闽江口琅岐岛朴树防护林种子雨时空分布特征1)
2015-03-10闫淑君陈莹陈英范海兰林勇明蔡园园陈梦依雷少飞
闫淑君 陈莹 陈英 范海兰 林勇明 蔡园园 陈梦依 雷少飞
(福建农林大学,福州,350002) (西安交通大学)
闽江口琅岐岛朴树防护林种子雨时空分布特征1)
闫淑君 陈莹 陈英 范海兰 林勇明 蔡园园 陈梦依 雷少飞
(福建农林大学,福州,350002) (西安交通大学)
以闽江口琅岐岛天然朴树防护林林下、林隙2种生境为对象,采用离地收集种子的方法,连续定位观测了朴树种子雨的数量动态。结果表明:朴树种群种子雨是从9月下旬开始,一直到次年的1月结束;林下的种子雨总量和完好种子比例,均极显著高于林隙内(P<0.01);林隙内种子雨量具有时空异质性特征,这对林隙内朴树更新格局产生深刻影响。总体上,林隙内南北向的种子雨,高于东西向的。林下与林隙内,朴树种子雨高峰期有所不同,林隙内种子雨高峰期在9月下旬到10月中旬;而林下的是在11月下旬到12月中旬,林下的种子雨在9月下旬到10月中旬也有一个小高峰。林隙内种子雨主要受天气因素影响;而林下的种子雨既受朴树生物学特性影响,又受天气因素影响。朴树在林隙内主要采用实生更新,不是因为种子源比林下多,而是林隙内环境更适合其种子萌发和幼苗生长。
种子雨;林隙;时空分布;朴树;闽江口
We studied the temporal and spatial distribution of the seed rain ofCeltissinensispopulation to reveal the mechanism of natural regeneration of the population in Langqi Island of Minjiang Estuary. We selected two kinds of micro-habitats (understory and forest gap) with the seed rain quantity ofC.sinensispopulation. The seed rain lasted from late September to the end of the following January, the total seed rain number and proportion of intact seeds in understory were significantly higher than those of forest gaps (P<0.01), seed rain in gaps had temporal and spatial heterogeneity characteristics, and it would affect regeneration pattern ofC.sinensispopulation significantly. The order of the amount of seeds in gap was 3>5>4>2>1>9>7>8>6, the difference between the total seed rain number and proportion of intact seeds in the third traps reached the extremely significant than the sixth, seventh, eighth and ninth sample plots (P<0.01). The total seed rain number was higher in north-south than east-west in gap. There were differences about the peak time of seed rain between understory and forest gap, the seed rain reached the peak in late September to mid-October in gap, as well as in late November to mid-December, and late September to mid-October in understory. The seed rain in gap was mainly affected by weather factors (wind, rain), and the seed rain in understory was influenced by the biological characteristics ofC.sinensisand the weather factor. Seedlings ofC.sinensiswere dominated in gaps, and the reason was the environment was more suitable for the seed germination and seedling growth, rather than the seed number in gap were larger than that in understory.
