不同林龄次生林繁殖鸟类群落结构1)
2015-03-10赵欣姜云垒尚伟平姚纪元李时王海涛
赵欣 姜云垒 尚伟平 姚纪元 李时 王海涛
(吉林农业大学,长春,130118) (东北师范大学)
不同林龄次生林繁殖鸟类群落结构1)
赵欣 姜云垒 尚伟平 姚纪元 李时 王海涛
(吉林农业大学,长春,130118) (东北师范大学)
采用样线调查法对40~80 a不同林龄次生林繁殖鸟类群落结构开展了研究。结果表明:随着林龄的增加,次生林繁殖鸟类群落组成有显著差异,不同林龄次生林的优势种不同。不同林龄鸟类群落的Simpson多样性指数差异极显著,Shannon-Wiener多样性指数差异显著,二者均呈现早期上升、中期下降、后期增加并趋于稳定的变化趋势。Pielou均匀度指数差异不显著。40 a次生林与50 a次生林的繁殖鸟类共有种最少,相似性系数最低,70 a次生林与80 a次生林的繁殖鸟类共有种最多,相似性系数最高。
次生林;繁殖鸟类;群落结构
We investigated the breeding bird community structure in five secondary forests with different stand ages (40-80 a) using the line intercept method. The the community species composition varied significantly between five secondary forests with different stand ages, and the dominant species were different. Simpson bird diversity index and Shannon-Wiener bird diversity index in forests with different stand ages differed significantly, which all increased at early stage, then declined at medium stage, and became stable at mature stage. The Pielou evenness index of bird communities in forests with different stand ages did not differ significantly, Pielou evenness index increased rapidly at first, then declined slowly, and reached a maximum at 60 a. There were fewest common species and lowest similarity coefficient between the 40-a forest and the 50-a forest; however, there were the most common species and the highest similarity coefficient between 70-a and 80-a forest.
