕ为表观量子效率,Rd为暗呼吸速率,Amax为最大净光合作用速率,θ为曲线性状参数(Larcher,1980)。
1.2.6叶绿素荧光参数测定
利用便携式荧光仪(PAM-2100,Germany)测定叶绿素荧光参数,测定前将叶片置于叶夹中暗适应30 min,每个测量选取9片叶,主要参数如下:光系统II最大光化学效率(Fv/Fm=(Fm-Fo)/Fm)、光化学荧光淬灭(qP=(Fm′-Fs)/(Fm′-Fo′)、非光化学荧光淬灭(NPQ=(Fm-Fm′)/Fm′)。其中,Fm和Fo是暗适应后的最大和初始荧光,Fm′和Fo′是为光适应后的最大荧光和初始荧光(Souza等,2004)。
1.2.7叶绿素含量测定
将叶圆片(直径0.6 cm)置于80%丙酮溶液中,存放于黑暗处,每个处理5个重复。待叶片完全褪色后用紫外分光光度计(UV-3802,Unico)测定浸出液在663、645、440 nm处的吸光值,后计算叶绿素a(Chl a)、叶绿素b(Chl b)和类胡萝卜素含量(Car)(Lin等,1984)。
1.2.8总花青素含量测定
将叶片材料剪碎后置于甲醇和盐酸混合溶液中(V甲醇∶V盐酸=99∶1),存放在4 ℃黑暗处5 d,每个处理5个重复。用紫外分光光度计(UV-3802,Unico)测定浸出液在530和650 nm处的吸光值,后计算总花青素含量(Murray和Hackett,1991;Reddy等,1995)。
1.2.9原花青素含量测定
将叶片材料剪碎后置于甲醇溶液中,用超声波水浴(40 ℃)提取2 h,待提取液冷却后过滤(0.45 μm)。将过滤液置于真空旋转式浓缩仪内,取50 mg浓缩后得到的固体溶解在25 mL蒸馏水中。取2 mL溶解液,加入20 mL硫酸亚铁溶液(盐酸∶正丁醇=2∶3),后将混合液置于95 ℃水浴下加热1 h。用紫外分光光度计测定液体在540 nm处的吸光值,后计算原花青素含量(Skerget等,2005),每个处理5个重复。
1.2.10植物营养元素测定
将叶片材料置于65 ℃下烘干至恒重,用重铬酸钾法测定全碳含量,凯氏定氮法测定全氮含量,钼锑比色法测定全磷含量(刘光崧,1996),每个处理5个重复。
1.3数据分析方法
采用单因素方差分析法(One-way ANOVA)分析光照条件对紫背天葵叶片形态、生理特征的影响,不同光照条件下各生长指标的差异采用LSD进行多重比较分析,数据差异显著临界值P=0.05。并用SPSS统计分析软件包(SPSS 12.0 for Windows,Chicago,USA)对光响应曲线进行拟合,用SigmaPlot及Excel绘制图形。
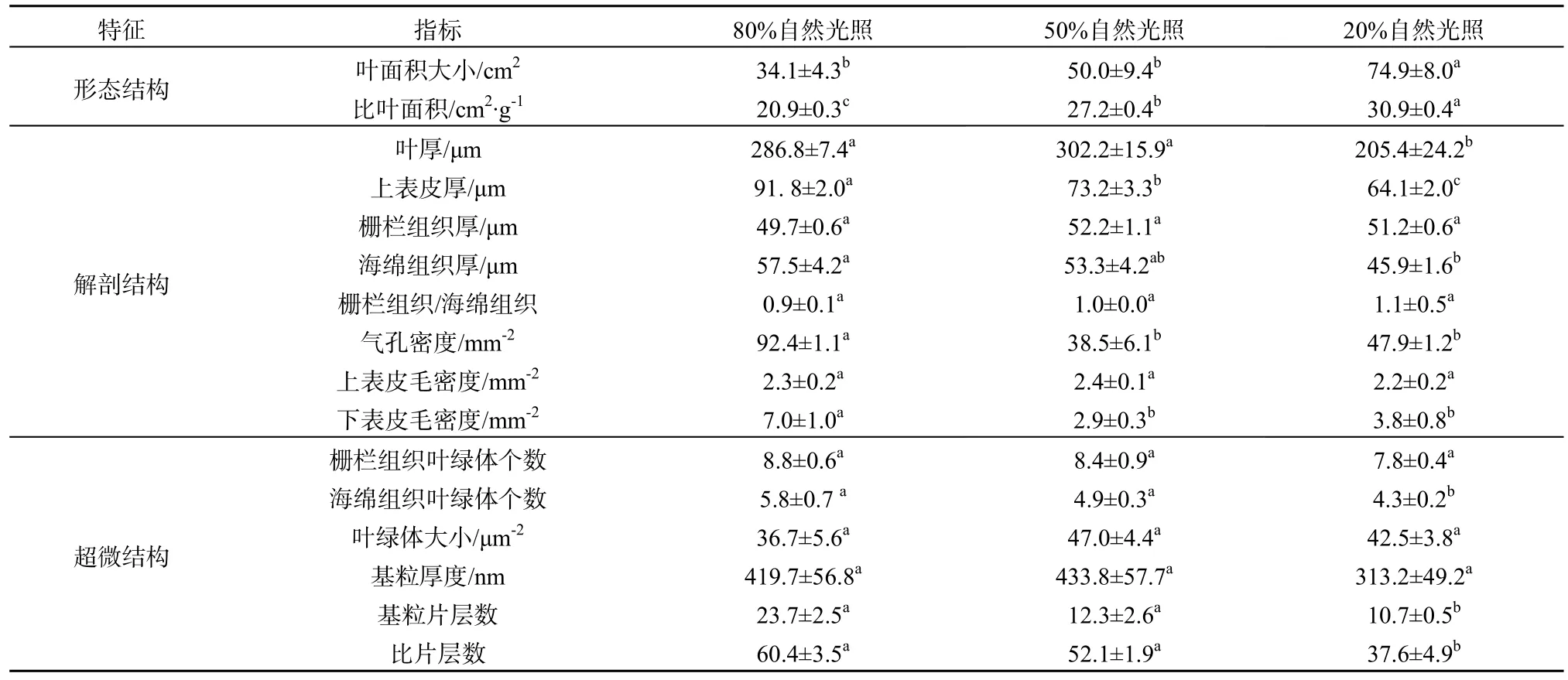
表1 3种光强下紫背天葵叶片的形态和解剖结构特征Table 1 Morphological and anatomical characteristics of B. fimbristipula in three different light conditions