种子作为植物有性繁殖的重要器官,它散布的空间格局对其萌发、幼苗存活和生长等一系列生态过程产生决定性的影响,决定着幼苗在群落中能够成功定居的范围。由于种子散布能够改变植物种群的大小,乃至建立新的种群,作为基因流的表现形式之一,种子散布不仅影响着种群的生物学特征,而且关系到种群的遗传结构,从而对种群的适应性、生活史特征的演化产生重要作用[1]。在特定时间和特定空间从母株上散落的一定数量的种子称为种子雨。种子雨中既有成熟种子,又有不成熟种子,还有死亡的种子。种子雨是植物体将其有性繁殖体(种子或果实)从母树向地表扩散的过程,是森林群落更新繁殖体的主要来源,是植物生命史动态过程中一个不可缺少的环节[2-3],对于它的了解,可以预测土壤种子库的物种组成和大小以及植被的更新演替的趋势。
由于林木自身特点和生态环境异质性,种子雨在发生时间、雨量、强度及散布特征等方面存在很大差异[4]。种子雨时空分配,影响着群落的物种组成、单个种群的变化以及整个群落的格局和过程[5]。
林隙形成后,改变了群落的垂直结构,种子雨的传播阻力减少,因此,会有更多的种子落入土壤中[6-7]。林隙更新过程中种子雨的输入、种子萌发、幼苗生长等,是森林动态循环的重要组成部分和关键环节。因此,研究林隙种子雨的动态变化,对提示森林更新的格局与过程具有重要意义[8]。但是,关于林隙内种子雨动态研究国内相关报道较少[9-10],对于沿海防护林种子雨动态的研究更是缺乏。
朴树(Celtissinensis)为榆科朴属植物,落叶阔叶树种。产于陕西、河南以南至华南,东至台湾,西至四川、云南;花期4月,果熟期10月。核果近球形,熟时橙红色,核果表面有凹点及棱背,种子的直径为3.5~4.5 mm,种长为3.6~6.6 mm[11]。位于琅岐岛的云龙村,村旁风成沙丘上的天然朴树林,距今已有400多年历史,古朴树木参天蔽日,纵横交错;林中乔木高大,朴树幼苗多并长势良好,适宜天然更新,且林下植被较木麻黄(Casuarinaequisetifolia)林下丰富,群落比较稳定。以往对琅岐朴树防护林的研究主要集中在朴树种子特征、朴树种群结构、空间格局、幼苗更新等方面[12-16],而对朴树种群种子雨的研究尚未见报道。
根据对朴树种群幼苗更新调查,林隙内朴树幼苗的密度大于林下,林隙内以实生更新为主,林下以萌生更新为主[16];那么,在朴树林中的林隙内,是不是林隙内朴树种子密度高于林下,导致林隙内实生苗多于林下?这是研究朴树采用不同更新方式进行更新的前提,需要进一步的验证。因此,本文对闽江入海口琅岐岛朴树防护林林隙种子雨的数量特征、动态变化等进行研究,为进一步揭示朴树种群更新机制提供科学依据。
1 研究区概况
研究区位于闽江入海处琅岐岛(119°32′22″E~l19°39′15″E,26°03′30″N~26°07′15″N)。琅岐岛自然条件得天独厚,全岛总面积92 km2,森林覆盖率27.6%,滩涂面积37 km2,海岸线长30 km。本区属于亚热带海洋性季风气候,四季温和,年均气温19.5 ℃,无霜期达360 d,雨量充沛,年均降水量1 600 mm,且多集中在3—7月。地带性植被为亚热带常绿阔叶林,朴树防护林位于琅岐岛的东南部,直面海域,时常受海风的干扰;台风天气集中在7—9月,常年主导风向为东南风。
天然朴树防护林总面积有10 hm2,乔木层植被为以朴树为主的单优种群群落,其群落东部外围分布着少量木麻黄(Casuarinaequisetifolia);灌木层植物有蔓胡颓子(Elaeagnuspungens)、黑面神(Breyniafruticosa)、檵木(Loropetalumchinense)、光叶海桐(Pittosporumglabratum)和东南紫金牛(Ardisiasieboldii)等;草本层植物主要有麦冬(Ophiopogonjaponicus)、肖梵天花(Urenalobata)、白花丹(Plumbagozeylanica)、刺蒴麻(Triumfettarhomboidea)、丁香蓼(Ludwigiaprostrata)和天门冬(Radixasparagi)等;藤本植物有鸡矢藤(Paederiascandens)、圆叶菝葜(Smilaxbauhinioides)和雀梅藤(Sageretiathea)等;附生植物有槲蕨(Drynariaroosii)。土壤属滨海风沙土,是在风成沙性母质上发育的,成土过程十分微弱,发育阶段相对年幼;土壤十分贫瘠,渗漏性强,保水保肥能力较差,养分质量分数低。
