4种次级洞巢鸟巢资源竞争对其繁殖优势度的影响1)
2015-03-08尚伟平姜云垒赵欣李时姚纪元邓秋香
尚伟平 姜云垒 赵欣 李时 姚纪元 邓秋香
(吉林农业大学,长春,130118) (北华大学)
责任编辑:程 红。
次级洞巢鸟是鸟类群落的重要组成部分[1]。其繁殖期活动都是从竞争有限的洞巢资源开始的[2],巢洞作为一种有限的资源[3],其数量直接影响着次级洞巢鸟的群落结构以及种群数量[4]。巢址资源对次级洞巢鸟繁殖的限制程度取决于其可利用巢址的数量以及他们对于巢洞的需求程度,而可利用巢址的数量受巢洞的形成率、损失率、次级利用者的竞争以及它们之间的领域性等因素的影响[5-7]。
次级洞巢鸟是鸟类群落的重要组成部分[1],其繁殖期活动都是从竞争有限的洞巢资源开始的[2]。巢洞作为一种有限的资源[3],是影响次级洞巢鸟群落结构以及种群数量的关键因素[4]。优势度假说认为在巢洞资源紧缺的情况下,劣势种的种群密度将下降,其原因是由于优势种占据了更多的巢洞资源[5-6];巢利用可塑性假说认为在适宜巢资源有限时,对巢洞类型选择严格的物种仅有较少的巢洞供其利用,其种群数量将下降,而对巢选择可塑性高的物种可利用更多类型的巢洞,能更好地适应不同类型巢资源的变化以及种内的竞争[6]。另外,物种之间的相互作用也可以影响次级洞巢鸟的群落结构及种群数量[7]。巢资源减少时,种间竞争加剧,如紫翅椋鸟(Sturnus vulgaris)将山蓝鸲(Sialia currucoides)、普通币鸟(Sitta europaea)将大山雀(Parus major)从其已占据的巢洞中驱逐出去而篡夺其巢,从而限制了其他鸟类种群数量的增加[4,7-8]。目前,虽然已对巢资源变化和种间竞争对次级洞巢鸟群落结构的影响方面展开了大量研究,但关于巢竞争与群落中物种的繁殖优势度的关系尚不十分清楚。
大山雀、白眉姬鹟(Ficedula zanthopygia)、普通币鸟和沼泽山雀(Parus palustris)为左家自然保护区内繁殖数量占有优势的小型次级洞巢鸟。本研究选择这4 种鸟为研究对象,通过分析其巢洞因子的生态位宽度,探讨巢竞争与4 种次级洞巢鸟类在群落中繁殖优势度的关系。
1 研究地概况
研究区域为吉林省左家自然保护区(126°00'~126°09'E,44°01'~44°06'N)。该区域是长白山亚区向西部平原过渡的丘陵地带,海拔210~540 m,主要林型为次生阔叶混交林,林龄一般为50~60 a。主要树种包括糠椴(Tilia mandshuria)、白皮柳(Salix pierrotii)、蒙古栎(Quercus mongolica)、水曲柳(Fraxinus mandshurica)、槐(Sophara faponica)、黑桦(Betula dahurica)、山杨(Populus davidiana)、春榆(Ulmus davidiana var.japonica)、樟子松(Pinus sylvestris var.mongolica)、黄花落叶松(Larix olgensis)等14 种乔木。树高16~21 m,树冠高5~8 m,胸径11~40 cm。灌木以丁香(Syringa spp.)、刺玫(Rosa davurica)、榛(Corylus heterophylla)为主,草本植物主要以禾本科(Poaceae)和菊科(Asteraceae)为主。研究区域内共有鸟类190 种,繁殖鸟类113 种,非繁殖鸟类77种[9]。通过对天然洞调查发现洞巢鸟共213 巢,其中大山雀、普通币鸟、沼泽山雀以及白眉姬鹟共171巢,占总数量的80.3%。
2 研究方法
2.1 调查方法