2 结果
2.13种光照处理下叶片的形态和解剖结构特征
方差分析结果表明,光照条件显著影响紫背天葵叶片的形态特性。与接近紫背天葵自然生境的50% NL处理下叶片的形态特征相比,高光照(80% NL)处理后叶片的大小没有显著差异性变化(P=0.187),比叶面积(SLA)显著的减小(P<0.001,表1)。在解剖结构特征上二者差异较小,只有上表皮厚度、下表皮毛密度、气孔密度3个指标在80% NL下要显著大于50% NL,叶厚、栅栏组织厚、海绵组织厚、栅栏组织/海绵组织厚、上表皮毛密度等指标在80% NL和50% NL处理下无显著差异性(P>0.05,表1,图1),且80% NL下叶片上表皮覆盖的蜡质要多于50% NL下(图2G、H)。叶绿体个数和大小、基粒厚度和片层数及比片层数等指标在两个处理间也没有显著性差异(P>0.05,表1,图2)。
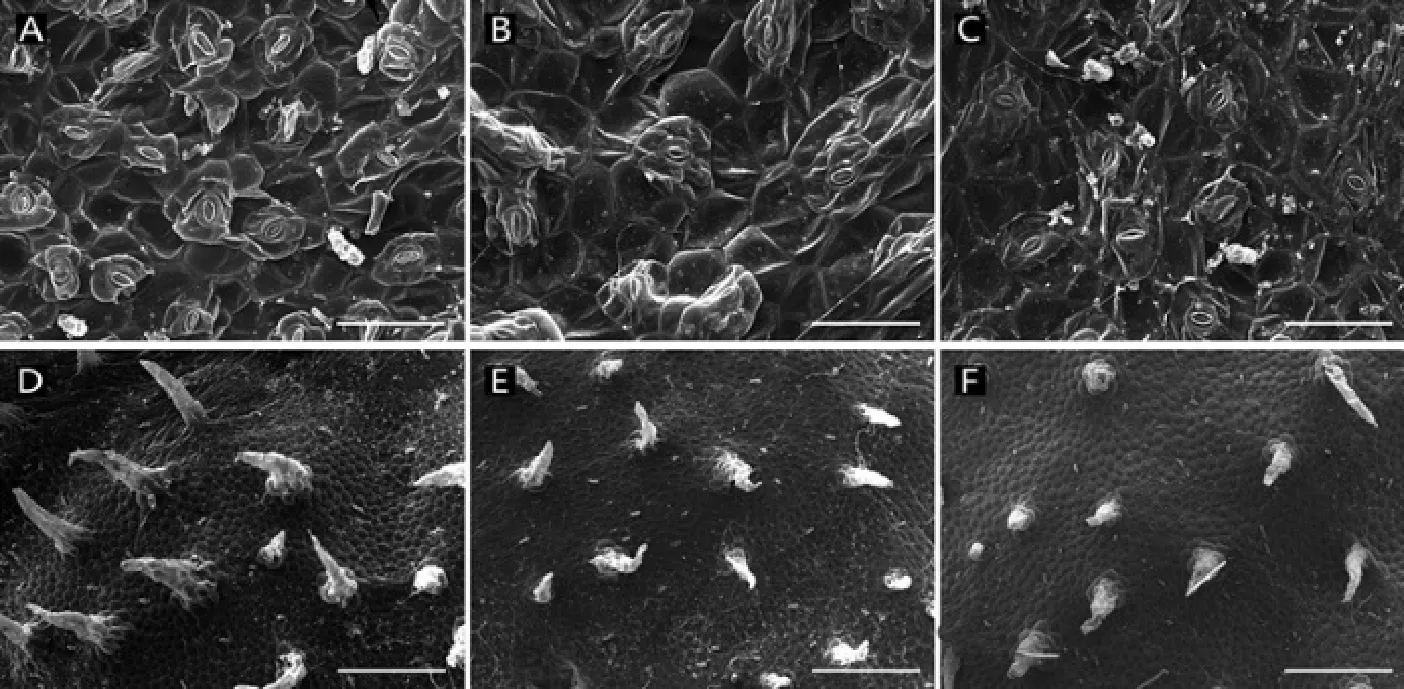
图1 3种光强下紫背天葵表皮扫描图;80%(A)、50%(B)和20%(C)NL下叶片气孔,Bar=100 μm;80%(D)、50%(E)和20%(F)NL下上表皮毛,Bar=500 μmFig. 1 SEM photomicrographs of leaf surface. Stomata of B. fimbristipula in 80% (A), 50% (B), 20% NL(C), Bar=100 μm; epidermic trichome of B. fimbristipula in 80% (D), 50% (E), 20% NL(F), Bar=500 μm
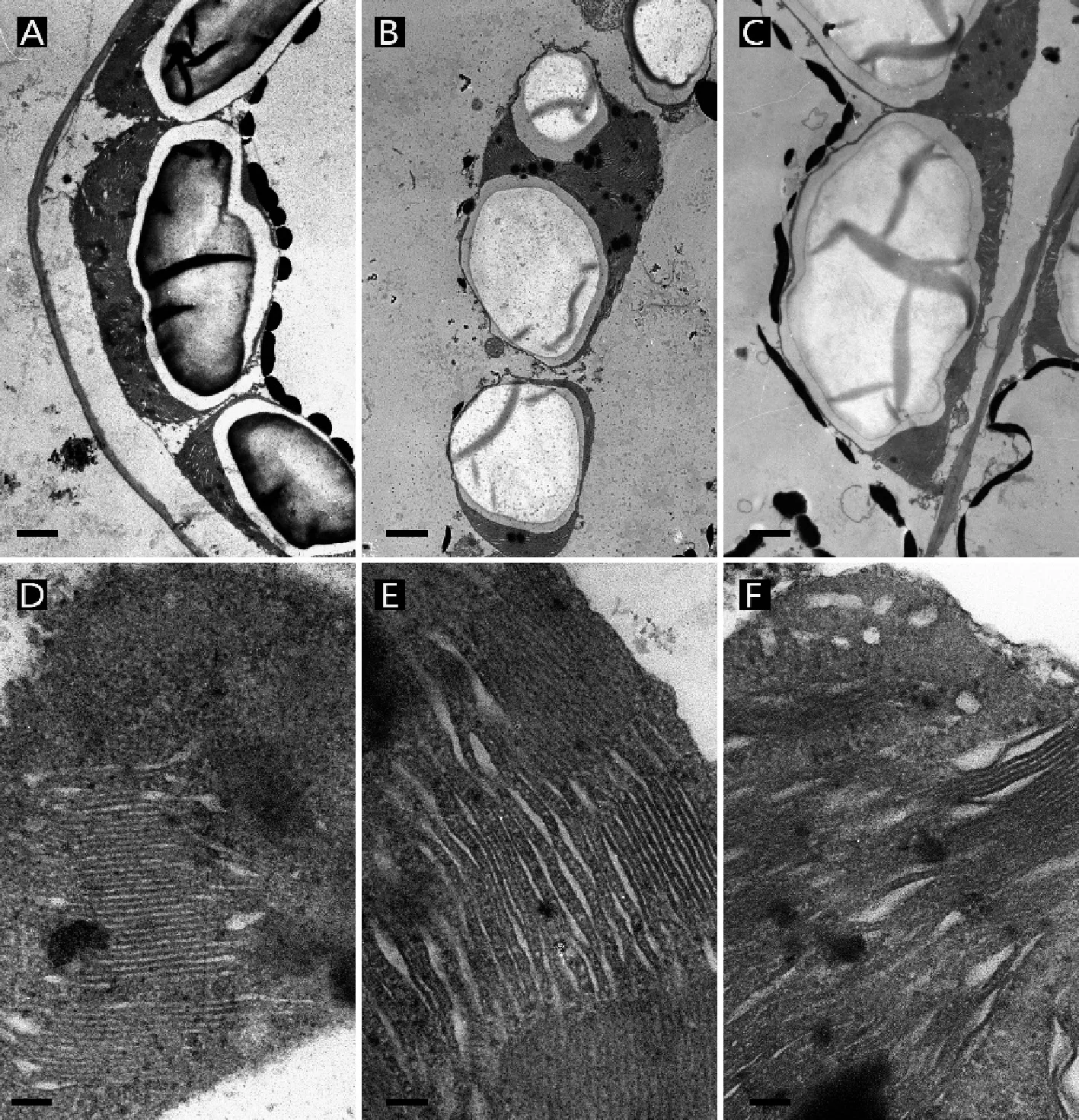
图2 3种光强下紫背天葵叶绿体超微结构,80%(A)、50%(B)和20%(C)NL下叶绿体,Bar=1 μm;80%(A)、50%(B)和20%(C)NL下叶绿体基粒,Bar=100 nmFig. 2 Ultrastructure traits of chloroplasts in leaves of B. fimbristipula in three different light. Chloroplasts of B. fimbristipula in 80% (A), 50% (B), 20% NL(C). Bar=1 μm; Grana lamella of B. fimbristipula in 80% (D), 50% (E), 20% NL(F), Bar=100 nm
