哀牢山滇南山杨幼苗生境选择特征1)
2015-03-10巩合德彭一航燕腾郑丽
巩合德 彭一航 燕腾 郑丽
(西南林业大学,昆明,650224) (云南省科学技术情报研究院)
哀牢山滇南山杨幼苗生境选择特征1)
巩合德 彭一航 燕腾 郑丽
(西南林业大学,昆明,650224) (云南省科学技术情报研究院)
为探究哀牢山滇南山杨(Populusrotundifolia)幼苗的生境选择特征,对其幼苗进行了移栽试验,移栽后的幼苗置于人工遮阴生境中,对其相关的生长参数和气体交换参数进行了测定。结果表明:滇南山杨幼苗生长参数和气体交换参数均随光照强度增加而增大,而比叶面积则随着光照强度增大而减小。可见,光照强度是影响植物生长的重要环境因子,在一定光照强度范围内,幼苗生长都随光照的增强逐渐加快。在1层和2层遮阴网条件下,与其它水分条件相比,土壤含水量为20%(极端干旱)时,滇南山杨幼苗的生长参数均表现出较高水平,说明不耐阴的树种滇南山杨对水分变化不敏感,而且有较高的耐旱性。
遮阴试验;土壤水分;滇南山杨;幼苗
Seedlings ofPopulusrotundifoliawere transplanted to different environmental conditions for observing their habitat preference. A couple of parameters of seedling early growth and gas exchange were measured in the artificial shading conditions outside of the forest. The parameters of seedling early growth and gas exchange ofPopulusrotundifoliashowed a positive correlation with the increase of light intensity, while their specific leaf area (SLA) decreased. Visible, light intensity was one of the important environmental factor affecting plant growth, and seedling growth was enhanced with the light gradually accelerated in a certain light intensity.P.rotundifoliaseedlings performed better at 20% soil moisture (drought stress) at 28.36% and 8.10% light levels; therefore,P.rotundifoliaseedlings might be more drought-tolerant.
幼苗的早期生长速率强烈影响甚至决定了幼苗能否成功定居[1]。在幼苗发育的初期,其子叶中贮藏的碳水化合物和矿物质较少,不能满足幼苗生长的需要,因此,轻微的环境胁迫往往就可能导致幼苗的死亡[2]。水分是影响幼苗生长的关键因子之一,水分胁迫能够明显地限制幼苗的生长。研究发现,在一定范围内,部分植物的叶片总数随着土壤水分水平的降低而减少,单个叶片的伸展也较小,从而导致叶片面积的减小。幼苗比叶面积随水分胁迫的加强而降低,较小的幼苗比叶面积能够增加单位叶面积对二氧化碳的吸收量,还可以在土壤水分亏缺时提高水分的利用效率[3]。Lambers等[4]认为,植物可能主要是通过降低幼苗的比叶面积来调整光合产物在叶片中的分配,以防御干旱带来的叶片损伤。