2013—2014年3—7月份,在研究区域内选择了6 块生境结构相似,植被生长状况良好的样地(图1),①~⑥号样地的面积分别为13.3、26.1、14.4、39.5、32.9、22.7 hm2。采用直接搜寻法查找确定可被次级洞巢鸟利用的适宜巢洞,对其进行标记、GPS定位。进入繁殖期后,隔日监测巢洞是否被利用。研究中借助管道窥探器(诺泰实业,NTS200)观察巢洞内状况。判断入住的标准为:筑巢期雌雄个体多次叼巢材进入巢洞;巢洞内出现新巢材、卵和雏鸟;或观察到亲鸟育雏。繁殖期结束后,利用游标卡尺、卷尺、罗盘和激光测距仪等对巢址选择变量进行测量。测量指标包括:洞口横径、洞口纵径、洞内径、洞深、巢口距地高、巢位径以及树胸径[8]。同时,记录巢洞类型:根据树洞的形成原因将巢洞类型分为结洞、裂洞及啄洞[10]。
人工巢箱的调查:人工巢箱采用内部规格为25 cm×12 cm×12 cm 的山雀式木板箱以及招引红隼(Falco tinnunculus)用的50 cm×35 cm×35 cm 的大巢箱[11-12],每年的3月初对试验样地内的巢箱进行添加以及更换。隔日对人工巢箱的利用状况进行监测,记录巢箱内情况,通过巢箱内巢材、以及进出巢箱的鸟类、巢箱内的鸟卵情况、雏鸟情况判断巢箱内的入住情况,采用与天然树洞相同的方法测量其巢变量。
2.2 数据处理
应用Berger-Parker 优势度指数[13]计算4 种次级洞巢鸟的种群优势度。其公式为,

式中:D 为优势度指数;Nmax为优势种的个体数;N为功能团全部物种的个体数。
利用卡方检验对4 种次级洞巢鸟类对巢洞类型的使用进行分析。
应用椭圆形面积公式计算巢洞口表面积,利用圆柱形体积公式计算巢洞内部的体积[14],公式分别为S=πab、V=πr2h。式中:S 为巢洞口面积;a 为洞口横径/2;b 为洞口纵径/2;V 为巢洞体积;r 为巢洞内径/2;h 为巢洞深度。

图1 次级洞巢鸟巢资源研究样地分布
应用香浓-威纳多样性指数[15]计算其巢洞因子的生态位宽度。公式为,
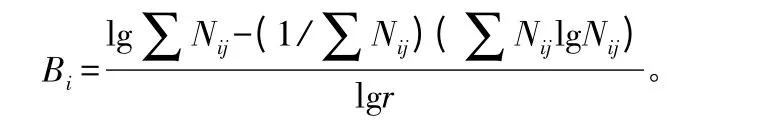
式中:Bi为i 种的生态位宽度;Nij为i 种利用j 资源等级的数值;r 为生态位的资源等级数。
将巢洞因子的生态位宽度进行主成分分析,确定可代表其生态位宽度的巢洞因子,进行生态位重叠的计算,并将巢洞因子的生态位宽度与其群落优势度进行Pearson 相关分析。
使用测量比例重叠的生态位重叠指数[16]进行生态位重叠指数计算,其公式为,

式中:Cih为i 物种和h 物种之间的生态位重叠指数;Nij为i 物种在j 资源等级中出现的数值;Ni为i 物种在所有资源等级中的数值;Nhj为h 物种在j 资源等级中出现的数值;Nh为h 物种在所有资源等级中的数值。
采用Pearson 相关对香浓-威纳为基础的生态位宽度指数以及Berger-Parker 优势度指数进行分析。
以上数据均使用Excel 和SPSS 21.0 完成。
3 结果与分析
3.1 次级洞巢鸟的繁殖优势度
大山雀、普通䴓、白眉姬鹟、沼泽山雀4 种次级洞巢鸟的繁殖种群数量分别为88、87、65、27 巢。大山雀的优势度最大(0.330),其后依次为普通币鸟(0.326)、白眉姬鹟(0.243)、沼泽山雀(0.101)。
3.2 次级洞巢鸟巢洞因子选择的主成分分析
4 种次级洞巢鸟巢洞因子生态位宽度的第一主成分由洞口面积、巢距地高度以及巢树胸径组成,其特征值为3.488,贡献率为49.82%(表1)。因此,选择这3 种因子的生态位宽度代表所有巢洞因子的生态位宽度。

表1 4 种次级洞巢鸟巢洞因子生态位宽度主成分分析
3.3 巢洞因子的生态位宽度
大山雀的洞口面积的生态位宽度(0.632),其后依次是普通币鸟(0.601)、白眉姬鹟(0.589)以及沼泽山雀(0.427);大山雀的巢距地高的生态位宽度最宽(0.811),其后依次为普通币鸟(0.785)、白眉姬鹟(0.646)以及沼泽山雀(0.571);大山雀的巢树胸径的生态位最宽(0.664),其后依次是普通币鸟(0.624)、白眉姬鹟(0.558)以及沼泽山雀(0.416)(表2)。