与50% NL处理下紫背天葵叶片特征相比,低光照(20% NL)处理后,叶片大小和SLA都显著增大(P<0.05,表1)。叶厚、上表皮厚显著减小(P<0.05),而栅栏组织、海绵组织、栅栏组织/海绵组织厚、气孔密度及上、下表皮毛密度上二者没有显著性差异(P>0.05,表1,图1)。光照处理对叶片超微结构也有一定影响,低光照显著降低了海绵组织叶绿体个数、基粒片层数和比片层数(P<0.05),对栅栏组织叶绿体个数、叶绿体大小、基粒厚度等指标无显著性影响(P>0.05,表1,图2)。低光照处理对叶绿体基粒片层形态发展有一定的影响,与50% NL处理下相比,低光照处理后基粒片层排列更加松散,在电镜下难以被辨别(图2F)。
2.23种光照处理下叶片光合生理特性
光响应曲线参数表明(表2),紫背天葵叶片表观量子效率随着光照强度的降低而显著上升(P<0.001),最大光合速率、光饱和点、光补偿点、瞬时水分利用率都随着光照强度的降低而显著下降(P<0.05)。80% NL下叶片暗呼吸速率与50% NL处理下无显著差异(P=0.843),且二者都显著的大于20% NL处理下叶片的暗呼吸速率(P<0.05)。在瞬时光能利用效率(ILUE)方面,80% NL下叶片有最大的ILUE,显著的大于50%和20% NL处理(P<0.05),后二者在ILUE上无显著差异(P=0.511)。
叶绿素荧光参数特征表明(表2),光照处理对光化学淬灭和非光化学淬灭没有显著影响(P>0.05),20% NL的紫背天葵光系统Ⅱ最大光化学效率显著低于80% NL(P<0.001)。
2.33种光照处理后叶片色素及营养元素含量
与50% NL相比,80% NL处理下叶片的总叶绿素含量、叶绿素a/b及原花青素含量无显著性变化(P>0.05),类胡萝卜素含量显著增加(P=0.028),总花青素的含量显著降低(P=0.021,表3)。20% NL处理下,叶片叶绿素a/b显著减小(P<0.05),而类胡萝卜素及原花青素含量显著增大(P<0.05,表3)。
光照条件对紫背天葵叶片的全氮、全磷含量及氮、磷比无显著影响(P>0.05,表4)。但低光照处理显著降低了叶片的全碳含量(P=0.005,表4)。

表3 3种光照条件下紫背天葵叶片色素含量Table 3 Pigment contents of B. fimbristipula in three different light condition
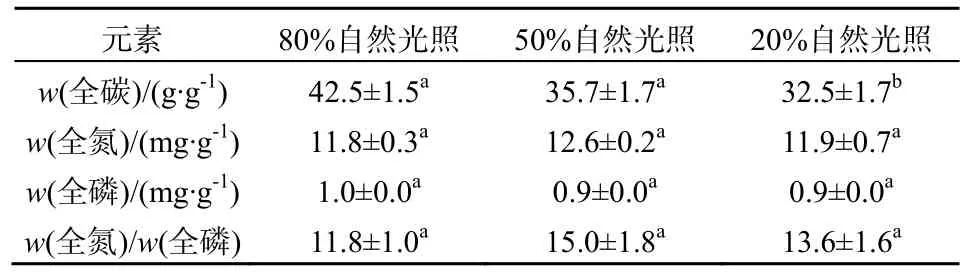
表4 3种光照条件下紫背天葵叶片碳、氮、磷含量及氮/磷特征Table 4 Macronutrient contents and N: P ratio in leaves of B. fimbristipula in three different light conditions

表2 3种光强条件下紫背天葵光和特性及荧光参数Table 2 Photosynthetic traits and chlorophyll fluorescence parameters of B. fimbristipula in three different light condition
3 讨论
3.1光照强度对紫背天葵叶片形态、解剖和生理特征的影响