在荫蔽环境下,如果一种植物比另一种植物具有相对更高的相对生长率,那么它在全光照下则表现为较低的相对生长率,说明植物在不同的光照条件下生长情况可能有很大差异[5]。有的树木幼苗能够形成一种利用林下低光照而使幼苗成功定居和最终发育到成熟阶段的策略[6]。于洋等[7]对热带雨林中的绒毛番龙眼幼苗早期建立的研究表明,不同的光照强度对幼苗生长有不同影响,其幼苗能在弱光环境下长期存活并缓慢生长,但在林窗环境下有较高的相对生长速率。笔者研究了滇南山杨幼苗在不同生境中的生长表现及其生境选择行为,旨在找出幼苗致死原因及更新特征,为当地常绿阔叶林的物种多样性保护和森林管理提供科学依据。
1 研究区概况
研究区域位于云南省中部的哀牢山国家级自然保护区的徐家坝地区,海拔2 400~2 600 m。根据哀牢山森林生态系统研究站的长期监测资料,研究区域的平均年降水量为1 931 mm,旱季、雨季分明,雨季(5—10月份)的降雨量占年降雨量的85%左右。年平均蒸发量为1 485 mm,年平均气温为11.3 ℃,最热月(7月份)气温为16.4 ℃,最冷月(1月份)气温为5.4 ℃,无霜期为200 d左右。土壤的成土母质由片岩、片麻岩和闪长岩组成,多发育为典型的黄棕壤,土壤肥沃、偏酸性(pH=4.4~4.9),土壤表面由厚度为3~7 cm的凋落物层覆盖,土壤有机碳、全氮和全磷质量分数分别为2.91%、0.52%和0.06%[8]。哀牢山的中山湿性常绿阔叶林保护完好,主要由壳斗科、茶科、樟科和木兰科的树种组成。其中,壳斗科的硬壳柯(Lithocarpushancei)、木果柯(Lithocarpusxylocarpus)、变色锥(Castanopsisrufescens),茶科的南洋木荷(Schimanoronhae)、翅柄紫茎(Stewartiapteropetiolata)、云南连蕊茶(Camelliaforrestii),樟科的黄心树(Machilusgamblei)、黄丹木姜子(Litseaelongata),木兰科的红花木莲(Manglietiainsignis)、多花含笑(Micheliafloribunda)是乔木层的主要优势种。此外,林中藤本植物和附生植物也相当丰富和发达,层间植物以木质大藤本为常见,附生植物以苔藓和蕨类植物为主,形成奇特的森林景观[9]。
2 材料与方法
选择阳性树种滇南山杨(Populusrotundifolia)正常生长的幼苗(幼苗形态、生长状况基本相同),对其幼苗开展移栽试验。移栽之前,先在原地测定其光合作用特征,主要测定最大净光合速率、暗呼吸速率及光饱和点和光补偿点,然后将幼苗连带其土壤移栽到实验盆中。
在林外建立不同梯度的遮阴试验,采用LI-6400(Li-Cor,Lincoln,Nebraska,USA)记录3种光处理荫棚内的光量子通量密度,从08:00—19:00,每隔1 h测定一次,连续测定3 d(正常天气)。分别用1层(光量子通量密度为林外裸地的28.36%)、2层(光量子通量密度为林外裸地的8.10%)和3层(光量子通量密度为林外裸地的2.37%)黑色尼龙遮阴网,分别模拟林窗中心、林窗边缘和林下的光照强度,每个光处理设3个重复,共计9个荫棚(规格为8 m×8 m×3 m)。荫棚四周封闭至距地面20 cm处,保持地表空气流通,使荫棚内外地表温度一致。在荫棚内用透明塑料遮雨,每一光水平下平均分成4个小区,设置4个土壤水分条件:20%、28%、38%、48%(体积含水量),分别代表极端干旱条件、2005—2006年最低土壤含水量、中度土壤含水量和最高土壤含水量的林下土壤水分条件(参阅哀牢山生态站的长期监测数据)。每天用土壤水分测量仪(MPM-160)测量每组的3盆幼苗,然后对每组的每盆幼苗补充相同的水分,将土壤水分含量控制在相对应的范围内(±2%)。为操作方便和防止小区间的土壤水分交换,相邻两小区间留出100 cm的通道。在每个小区放置10盆幼苗,共有幼苗10×3个重复×4个水分处理×3个光照强度=360株幼苗。试验从2009年6月21日开始,至2010年8月25日结束。