鸟类群落结构与栖息地的气候、地形、地势及植被类型密切相关,即生态系统的多样性决定了鸟类群落结构的多样性[1]。林龄是划分林分的主要森林结构特征之一,不同林龄的森林林下灌木和草本植物的物种丰富度、均匀度、Simpson指数和Shannon-Wiener指数均存在明显的差异性和波动性[2]。林龄对植物群落结构和动物群落结构都有显著的影响[3]。鸟类群落的种-多度与林地的垂直结构明显相关:林下植物物种丰富度影响冠层取食鸟类和冠层营巢鸟类的物种丰富度;冠层植物物种丰富度影响树干取食鸟类的物种丰富度[4];灌丛植物物种丰富度越高,鸟类物种多样性越高[5]。鸟类的群落结构还与冠层高度、冠层盖度、灌木高度、树高、树胸径和枯木的数量[6-7]等有关。次生林是由于人为破坏性干扰或异常自然干扰,使原始林的林分结构、建群物种组成或基本功能发生了显著变化,经过天然更新或人工诱导天然更新恢复形成的林分[8]。次生林作为鸟类群落研究的载体,已经得到了鸟类学家的广泛关注,但有关不同林龄次生林繁殖鸟类群落结构的研究还相对较少。因此,笔者对不同林龄次生林鸟类群落结构开展研究,探讨是否随着林龄的变化鸟类群落的物种多样性、丰富度和均匀度等群落特征也随之变化。
1 研究地自然概况
选择吉林省延边朝鲜族自治州敦化地区黄泥河(127°51′~128°14′E,43°55′~44°06′N,林龄约40 a)、吉林市昌邑区左家(126°01′~126°08′E,44°00′~44°07′N,林龄约50 a)、吉林市蛟河地区的天岗和旺起(126°35′~128°2′E,42°06′~43°51′N,林龄约60 a)、通化辉南三角龙湾(126°13′~126°32′E,42°16′~42°26′N,林龄约70 a)、吉林市桦甸(126°16′~127°45′E,42°34′~43°29′N,林龄约80 a)的不同林龄次生林为研究区域(图1)。这5个研究地点在植被类型与组成、人为活动和管理上基本相同[9-13]。
2 研究方法
2.1 野外调查
2011—2014年5月下旬至6月上旬,采用无距离样线法[14]对不同林龄次生林繁殖鸟类群落结构进行调查。在每个研究区域内选择5块植被类型相似的生境各布设1条固定样线,共计25条,样线长度为3.0~3.5 km。样线调查时间选择在每年的5月下旬至6月上旬,鸟类调查时间为05:00—09:00和15:00—19:00[15]。调查时以1.5~2.0 km·h-1的速度沿样线前行,记录样线两侧遇见和听到的鸟种类及数量。利用GPS记录航迹,以确保每次调查样线的一致性。每年每条样线重复调查2次,时间间隔为1 d。
图1 次生林繁殖鸟类群落结构研究样点分布
2.2 数据处理与分析
以样线内鸟类调查结果评价各研究区域内鸟类多样性。由于Simpson多样性指数和Shannon-Wiener多样性指数对物种丰富度、均匀度以及优势度的敏感性有所不同[16],所以用2种多样性指数对群落物种多样性进行分析。用Pielou指数进行群落均匀度分析,用Sorensen指数进行群落相似性分析,用Berger-Parker优势度指数对物种优势度进行分析。
①Berger-Parker优势度指数,I=ni/N。式中:I为优势度指数;ni为物种i的个体数量;N为群落中全部物种的总数量。当优势度指数I≥0.1时,定为优势种;0.01≤I<0.10时,定为常见种;I<0.01时,定为稀有种[17-18]。

④Pielou均匀度指数[21],E=H′/lnS。
⑤Sorensen相似性系数[19],S=2c/(a+b)。式中:S为相似性系数;a为群落A中的物种数;b为群落B中的物种数;c为A、B群落中共有的物种数。
用SPSS 21.0进行数据分析。采用One Way-ANOVA Test对不同林龄次生林繁殖鸟类群落组成的差异性进行分析;采用One Way-ANOVA对不同林龄次生林繁殖鸟类Simpson多样性指数、Shannon-Wiener物种多样性指数和Pielou均匀度指数进行比较分析(P=0.05)[2]。
3 结果与分析
3.1 不同林龄次生林繁殖鸟类群落物种组成