表2 巢洞因子生态位宽度与繁殖优势度的相关性
3.4 巢洞类型和巢洞因子的生态位重叠
对巢洞的洞口面积、巢距地高、巢树胸径以及巢洞类型4 种因子进行生态位重叠分析(表3)。大山雀与白眉姬鹟巢洞面积生态位重叠最大(0.855),沼泽山雀与普通币鸟重叠最小(0.637);普通币鸟与大山雀巢距地高生态位重叠最大(0.726),沼泽山雀与白眉姬鹟重叠最小(0.298);大山雀与白眉姬鹟选择的巢树胸径生态位重叠最大(0.921),大山雀与沼泽山雀重叠最小(0.744);大山雀与白眉姬鹟巢洞类型生态位重叠最大(0.782),沼泽山雀与白眉姬鹟、沼泽山雀与大山雀的重叠最小(0.379)。结果表明:4 种次级洞巢鸟对4 种巢洞因子的选择利用可能存在着较强的竞争。

表3 4 种次级洞巢鸟巢洞因子生态位重叠
3.5 4 种次级洞巢鸟巢洞类型的选择
大山雀对巢洞资源的利用存在极显著差异(χ2=99.615,P<0.01),主要利用结洞、啄洞、裂洞、大巢箱、小巢箱,甚至电线杆顶端的空隙和石缝等;普通䴓对巢洞资源的利用存在显著差异(χ2=7.724,P<0.05),可利用啄洞、结洞和小巢箱,但不用裂洞;白眉姬鹟对巢洞资源的利用存在极显著差异(χ2=22.077,P<0.01),可利用结洞、啄洞和小巢箱;沼泽山雀对巢洞资源的利用存在极显著差异(χ2=37.714,P<0.01),可利用结洞和裂洞,但极少利用巢箱和啄洞。
3.6 巢洞因子生态位宽度与繁殖优势度
大山雀、普通币鸟、白眉姬鹟和沼泽山雀的洞口面积生态位宽度与其繁殖优势度呈显著正相关(R=0.964,P=0.036);巢距地高的生态位宽度与其繁殖优势度呈显著正相关(R=0.952,P=0.048);巢树胸径的生态位宽度与其繁殖优势度呈极显著正相关(R=0.991,P=0.009)。
4 结论与讨论
生态位研究是近代理论生态学的重要内容,种的生态位宽度和种间生态位重叠被认为是物种多样性及群落结构的决定因素[17]。生态位宽度越大的物种,其对环境的适应能力越强[18],可以适应较激烈的种间竞争,从而增加其繁殖优势度。对于存在竞争的两个物种,如果攻击性较强物种的生态位较窄,竞争力小的物种可能就会通过增加生态位宽度以适应种间竞争[14]。本研究所涉及的4 种次级洞巢鸟中,普通䴓与大山雀都偏爱啄木鸟的旧洞,且繁殖时间存在重叠,种间竞争较激烈[8,19],普通䴓竞争力较强,其选择巢洞类型的生态位较窄[8],竞争结果对大山雀不利。本研究发现,大山雀在巢洞类型的利用以及巢洞因子的生态位宽度均高于普通䴓,表明大山雀可能通过增加生态位宽度来应对与普通䴓的种间竞争,导致其繁殖优势度也较普通䴓略高。沼泽山雀只选择裂洞和结洞[8],巢洞因子生态位最窄,与大山雀和普通䴓在这2 种类型巢洞利用上存在竞争,但其个体较小,且繁殖时间相对晚,在种间竞争中不占优势,导致其繁殖优势度最低。本研究所涉及的4 种次级洞巢鸟中,白眉姬鹟为夏候鸟,繁殖时间较晚,5月中下旬进行繁殖[20],其竞争压力较大,它们选择的巢洞因子生态位较大山雀与普通䴓小、较沼泽山雀大,因此,可能是导致其繁殖优势度较大山雀与普通䴓小、较沼泽山雀大的原因之一。
当然,这4 种次级洞巢鸟对巢资源的竞争结果可能还受其它因素的共同影响,如研究区域内可利用的巢洞资源量等,这些尚需在今后的研究中进一步深入探讨。
致谢:感谢樊倩西、崔响、李玲玉、李振奎、赵殿正、刘珍妮同学在野外调查中给予的帮助,感谢东北师范大学王海涛教授对本论文提出的宝贵意见。
[1] Newton I.The role of nest sites in limiting the numbers of holenesting birds:a review[J].Biological Conservation,1994,70(3):265-276.