测定滇南山杨幼苗不同光照梯度和不同水分梯度下的生长和生理特征。每个月记录幼苗株高、基径、叶片(复叶)数、单株叶面积,其中单株叶面积的计算方法为:随机采3个幼苗叶片回室内,用扫描仪获得待测叶片的平面图像,统计图像中叶片所占的像素数,乘以每个像素所占的实际面积就可以计算出叶片的面积。1 a后将幼苗分根、茎(含分枝)和叶片在烘箱中85 ℃烘干48 h后分别称质量。根据以上数据计算出幼苗的总生物量、根冠比(RSR)、幼苗比叶面积(SLA)、质量相对生长速率(RGRM)。计算公式如下:
RSR=根干质量/茎叶干质量。
SLA=叶面积/叶片干质量。
RGRM=(lnW2-lnW1)/(T2-T1)。
式中:W1为时间T1时的幼苗干质量;W2为时间T2时的幼苗干质量。
在幼苗生长6个月后,对荫棚内的一半幼苗进行光合作用特征测定,进而推算所有幼苗的情况。所有测定均于09:00—12:00进行,每个月测定1次,共测定6个月,数据用光合助手分析和处理。
3 结果与分析
3.1 幼苗在不同光环境下对土壤水分梯度的响应
3.1.1 不同层遮阴网下幼苗对土壤水分梯度的响应
由表1可见,1层遮阴网条件下,滇南山杨幼苗的基径增量、叶片增量、根冠比、质量相对生长速率和总生物量在土壤含水量为48%的水分条件下最大,而株高增量在土壤含水量为28%时最大,并且显著高于土壤含水量38%时。幼苗存活率在土壤含水量为28%和38%时最高,比叶面积在各土壤含水量条件下无显著差异(p>0.05)。2层遮阴网条件下,滇南山杨幼苗的高度增量在土壤含水量为38%时最大,其叶片增量、质量相对生长速率和总生物量在土壤含水量为48%时最大。在土壤含水量为20%时,其基径增量和根冠比最大,比叶面积最小,而且土壤含水量为20%时的基径增量显著高于48%的结果,土壤含水量为38%时的高度增量显著高于28%时的结果。3层遮阴网条件下,滇南山杨幼苗的存活率普遍较低,其生长参数变化也无明显的规律。基径增量、质量相对生长速率和总生物量都在土壤含水量为20%时最大,株高增量和根冠比在土壤含水量为28%时最大,叶片增量在土壤含水量为38%时最大,土壤含水量为48%时,其比叶面积最大,但基径增量、高度增量和叶片增量却最小。质量相对生长速率和总生物量在土壤含水量为28%时最小,而且土壤含水量在28%时的比叶面积显著低于48%时的结果。
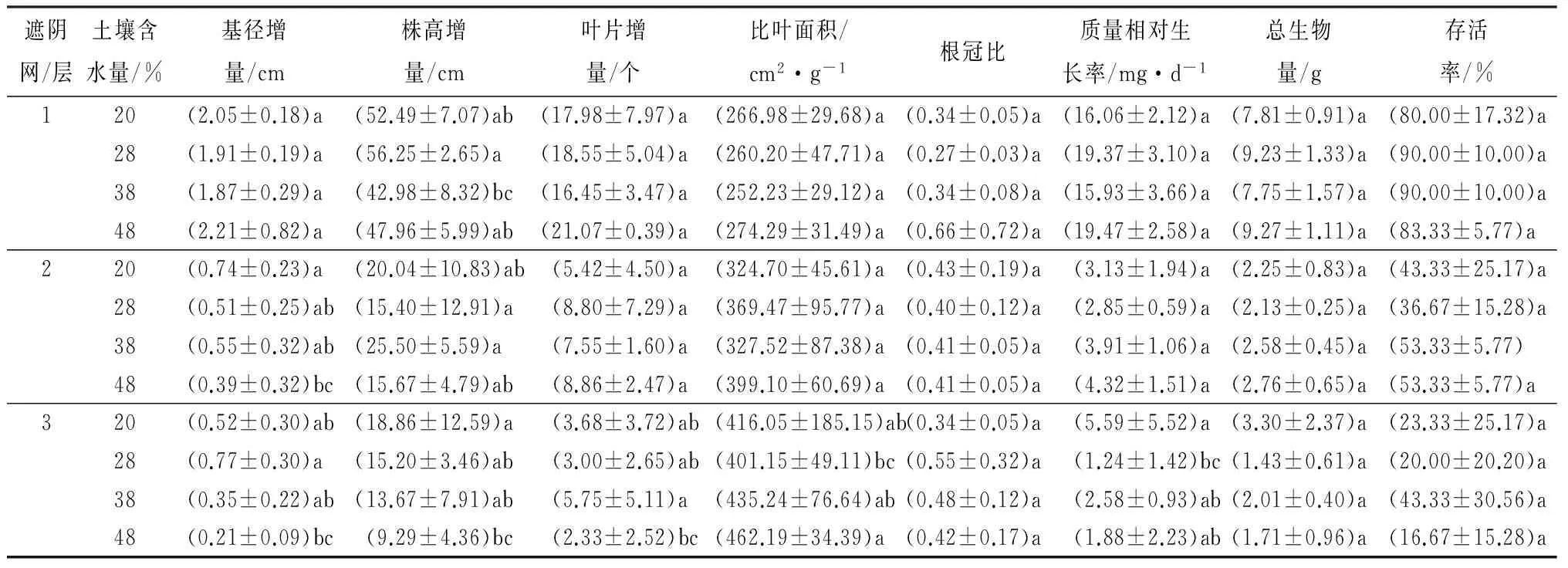
表1 不同光照强度和不同水分条件对滇南山杨幼苗生长的影响
注:表中数据为平均值±标准差。同列不同字母表示在不同生境间差异显著(p<0.05)。
3.1.2 幼苗对极端干旱条件(土壤含水量20%)的响应
与其它水分条件相比,土壤含水量为20%时,在1层遮阴网遮阴处理下,滇南山杨的幼苗除了存活率最低外,其它生长参数均表现为较高水平;2层遮阴网处理下,滇南山杨幼苗的叶片增量和比叶面积最低;3层遮阴网处理下,滇南山杨幼苗的根冠比最小(表1)。
3.2 幼苗气体交换参数在不同光环境下对土壤水分梯度的响应
3.2.1 不同遮阴处理下幼苗气体交换参数对土壤水分梯度的响应
由表2可见,1层遮阴网条件下,滇南山杨幼苗的暗呼吸速率、光补偿点和光饱和点都在土壤含水量为48%时最大,土壤含水量为38%时最大净光合速率最大,最大净光合速率和光饱和点都在土壤含水量为28%时最小。2层遮阴网条件下,滇南山杨幼苗在土壤含水量为48%时暗呼吸速率和光补偿点最大,土壤含水量为38%时最大净光合速率和光饱和点最大,而且土壤含水量为48%时的光补偿点显著高于土壤含水量为20%、28%和38%时的结果。3层遮阴网条件下,滇南山杨幼苗的暗呼吸速率在土壤含水量为20%时最大,最大净光合速率和光饱和点在土壤含水量为38%时最大,而且其最大净光合速率在土壤含水量为20%、28%和38%时显著高于48%时的结果,其光补偿点在土壤含水量为48%时最大。