不同林龄次生林繁殖鸟类群落物种组成差异极显著(F=20.773,df=4,P=0.003)。40 a次生林的鸟类群落共有9目,22科,58种;50 a次生林里记录到的繁殖鸟类共有12目,32科,74种;60 a次生林的鸟类群落共有9目,23科,61种;70 a次生林的鸟类群落共有9目,25科,72种;80 a次生林的鸟类群落共有12目,29科,73种。不同林龄次生林鸟类群落的优势种不同(表1)。
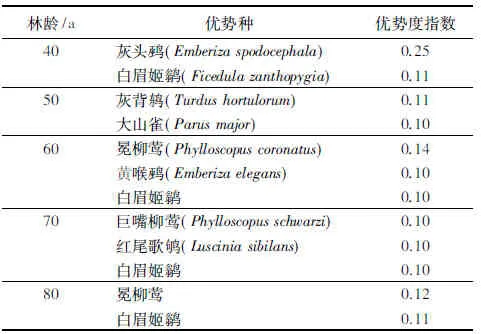
表1 不同林龄次生林繁殖鸟类群落优势种及优势度
3.2 不同林龄次生林繁殖鸟类群落的多样性
随着林龄的增加,不同林龄次生林繁殖鸟类群落的Simpson多样性指数差异极显著(F=23.104,df=4,P=0.001),Shannon-Wiener多样性指数也存在极显著差异(F=4.764,df=4,P=0.011),二者显现出一致的变化趋势。Simpson指数和Shannon-Wiener指数均在40 a时最小,分别为0.910和3.036,在50 a时大幅上升达到最大,分别为0.948和3.356,在60 a时逐渐下降,在70 a和80 a时趋于平缓(表2)。
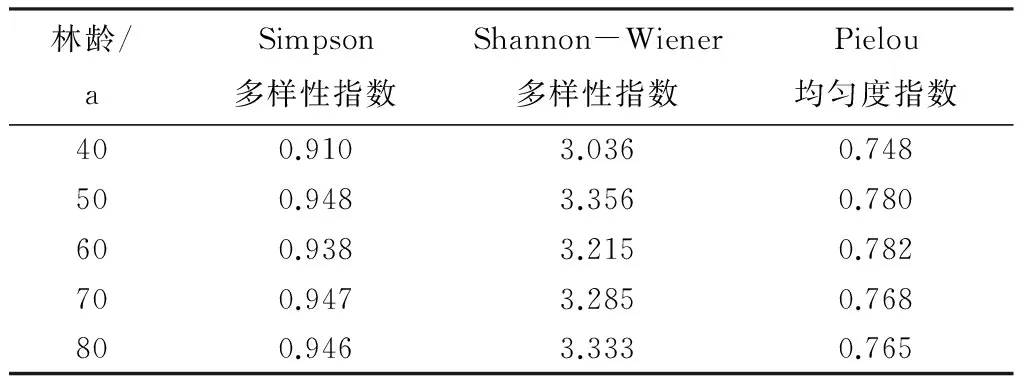
表2 不同林龄次生林鸟类群落多样性指数及均匀度指数
3.3 不同林龄次生林繁殖鸟类群落的均匀度
不同林龄次生林繁殖鸟类Pielou均匀度指数差异不显著(F=2.166,df=4,P=0.123)。不同林龄次生林繁殖鸟类群落的Pielou均匀度指数随着林龄的增加显现出了与多样性指数稍有不同的趋势(表2)。Pielou均匀度指数在40a时最小,为0.748,快速上升到50 a后继续缓慢上升到60 a,在60 a时达到最大值,为0.782,随后呈现缓慢下降并逐渐趋于平缓的趋势。
3.4 不同林龄次生林繁殖鸟类群落的相似性
40 a次生林的繁殖鸟类群落与50 a次生林的繁殖鸟类群落的共有种最少,为42种,相似性系数最低,为0.636。70 a次生林的繁殖鸟类群落与80 a次生林的繁殖鸟类群落的共有种最多,为59种,相似性系数也最高,为0.787(表3)。
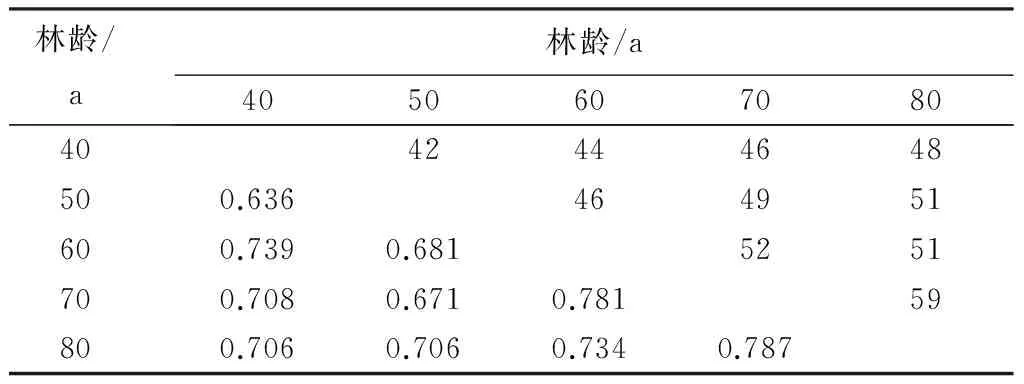
表3 不同林龄次生林繁殖鸟类群落的相似性及共有种
注:左下角为群落相似性系数,右上角为群落共有种数量。
4 结论与讨论
随着林龄的增长,不同林龄次生林繁殖鸟类群落的物种组成、Simpson多样性指数和Shannon-Wiener多样性指数存在相同的变化趋势,均在40 a时最低,大幅度增长到50 a时又迅速下降,从60 a开始稳定而缓慢增长的过程。这与其他学者的研究结果[6-7]基本一致。森林可以为鸟类的生存提供必要的食物和栖息地[22],因而森林演替的不同阶段对鸟类群落结构的变化存在着巨大的影响[23]。在次生林的形成初期,植物资源等鸟类生存所必须的资源相对缺乏,因而鸟类的种类比较少,多样性比较低。但由于初期的次生林存在相对较大的发展空间,所以植物资源会经历迅速发展的阶段,鸟类数量和多样性也随之迅速增加。达到最大值后物种更替变慢,种间竞争加大,鸟类数量和多样性开始下降,最后次生林趋于稳定[24],鸟类数量和多样性也趋于稳定。
均匀度指数反应了个体在各种间分配的均匀性。40 a的次生林均匀度指数最低,说明在40 a的次生林中各个物种的个体数量差异最大,60 a的次生林均匀度指数最高,说明在60a的次生林中各个物种的个体数量差异最小。这与次生林植被空间异质性和生境复杂程度等因素有关[25]。不同梯度上的生境差异,导致了鸟类群落组成的不同,表现出生境空间分布的异质性和复杂性,这种异质性和复杂性导致了鸟类群落物种的多样性[26]。因为食物资源、巢址资源等对鸟类群落结构均有很大的影响[27]。又由于次生林植物群落多样性会经历先上升再下降的发展阶段[28],鸟类群落均匀度也随之产生相应的变化。