[2] Minot E O,Perrins C M.Interspecific interference competitionnest sites for blue and great tits[J].The Journal of Animal Ecology,1986,55(1):331-350.
[3] Politi N,Hunter M Jr,Rivera L.Availability of cavities for avian cavity nesters in selectively logged subtropical montane forests of the Andes[J].Forest Ecology and Management,2010,260(5):893-906.
[4] Robles H,Ciudad C,Matthysen E.Tree-cavity occurrence,cavity occupation and reproductive performance of secondary cavity-nesting birds in oak forests:The role of traditional management practices[J].Forest Ecology and Management,2011,261(8):1428-1435.
[5] Strubbe D,Matthysen E.Experimental evidence for nest-site competition between invasive ring-necked parakeets (Psittacula krameri)and native nuthatches (Sitta europaea)[J].Biological Conservation,2009,142(8):1588-1594.
[6] Aitken K E H,Martin K.Resource selection plasticity and community responses to experimental reduction of a critical resource[J].Ecology,2008,89(4):971-980.
[7] Blanc L A,Walters J R.Cavity excavation and enlargement as mechanisms for indirect interactions in an avian community[J].Ecology,2008,89(2):506-514.
[8] 王海涛,高玮.次级洞巢鸟对次生林天然树洞的利用[J].动物学研究,2002,23(2):136-140.
[9] 孙寒梅,高玮,宫亮,等.吉林省左家自然保护区鸟类组成及其多样性研究[J].东北师大学报:自然科学版,2008,40(1):100-110.
[10] 高玮,王海涛,王日昕.中国东北地区洞巢鸟类生态学[M].长春:吉林科学技术出版社,2004.
[11] 宋杰.人工巢箱的制作和使用[J].生物学通报,1994,29(4):14-17.
[12] Aviles J M,Sanchez J M,Parejo D.Nest-boxes used by Eurasian kestrels Falco tinnunculus are preferred by rollers Coracias garrulus[J].Folia Zoologica,2001,50(4):317-320.
[13] Magurran A E.Ecological diversity and its measurement[M].Princeton:Princeton University Press,1988.
[14] Martin K,Aitken K E H,Wiebe K L.Nest sites and nest webs for cavity-nesting communities in interior British Columbia,Canada:nest characteristics and niche partitioning[J].The Condor,2004,106(1):5-19.
[15] 孙儒泳.动物生态学原理[M].北京:北京师范大学出版社,2001.
[16] Colwell R K,Futuyma D J.On the measurement of niche breadth and overlap[J].Ecology,1971,52(4):567-576.
[17] 李军玲,张金屯.太行山中段植物群落优势种生态位研究[J].植物研究,2006,26(2):156-162.
[18] 林思祖,黄世国,洪伟,等.杉阔混交林主要种群多维生态位特征[J].生态学报,2002,22(6):962-968.
[19] 易国栋,杨志杰,王海涛,等.普通币鸟的巢位选择和繁殖成功巢特征[J].动物学杂志,2005,39(6):23-26.
[20] 魏忠民,姜云垒,张立世,等.人工巢箱条件下白眉姬鹟育雏期双亲投入[J].东北林业大学学报,2012,40(6):141-143.