2 研究方法
在琅岐朴树天然林林隙内,在东西南北中5个方位设置种子收集框(如图1)。其中:种子雨收集样方1、4、5、8为林隙边缘区,2、3、6、7为林隙近心区,9为林隙中心区。选择3个林隙,共设置27个种子收集框,同样在林下也随机设置27个种子收集框。收集框面积为1 m×1 m,框口四边用木条围成,尼龙网(1 mm×1 mm)做网底;为了避免种子被小动物(特别是海鸟)采食,在框口上面增加一个铁丝网,网眼大小为15 mm×15 mm;在种子雨收集器尼龙网网底开一个口,缝上拉链,每次从网底收集种子。

图1 林隙内种子雨样地分布
朴树果熟期为10月,本研究于2011年8月20日放置种子雨收集框,每隔1个月(每月的20日)收集一次,直到种子下落结束。在试验范围内,以林下朴树种子雨月总量首次超过30粒·m-2为朴树种子雨开始的标志,当种子雨月总量小于30粒·m-2时,即认为种子雨结束。
从林隙和林下收集到的种子中,挑选出朴树种子。将朴树种子按完好种子和非完好种子进行分类,种子颗粒饱满、完整为完好种子,而霉烂种子(用手轻压即破碎)和空粒种子(因动物及虫害破坏形成)归为非完好种子。
所有数据采用SPSS软件进行处理和分析;采用单因子方差分析和最小显著差异法,比较不同数据间的差异显著性。
3 结果与分析
3.1 林隙与林下朴树种子雨大小
琅岐岛朴树防护林林下与林隙内种子雨数量(见表1)。通过对林下与林隙内种子雨量进行方差分析及多重比较,表明:林下与林隙内种子雨总量、完好种子和非完好种子之间的差异,均达到极显著水平(P<0.01);尽管林隙内的种子与林下相比较少,但在外界作用(海风、海鸟等)下,还是有59.6粒·m-2的完好种子量,这为朴树在林隙内进行实生更新提供了种源。
(3)《唐虞之道》简19:“方在下位,不以匹夫为坙(轻),及其有天下也,不以天下为重。”(坙,见母耕部;轻,溪母耕部。)

表1 林下与林隙内朴树种子雨及组成
注:表中数值为“平均值±标准差”;同列不同大写字母表示差异达极显著水平(P<0.01)。
3.2 林隙内种子雨空间分布
林下与母树较近,种子一般分布于母树周围,由于朴树防护林是以朴树为单优势种群的群落,个体间的距离较近,很难确定种子的母树个体,所以,没有对朴树种子在母树周围空间分布进行分析。在林隙内,不同的方位种子雨是否存在差异呢?通过对林隙内不同方位种子雨的收集和统计,结果见表2。对林隙内不同方位种子雨量的方差分析及多重比较,结果表明:林隙内,朴树种子雨在空间上的分布存在异质性,且有的收集样方之间的差异达到极显著水平。在林隙内,种子雨总量由大到小的样方顺序为:3、5、4、2、1、9、7、8、6;林隙内,样方3的种子雨量与样方6、7、8、9差异均达到极显著水平(P<0.01),与收集样方1、2、4、5差异均不显著;林隙内完好种子的空间分布由大到小的样方顺序为:3、2、1、5、4、9、7、8、6,种子雨收集样方4、5尽管种子总量比较高,但是完好种子少于样方1和2,样方3的完好种子量与样方4、6、7、8、9的差异达到极显著水平(P<0.01)。
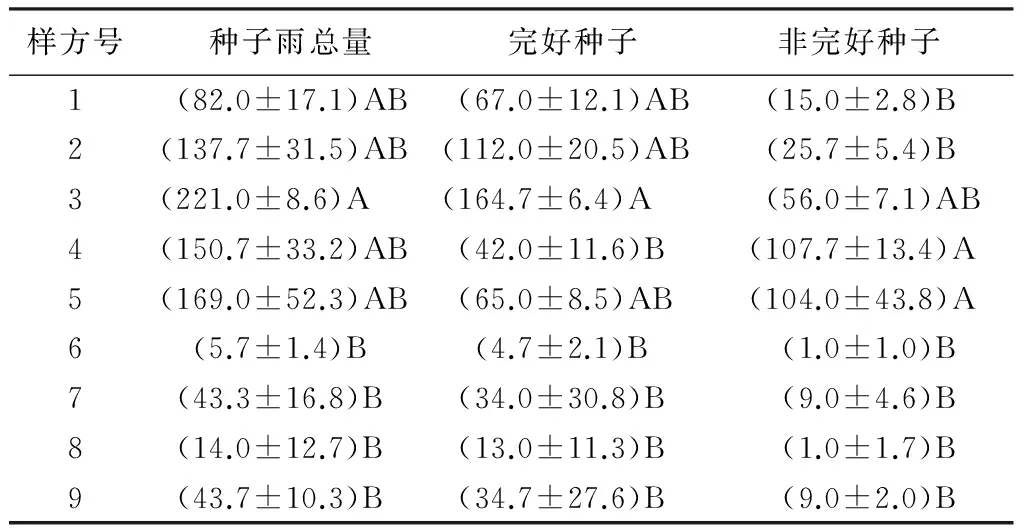
表2 林隙内朴树种子雨空间分布 粒·m-2
注:表中数值为“平均值±标准差”;同列不同大写字母表示差异达极显著水平(P<0.01)。