光对植物的形态特征和生理功能有着重要影响(Deng等,2012)。叶片不仅是紫背天葵主要的药用部位,同时也是其进行光合作用等生理代谢活动的重要器官,栽培在不同光照下的紫背天葵在叶片结构、光合作用特性和代谢物质含量上都表现出一定的差异性。
80% NL的高光照处理下(光合有效辐射较强),紫背天葵叶片接受更多的光能,在生长过程中,叶片通过增大气孔以促进气体交换,同时通过增大下表皮毛密度及表皮覆盖的蜡质来减少水分蒸发及增加叶片抗旱能力(孟雷等,2002;吕晋慧等,2012)。80% NL处理下紫背天葵叶片SLA显著降低,这与高光强下叶片通常降低SLA的研究结果一致,表明其通过调整叶片厚度和叶肉细胞密度而减小SLA这一结构性状,从而有利于自身对光的截获,同时增强叶片对强光的防护作用(张林和罗天祥,2004)。20% NL的低光照处理下叶片通过增加叶面积来增大自身的光接受面积,减少上表皮厚度及叶片厚度以利于光照穿透叶片表皮达到叶肉组织并增强叶片对光的捕获能力,进而缓解光照下降对其光合能力的限制(Deng等,2012;Quero等,2006;Markesteijn等,2007)。植物叶片内叶绿体数量和结构与其光合能力密切相关,低光照处理下紫背天葵叶片降低了其海绵组织、叶绿体个数及基粒片层数,以适应较低光照环境,但较低的光照似乎对叶绿体基粒的发育造成了一定负面影响,致其难以被清晰地辨别,这与Deng等人研究多瓣茉莉长期生长在低光照下损害了叶绿体发育的结果一致(Liang等,2010;Deng等,2012)。
植物生理特性受光照影响显著,与50% NL相比,80% NL下紫背天葵叶片表观量子效率显著减小,20% NL处理下叶片表观量子效率显著增大,表明随着光照降低叶片光量子捕获能力增大,以促进其在弱光环境下对光照的吸收(薛伟等,2011)。此外,研究表明,植物在适宜环境下生长时其表观量子效率通常在0.03~0.05,本研究中3种光照梯度下表观量子效率处于或高于上述范围,表明3种实验条件均适合紫背天葵的生长(Larcher,1980;刘建锋等,2011)。研究表明,光饱和点能反应植物对强光的适应能力,光补偿点则能反应植物对弱光的适应能力(张其德,1992;刘玲等,2009)。80% NL处理下紫背天葵叶片的光饱和点和净光合速率都显著的高于50% NL下叶片的这两个值,表现出其对高光照具较好的利用能力,而20% NL处理显著的降低了紫背天葵叶片光补偿点,则表明其对弱光具有较强的利用能力。同时,与前人的研究结果相似的是,20% NL处理下叶片通过降低暗呼吸速率来减小对光合产物的消耗(Jiang等,2004;孙小玲等,2010)。80% NL下的紫背天葵叶片光系统Ⅱ最大光化学效率(Fv/Fm)与50% NL处理下无显著性差异,表明其在较强光照环境下没有出现光抑制,而显著地大于20% NL下的叶片Fv/Fm则表明较低光照降低了其光系统II潜在光合能力(Demmig 和Adams,2006;刘建锋等,2011)。