次生林形成初期,鸟类种类最少,但随着林龄的增加和林地的成熟,一些喜欢在成熟林中营巢[29]和取食[5]的鸟类开始入住,鸟类丰富度开始增加,共有种也在增加,相似性系数也随之增加,因此40 a和50 a次生林的相似性系数最低,70 a和80 a次生林相似性系数最高。
次生林是植物群落演替过程中的一个重要阶段,它能为鸟类的取食、繁殖、营巢等行为提供必不可少的资源支持。而林龄作为次生林的一个重要林分特征,与林内植被组成和结构层次都显著相关[30-31]。因此,对不同林龄次生林鸟类群落结构进行研究,对次生林的保护和对鸟类群落的保护都有积极的意义。
致谢:感谢相桂权、张立世老师;李玲玉、李振奎、赵殿正、刘珍妮、刘丽秋、陈艳东、樊倩西、王琳同学在野外调查中给予的帮助。
[1] 王斌,马建章.西藏珠穆朗玛峰国家级自然保护区鸟类群落结构与多样性[J].生态学报,2013,33(10):3057-3064.
[2] 崔宁洁,陈小红,刘洋.不同林龄马尾松人工林林下灌木和草本多样性[J].生态学报,2014,34(15):4313-4323.
[3] Moning C, Müller J. Critical forest age thresholds for the diversity of lichens, molluscs and birds in beech (FagussylvaticaL.) dominated forests[J]. Ecological Indicators,2009,9(5):922-932.
[4] Laiolo P. Effects of habitat structure, floral composition and diversity on a forest bird community in north-western Italy[J]. Folia Zoological,2002,51(2):121-128.
[5] Paker Y, Yom-Tov Y, Alon-Mozes T, et al. The effect of plant richness and urban garden structure on bird species richness, diversity and community structure[J]. Landscape and Urban Planning,2014,122:186-195.
[6] Styring A R, Ragai R, Sheldon F H. Bird community assembly in Bornean industrial tree plantations: effects of forest age and structure[J]. Forest Ecology and Management,2011,261(3):531-544.
[7] Nikolov S C. Effect of stand age on bird communities in late-successional Macedonian pine forests in Bulgaria[J]. Forest Ecology and Management,2009,257(2):580-587.
[8] 朱教君,刘世荣.次生林概念与生态干扰度[J].生态学杂志,2007,26(7):1085-1093.
[9] 王海民,林文树.黄泥河林业局森林资源现状与可持续经营对策[J].东北林业大学学报,2013,41(9):119-122.
[10] 王军.左家自然保护区植物群落特征及其演替研究[D].长春:东北师范大学,2007.