总体上,林隙内南北方向的种子雨大于东西向,西半部的种子雨大于东半部的;这种分布状态正好说明了林隙内的种子多是由于风的作用掉落,由于本研究区正好位于琅岐岛东南角,风向多为东南风,朴树种子重量又较轻,在风的作用下,朴树种子飘落在林隙内,为朴树在林隙内更新提供了种源。
3.3 林下与林隙内种子雨动态
由表3可知,朴树种子成熟期是10月,林隙与林下种子雨始末期都是从9月下旬开始至翌年1月结束,而且都有2个峰值,但是也存在着差异。

表3 林下与林隙内种子雨动态 粒·m-2
注:表中数值为“平均值±标准差”;同列不同大写字母表示差异达极显著水平(P<0.01)。
林下的种子雨高峰是11月下旬到12月中旬,但在9月下旬到10月中旬也出现了一次峰值;一般地,种子雨的峰值只有一个[17],但朴树种子雨出现了两个峰值,这主要是由于天气因素造成的,2011年9月底到10月初期间,研究区出现连续降雨的天气并伴有微风,本研究区所处位置受到海风的影响较大,导致种子脱落;从收集到的种子成熟程度来说,这时期的种子多为非完好种子,而11月下旬到12月中旬期间,种子多为完好种子,这时期是朴树种子成熟脱落期;因此,朴树种子雨两个峰值形成的原因不同,一个是环境因素形成的,另一个是生物学特性决定的。通过对种子雨量的方差分析及多重比较,从量上看,11月20日—12月20日期间,种子雨量最大,但与9月20日—10月20日、10月20日—11月20日期间种子雨量差异不明显,这三个时期的种子雨量均与12月20日—次年1月10日期间的呈极显著差异。
3.4 林隙内不同方位种子雨动态
从林隙内不同方位样地的种子雨动态变化来看(见表4)。不同样地在同一时间段内的种子雨量有所差异,在9月20日—10月20日、10月20日—11月20日和11月20日—12月20日三个时期,种子雨量差异性达到极显著,12月20日—2012年1月20日期间各样地种子雨量差异不显著;总体上,林隙内种子雨量高峰期是在9月20日—10月20日,第二个小高峰期11月20日—12月20日。
9月20日—10月20日期间,种子雨收集样方3、4和5种子雨量较多,且3和4的种子雨量均与1、2、6、7、8和9的差异达到极显著水平;10月20日—11月20日期间,种子雨收集样方2的种子雨量与样方1、4、6、7、8和9的差异达到极显著水平;11月20日—12月20日期间,收集样方2和3的种子雨量与6、7、8和9的差异达到极显著水平,而收集样方1、2、3、4和5之间种子雨量差异不显著;12月20日—2012年1月20日期间,种子雨处于结束时期,各种子雨收集样方种子雨量都较小,且差异不显著。

表4 林隙内种子雨的空间分布 粒·m-2
注:表中数值为“平均值±标准差”;同列不同大写字母表示差异达到极显著水平(P<0.01)。
4 结论与讨论
通过对朴树种子雨的时间动态的研究可知,林下与林隙内种子雨的变化有所不同,林下的种子雨高峰是11月下旬到12月中旬,但在9月下旬到10月中旬也出现了一次峰值。朴树种子雨出现两个峰值,这主要是由于天气因素造成的,2011年9月底出现连续降雨的天气,再加上所处位置受到海风的影响,导致种子脱落。从收集到的种子成熟程度来说,这时期的种子多为非完好种子,而11月下旬到12月中旬期间的种子多为完好种子,这时期种子雨高峰期则是由于朴树种子成熟而自然脱落,此时,也同样有受到外界自然力的作用。说明了朴树种子雨受两个因素的影响,一是朴树生物学特性,另一个是环境因子(如风向、风速、雨量)。已有研究也证实:种子雨量不仅与树木生物学特性有关,环境因子(如地形、风向、风速、气温、湿度、雨量等)也影响种子雨的分布[18-19];本研究仅对朴树种子雨量时空分布进行了研究,但是对于风向、风速、雨量等环境因子对种子雨的影响没有开展,为了更进一步了解朴树种子雨受外界环境因子的影响,还需在这方面开展更深入的研究。