遮阴在改变光照强度的同时也改变了光谱组成,一般会使蓝光比例增高。20% NL处理下紫背天葵通过增加其叶片内叶绿素b的含量来提高自身对漫射光中蓝光的利用率。植物体内类胡萝卜素行使光能捕获和光破坏防御两大功能(孙小玲等,2010),与50% NL处理相比,处于80% NL和20% NL处理下叶片的类胡萝卜素含量都要显著的增大,在较强光照下类胡萝卜素有助于叶片将过剩的光能以热量的形式散发出去,对光合器官起保护作用,而在低光照下则可能通过增加叶片内类胡萝卜素的含量来进一步增加对光能的吸收和传递(Liang等,2010;孙小玲等,2010)。在50% NL下的紫背天葵叶片具有相对较低的类胡萝卜素含量及适中的叶绿素含量,说明这一光照强度可能对紫背天葵叶片生长较有利,叶片能充分的利用这一光强下的光能。
次生代谢产物在药用植物体内的合成和积累与所处环境具有紧密的联系,其含量是药材品质的重要保障之一(黄璐琦等,2004)。紫背天葵叶片花青素的含量决定其药用价值,本文研究结果表明,80% NL高光强处理显著降低了叶片内总花青素含量,表明较强光照不利于叶片内花青素的合成和积累,而20% NL低光照处理下花青素含量与对照无显著差异,这可能是由于随着光照的减弱,其叶片光合能力减弱,合成花青素所必须的糖分、蛋白等供应不足从而也限制了植株花青素合成(胡可等,2010)。此外,遮阴在影响光照条件的同时也对叶片所处环境的温度、空气湿度等其他因素产生影响,这些因素也可能间接影响了花青素的合成(Coley和Aide,1989)。
80% NL处理下叶片全碳、全氮、全磷含量与50% NL没有显著性差异,这表明虽然高光强下叶片光合能力提高,但其光合产物积累却没有相应的增加,而20% NL环境下紫背天葵叶片全碳含量则显著降低,光合作用产物是参与初生和次生代谢的必需物质(Croteau,1972;焦晓林和高薇薇,2011),这表明50% NL下叶片具有合适的光合产物积累能力,有利于花青素的合成积累。
3.2紫背天葵叶片对光的适应性及其光照试验实践价值
通过比较生长于不同光照环境下的紫背天葵叶片形态和生理特征,发现其对光照的适应范围较大。紫背天葵作为阴生植物,50% NL下具有典型的阴生植物特性,如较大的比叶面积、表观量子效率,较低的光饱和点和补偿点,而在强光照和低光照环境下,植株的形态和生理特性则出现了适应性差异。较高光照环境下,植株通过调整形态结构以增强抗旱、保水、气体交换和增大光合作用和水分利用能力,能维持自身正常的生长和代谢,但较强的光照显然不利于体内花青素的积累。而处于低光照环境下时,紫背天葵则通过调节叶片厚度和体内色素含量来增加其对弱光的利用和适应能力,同时降低自身光合速率和暗呼吸速率以适应较低光照环境,虽然能维持自身的生长及代谢,但因自身光合能力过低,对花青素的合成产生了一定的限制作用。处于50% NL下的生长的紫背天葵,其叶片内花青素含量及叶片光能利用效率均处于较高水平,适于生长和积累花青素。可见,过高或过低的光照条件均不利于紫背天葵叶片花青素的积累,在紫背天葵的生产栽培中,可参考50%左右自然光照作为其生长的光环境条件,以促进紫背天葵生长及叶片药用成分的积累。