[11] 王辉,秦树林.吉林省蛟河林业实验区管理局森林资源现状及可持续经营对策[J].林业勘查设计,2012(2):2-5.
[12] 孙培琦.龙湾自然保护区生态资源特征及其可持续发展[J].吉林林业科技,2004,33(2):23-271.
[13] 桦甸市人民政府.桦甸市资源优势[EB/OL].(2013-05-21)[2015-01-10].http://www.huadian.gov.cn/zsyz/zyys/665.html.[14] 许龙,张正旺,丁长青.样线法在鸟类数量调查中的运用[J].生态学杂志,2003,22(5):127-130.
[15] 蔡音亭,干晓静,马志军.鸟类调查的样线法和样点法比较:以崇明东滩春季盐沼鸟类调查为例[J].生物多样性,2010,18(1):44-49.
[16] 许晴,张放,许中旗.Simpson指数和Shannon-Wiener指数若干特征的分析及“稀释效应”[J].草业科学,2011,28(4):527-531.
[17] 马杰,阎文杰,孙儒泳,等.东灵山辽东栎林啮齿动物群落组成及多样性[J].动物学杂志,2003,38(6):37-41.
[18] 李健,王文.内蒙古红花尔基地区草原—森林生态系统不同生境鸟类多样性[J].东北林业大学学报,2009,37(10):39-43.
[19] 孙儒泳.动物生态学原理[M].北京:北京师范大学出版社,2001.
[20] 冯江,高玮,盛连喜.动物生态学[M].北京:科学出版社,2005:505.
[21] 张金屯.数量生态学[M].北京:科学出版社,2004:91.
[22] Rosenvald R, Lõhmus A, Kraut A, et al. Bird communities in hemiboreal old-growth forests: The roles of food supply, stand structure, and site type[J]. Forest Ecology and Management,2011,262(8):1541-1550.
[23] 范喜顺,胡德夫,陈合志.华北平原耕作区鸟类群落结构与林业生态关系研究[J].干旱区研究,2005,22(2):186-191.
[24] 杨立学,孙跃志.不同林龄胡桃楸林下植物多样性的差异[J].生态学杂志,2013,32(4):807-812.
[25] 徐玲.崇明东滩湿地植被演替不同阶段鸟类群落动态变化的研究[D].上海:华东师范大学,2004.
[26] 姜云垒,高玮,王海涛.长白山北坡鸟类多样性研究Ⅱ:鸟类群落组成随海拔梯度变化[J].吉林农业大学学报,2006,28(3):265-271.
[27] 王海涛.鸟类群落结构形成的因素分析[D].长春:东北师范大学,2003.
[28] 王春香.晋西黄土区人工林下植物多样性与土壤养分研究[D].北京:北京林业大学,2014.
[29] Hobson K A, Bayne E. The effects of stand age on avian communities in aspen-dominated forests of central Saskatchewan, Canada[J]. Forest Ecology and Management,2000,136(1):121-134.
[30] 乔秀娟,曹敏,林华.西双版纳不同林龄次生植物群落优势树种的热值[J].植物生态学报,2007,31(2):326-332.
[31] 刘彤,胡丹,魏晓雪,等.红松人工林林下植物物种多样性分析[J].东北林业大学学报,2010,38(5):28-29.
Community Structure of Breeding Bird in Secondary Forests with Different Stand Ages
Zhao Xin, Jiang Yunlei, Shang Weiping, Yao Jiyuan, Li Shi(Jilin Agricultural University, Changchun 130118, P. R. China); Wang Haitao(Northeast Normal University)//Journal of Northeast Forestry University,2015,43(7):120-122,126.
Secondary forest; Breeding bird; Community structure
1)国家自然科学基金项目(31270471)和生物多样性示范观测—吉林省7个观测样区项目。
赵欣,女,1990年4月生,吉林农业大学生命科学学院,硕士研究生。E-mail:xingfei809813@163.com。
姜云垒,吉林农业大学动物科学技术学院,教授。E-mail:jiangyl487@nenu.edu.cn。
2015年1月27日。
Q958.1
责任编辑:程 红。