在朴树防护林内,由于外力(海风、海鸟)的作用,种子雨在空间上表现出明显的异质,林下的种子雨总量、完好种子和非完好种子与林隙内相比均达到极显著差异;朴树种群更新苗在林隙内主要采取实生更新,而在林下则主要采用萌生更新[16];影响朴树在林下与林隙内不同更新方式的原因不是种源的多少,而是林隙与林下微生境的差异,即林隙内的微生境更适合朴树种子萌发、生长,而林下的微生境不利于其萌发、生长。但是引起林隙与林下微生境差异的因素有很多(如:光照强度、温湿度、土壤理化性质等)[8,20-21]。对林隙与林下的环境因子观测可知,在夏季,林隙内的光照强度、温度、土壤温度均高于林下,湿度低于林下;在冬季,林隙内的光照强度、温度高于林下,林隙内湿度与林下相当,土壤温度在12:00—16:00高于林下。在实验室培养箱中,对朴树种子进行萌发实验,朴树种子基本上不萌发;而从研究地取土样(包括凋落物层及以下土壤表土层0~5 cm)带回进行室外观测,结果在林隙内的土样有朴树苗长出。由此推断,凋落物在朴树种群天然更新过程中起到了重要作用。凋落物通过改变微生境中的土壤表面温度、湿度,影响资源的有效性(光、水、养分),并影响生物间的作用,从而直接或间接地影响植物更新[22-24]。那么,在朴树林中,凋落物时空分布有何特征,凋落物分解释放的物质对朴树种子萌发产生何效应?是不是林隙内凋落物促进朴树种子萌发,而林下的凋落物抑制朴树种子萌发?还是林隙内凋落物与环境因子共同作用促进朴树种子萌发?这将是今后研究的方向。
[1] Willson M F, Traveset A. The ecology of seed dispersal[C]Fenner M ed. Seeds: The Ecology of Regeneration in Plant Communities. CAB International, Wallingford,2000:85-110.
[2] 尹华军,程新颖,赖挺,等.川西亚高山65年人工云杉林种子雨、种子库和幼苗定居研究[J].植物生态学报, 2011,35(1):35-44.
[3] Moles A T, Drake D R. Potential contribution of the seed rain and seed bank to regeneration of native forest under plantation pine in New Zealand[J]. New Zealand Journal of Botany,1999,37(1):83-93.
[4] Peng J, Li X G, Fu YC, et al. Seed rain and seed bank of constructive species in evergreen broadleaved forest at Chongqing Simian Mountain[J]. Chinese Journal of Applied Ecology,2000,11(1):22-24.
[5] Nathan R, Muller-Landau H C. Spatial patterns of seed dispersal, their determinants and consequences for recruitment[J]. Trends in Ecology & Evolution,2000,15(7):278-285.
[6] Malgorzata J B, Peter J G. Changing perspectives on the role of the soil seed bank in northern temperate deciduous forests and in tropical lowland rain forests: parallels and contrasts[J]. Perspectives in Plant Ecology, Evolution and Systematics,2006,8(1):3-21.