4 结论
本研究表明,不同光照对紫背天葵叶片形态结构、生理指标及次生代谢物质等特征具有一定的影响。与正常光照相比,生存于较高和较低光照处理下的紫背天葵都能通过调节自身形态特征来改善生理功能,从而达到对光照环境的适应。紫背天葵作为典型的阴生植物,对光照有着较强的适应能力,在较高和较低光强下都能正常的维持自身的生长代谢。但作为药用植物,人工生产中对药用有效成分的维持尤为重要。本研究表明在50% NL下,紫背天葵能充分的利用光能,具有较强的光合能力和花青素合成能力,故人工生产上可参考接近自然环境的50% NL条件。但因花青素的合成除了受光照条件的影响,其他诸如温度、空气湿度、糖分等因素对植物花青素的合成也具有一定的影响,因此进一步研究外界环境因子与紫背天葵体内花青素合成的关系,对大规模生产药用有效成分高的紫背天葵具有重要的指导意义,同时也能减少野外红葵采摘现状,缓解红葵生存压力。
参考文献:
COLEY P D, AIDE T M. 1989. Red coloration of tropical young leaves: a possible antifungal defence? [J]. Journal of Tropical Ecology, 5(3): 293-300.
DAI Y J, SHEN Z G, LIU Y, et al. 2009. Effects of shade treatments on the photosynthetic capacity, chlorophyll fluorescence, and chlorophyll content of Tetrastigma hemsleyanum Diels et Gilg [J]. Environmental and Experimental Botany, 65(2-3): 177-18.
DEMMIG-ADAMS B, ADAMS W W. 2006. Photoprotection in an ecological context: the remarkable complexity of thermal energy dissipation [J]. New Phytologist, 172(1): 11-21.
DENG Y M, SHAO Q S, LI C C, et al. 2012. Differential responses of double petal and multi petal jasmine to shading: II. Morphology, anatomy and physiology [J]. Scientia Horticulturae, 144: 19-28.
GRATANI L, COVONE F, LARCHER W. 2006. Leaf plasticity in response to light of three evergreen species of the Mediterranean maquis [J]. Trees, 20(5): 549-558.
JIANG Y, DUNCAN R R, CARROW R N. 2004. Assessment of low light tolerance of seashore paspalum and bermudagrass [J]. Crop Science, 44(2): 587-594.
LARCHER W. 1980. Physiological Plant Ecology [M]. Berlin: Springrt.
LIANG K M, LIN Z F, REN H, et al. 2010. Characteristics of sun- and shade-adapted populations of an endangered plant Primulina tabacum Hance [J]. Photosynthetica, 48(4): 494-506.
LIN Z F, LI S S, LIN G Z, et al. 1984. Superoxide dismutase activity and lipid peroxidation in relation to senescence of rice leaves [J]. Acta Botanica Sinica, 26: 605-15.
LIU N, GUAN L. 2012. Linkages between woody plant proliferation dynamics and plant physiological traits in southwestern North America [J]. Journal of Plant Ecology, 5(4): 407-416.
MARKESTEIJN L, POORTER L, BONGERS F. 2007. Light-dependent leaf trait variation in 43 tropical dry forest tree species [J]. American Journal of Botany, 94(4): 515-525.
MURRAY J R, HACKETT W P. 1991. Dihydroflavonol reductase activity in relation to differential anthocyanin accumulation in juvenile and mature phase Hedera helix L [J]. Plant Physiology, 97(1): 343-351.
NIJS I, FERRIS R, BLUM H. 1997. Stomatal regulation in a changing climate: a field study using free air temperature increase (FATI) and free air CO2enrichment (FACE) [J]. Plant Cell and Environment, 20(8): 1041-1050.
QUERO J L, VILLAR R, MARANON T, et al. 2006. Interactions of drought and shade effects on seedlings of four Quercus species: physiological and structural leaf responses [J]. New Phytologist, 170(4): 819-834.
CROTEAU R, BURBOTT A J, LOOMIS W D. 1972. Apparent Energy Deficiency in Mono-and Sesqui-terpene Biosynthesis in Peppermint [J]. Phytochemistry, 11(10): 2937-2948.
REDDY V S, DASH S, REDDY A R. 1995. Anthocyanin pathway in rice (Oryza sativa L.): identification of a mutant showing dominant inhibition of anthocyanins in leaf and accumulation of proanthocyanins in pericarp [J]. Theoretical and Applied Genetics, 91(2): 301-312.
SKERGET M, KOTNIK P, HADOLIN M, et al. 2005. Phenols, proanthocyanidins, flavones and flavonols in some plant materials and their antioxidant activities [J]. Food Chemistry, 89: 191-198.
SOUZA R P, MACHADO E C, SILVA J A B, et al. 2004. Photosynthetic gas exchange, chlorophyll fluorescence and some associated metabolic changes in cowpea (Vigna unguiculata) during water stress and recovery [J]. Environmental and Experimental Botany, 51(1): 45-56.
段志芳, 付莉, 黄丽华, 等. 2006. 紫背天葵红色素与牛血清白蛋白的作用研究[J]. 中国野生植物资源, 25(1): 50-52.
古粹芝. 1999. 中国植物志第52卷第1分册[M]. 北京: 科学出版社: 190-192.
胡可, 韩科厅, 戴思兰. 2010. 环境因子调控植物花青素苷合成及呈色的机理[J]. 植物学报, 45(3): 307-317.
黄璐琦, 陈美兰, 肖培根. 2004. 中药材道地性研究的现代生物学基础及模式假说[J]. 中国中药杂志, 29(6): 494-496.
焦晓林, 高薇薇. 2011. 环境因子对药用植物三萜皂苷合成影响的研究进展[J]. 42(2): 398-402.
刘光崧. 1996. 土壤理化分析和剖面描述[M]. 北京: 中国标准出版社.
刘建锋, 杨文娟, 江泽平, 等. 2011. 遮荫对濒危植物崖柏光合作用和叶绿素荧光参数的影响[J]. 生态学报, 31(20): 5999-6004.
刘玲, 刘淑明, 孙丙寅. 2009. 不同产地花椒幼苗光合特性研究[J]. 西北农业学报, 18(3): 160-165.
吕晋慧, 王玄, 冯雁梦, 等. 2012. 遮荫对金莲花光合特性和叶片解剖特征的影响[J]. 生态学报, 32(19): 6033-6043.
孟雷, 陈温福, 李磊鑫, 等. 2002. 减弱光照强度对水稻叶片气孔性状的影响[J]. 沈阳农业大学学报, 33(2): 87-89.