[7] JuanMD,RobinLC.Interactingeffectsofcanopygap,understoryvegetationandleaflitterontreeseedlingrecruitmentandcompositionintropicalsecondaryforests[J].ForestEcologyandManagement,2008,255(11):3716-3725.[8] 宋新章,肖文发.林隙微生境及更新研究进展[J].林业科学,2006,42(5):114-119.
[9] 龙翠玲,余世孝.茂兰喀期特森林林隙种子雨、种子库空间变异[J].云南植物研究,2007(3):327-332.
[10] 吴敏,张文辉,周建云,等.秦岭北坡不同生境栓皮栎种子雨和土壤种子库动态[J].应用生态学报,2011,22(11):2807-2814.
[11] 秦一芳.朴树种子生理生化及萌发特性研究[D].福州:福建农林大学,2013.
[12] 林承超,魏守珍,肖海燕.从琅岐岛滨海沙丘朴树群落特征探讨福建沿海防护林更新与改造问题[J].生态学报,1995,15(1):54-60.
[13] 蔡小英,范海兰,洪滔,等.福州琅岐风景区景观林朴树种群生命表分析[J].北华大学学报:自然科学版,2006,7(6):545-548.
[14] 闫淑君,洪伟,吴承祯.海风干扰形成朴树残桩萌枝更新的初步研究[J].热带作物学报,2010,31(11):1921-1927.
[15] 闫淑君,洪伟,吴承祯.闽江口琅岐岛朴树种群结构与空间分布格局[J].生态学杂志,2011,30(2):221-226.
[16] 闫淑君,洪伟,林勇明,等.闽江口琅岐岛风景区朴树种群天然更新特征[J].林业科学,2013,49(4):147-151.
[17] 张希彪,王瑞娟,上官周平.黄土高原子午岭油松林的种子雨和土壤种子库动态[J].生态学报,2009,29(4):1877-1884.
[18] 杜彦君,马克平.浙江古田山自然保护区常绿阔叶林种子雨的时空变异[J].植物生态学报,2012,36(8):717-728.
[19] 郭俊杰,赵志刚,欧景莉,等.广西靖西西南桦天然林种子雨的时空动态[J].植物生态学报,2012,36(8):729-738.
[20] Oliveira M V N, Ribas L A. Forest regeneration in artificial gaps twelve years after canopy opening in Acre State Western Amazon[J]. Forest Ecology and Management,2011,261(11):1722-1731.
[21] Hart J L, Buchanan M L, Clark S L, et al. Canopy accession strategies and climate-growth relationships in Acer rubrum[J]. Forest Ecology and Management,2012,282(15):124-132.
[22] Cavieres L A, Chacon P, Penaloza A, et al. Leaf litter ofKageneckiaangustifoliaD. Don (Rosaceae) inhibits seed germination in sclerophyllous montane woodlands of central Chile[J]. Plant Ecology,2007,190(1):13-22.
[23] Dupuy J M, Chazdon R L. Interacting effects of canopy gap, understory vegetation and leaf litter on tree seedling recruitment and composition in tropical secondary forests[J]. Forest Ecology and Management,2008,255(11):3716-3725.
[24] Bray S R, Kitajima K, Mack M C. Temporal dynamics of microbial communities on decomposing leaf litter of 10 plant species in relation to decomposition rate[J]. Soil Biology and Biochemistry,2012,49:30-37.
Temporal and Spatial Distribution of Seed Rain ofCeltissinensisCommunity Shelter Forest in Langqi Island of Minjiang Estuary, Fujian Province
Yan Shujun, Chen Ying, Chen Ying, Fan Hailan, Lin Yongming, Cai Yuanyuan, Chen Mengyi(Fujian Agricultural and Forestry University, Fuzhou 350002, P. R. China); Lei Shaofei(Xi’an Jiaotong University)//Journal of Northeast Forestry University,2015,43(1):13-16,31.
Seed rain; Gap; Temporal and spatial distribution;Celtissinensis; Minjiang Estuary
1) 国家自然科学基金项目(31100303);福建省科技重大专项项目(2012NZ0001)。
闫淑君,女,1975年6月生,福建农林大学园林学院,副教授。E-mail:ysjch2000@gmail.com。
2014年4月17日。
Q948.1
责任编辑:王广建。