邵玲, 陈雄伟, 梁广坚, 等. 2014. 广东肇庆鼎湖地区野生紫背天葵种质资源研究[J]. 中药材, 37(2): 202-206.
邵玲, 梁霞. 2012. 秋海棠科紫背天葵生物学特性[J]. 农学学报, 2(8): 49-52.
孙小玲, 许岳飞, 马鲁沂, 等. 2010. 植株叶片的光和色素构成对遮阴的响应[J]. 植物生态学报, 34(8): 989-999.
覃凤飞, 李强, 崔棹茗, 等. 2012. 越冬期遮阴条件下3个不同秋眠型紫花苜蓿品种叶片解剖结构与其光生态适应性[J]. 植物生态学报, 36(4): 333-345.
谭雄斯, 王景, 唐铁鑫. 2012. 紫背天葵药材质量标准研究[J]. 广东药学院学报, 28(4): 415-418.
王洋, 戴绍军, 阎秀峰. 2004. 光强对喜树幼苗叶片次生代谢产物喜树碱的影响[J]. 生态学报, 24(6): 1118-1122.
邢福武. 2005. 中国的珍稀植物[M]. 长沙: 湖南教育出版社: 107-108.
许旋, 卢忠, 罗一帆. 2000. 不同等级紫背天葵微量元素含量的测定[J].广东微量元素科学, 7(4): 55-57.
薛伟, 李向, 朱军涛, 等. 2011. 遮阴对疏叶骆驼刺叶形态和光合参数的影响[J]. 植物生态学报, 35(1): 82-90.
杨勇, 赖永超, 刘楠, 等. 2010. 艳桢桐的生态生物学特征[J]. 安徽农业科学, 38(19): 10040-10043, 10071.
张兰英, 李耿光. 1986. 影响紫背天葵试管苗花青甙含量的因素Ⅱ[J].云南植物研究, 1: 60-66.
张林, 罗天祥. 2004. 植物叶寿命及其相关叶性状的生态学研究进展[J].植物生态学报, 28(6): 644-852.
张其德. 1992. 大气CO2含量升高对光合作用的影响[J]. 植物通报, 9(4): 18-23.
张治安, 徐克章, 任跃英, 等. 1994. 光照条件对参株碳水化合物和人参皂试含量的影响[J]. 吉林农业大学学报, 16(3): 15-17.
Responses of Leaf Morphological and Physiological Characteristics of Begonia fimbristipula Hance to Light Intensity
WANG Yue1, 2, LIU Nan1, REN Hai1, SHAO Ling3, CHEN Xiongwei3, CHEN Gang3, WANG Jun1
1. South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China; 2. University of Chinese Academy of Sciences, Beijing 100049, China; 3. College of Life Science, Zhaoqing University, Zhaoqing 526061, China
Abstract:Begonia fimbristipula Hance is a unique and endangered medicinal plant species in China. This study was conducted to detect the effects of light intensity on leaf morphological and physiological characteristics, nutrients and secondary metabolite contents of B. fimbristipula, germinated from bulbs, through artificial shade treatments. Specific leaf area and apparent quantum yield of B. fimbristipula leaves decreased with the increment of irradiation, while adaxial epidermis thickness showed the opposite trend. Compared with plants grown under control (50% natural light), plants grown under high irradiation (80% natural light) had higher maximum photosynthetic rate, light saturation point, light compensation point, instantaneous light and water use efficiency, but much lower anthocyanin content in leaves. Plants grown under low irradiation (20% natural light) were lower in chloroplast number in spongy, lamellae per granum, specific lamellae per μm, maximum photosynthetic rate, dark respiration rate, light saturation point, light compensation point, instantaneous water use efficiency and chlorophyll a/b than the plants grown under control. No significant difference in anthocyanin content was observed between plants grown under 20% natural light and control. These results indicated that B. fimbristipula can survive in high or low light condition through acclimatizing its morphological and physiological traits. Respecting to the maintenance of medicinal ingredients in leaves, the 50% natural light level is recommended in the artificial cultivation practices of B. fimbristipula, since plant has better photosynthesis and anthocyanin synthesis abilities (0.4 μmol·g-1) under such light condition.
Key words:Begonia fimbristipula Hance; irradiation; morphological and anatomical traits; photosynthetic physiology; secondary metabolite
收稿日期:2015-04-09
*通信作者:王俊,助理研究员,博士。Email: wxj@scbg.ac.cn
作者简介:汪越(1989年生),女,硕士研究生,主要研究方向为恢复生态学。E-mail: yueyayue@126.com
基金项目:广东省科技计划项目(2013B060400016)
中图分类号:Q945.79
文献标志码:A
文章编号:1674-5906(2015)06-0957-08
DOI:10.16258/j.cnki.1674-5906.2015.06.008