3.2.2 幼苗气体交换参数对极端干旱条件(土壤含水量20%)的响应
与其它土壤含水量相比,土壤含水量20%时,在1层遮阴网处理下,滇南山杨幼苗的暗呼吸速率、最大净光合速率和光补偿点均最低;在2层遮阴网处理下,滇南山杨幼苗的暗呼吸速率和最大净光合速率最低;在3层遮阴网处理下,滇南山杨幼苗的光补偿点和光饱和点均最低,但暗呼吸速率最大。
表2 光照强度和水分条件对滇南山杨幼苗气体交换参数的影响

遮阴网/层土壤含水量/%暗呼吸速率/μmol·m-2·s-1最大净光合速率/μmol·m-2·s-1光补偿点/μmol·m-2·s-1光饱和点/μmol·m-2·s-1120(0.55±0.45)a(13.20±2.00)a(8.79±7.07)a(238.67±26.73)a28(0.56±0.37)a(11.80±4.04)a(8.77±4.78)a(198.00±58.64)a38(0.89±0.26)a(14.30±3.35)a(14.47±4.62)a(243.67±57.07)a48(1.65±0.64)a(13.97±0.51)a(28.33±12.95)a(263.00±25.36)a220(0.19±0.03)bc(5.61±0.67)a(4.00±1.15)b(122.67±16.65)a28(0.24±0.09)ab(6.24±1.15)a(3.83±0.96)b(104.43±10.92)a38(0.31±0.09)ab(7.90±0.66)a(4.52±0.69)b(123.00±7.00)a48(0.53±0.16)a(6.71±0.21)a(8.61±1.78)a(122.07±23.19)a320(0.32±0.03)a(6.97±0.27)a(4.67±0.61)a(108.33±9.71)a28(0.30±0.07)a(7.05±1.13)a(4.69±1.09)a(113.00±16.09)a38(0.30±0.26)a(7.43±1.03)a(5.01±4.16)a(136.67±8.96)a48(0.22±0.03)a(4.51±0.22)b(6.04±1.57)a(127.67±14.36)a
注:表中数据为平均值±标准差。不同字母表示在在相同光照条件下不同土壤含水量间差异显著(p<0.05)。
4 讨论
4.1 人工遮阴不同光照梯度下幼苗存活及生长
在人工遮阴不同的光照梯度下,滇南山杨幼苗的基径增量、株高增量、叶片增量、质量相对生长速率、总生物量和最大净光合速率均随光照强度增加而增大,而比叶面积随光照强度增大而减小,与闫兴富等研究结果[10]一致。但本项研究的最大光强仅为林外裸地的28.36%,并不能说明光强越大,幼苗生长越好,可能存在一个适度的光照强度范围,在此范围内幼苗生长情况最好,低于或超过这个范围可能都不利于幼苗的生长[11-13]。研究结果表明,光是影响植物生长的重要环境因子,在一定光照强度范围内,幼苗生长都随光照的增强逐渐加快。滇南山杨的根冠比在较弱的光强(裸地光强的2.37%)下最大,不同于闫兴富等[10]的研究结果,这可能是本研究结果来自于土壤水分充足的条件(土壤含水量38%时),也说明光照和土壤含水量均是影响树种幼苗根冠比的重要因素[14]。
滇南山杨幼苗的各项气体交换参数均在1层遮阴网处理下最大,但随光照强度的降低,气体交换参数结果无规律,而方江保等[15]对苦槠(Castanopsissclerophylla)幼苗和姚志刚等[16]对银缕梅(Parrotiasubaequalis)幼苗的研究发现,随光强的降低,最大净光合速率、光补偿点和光饱和点均降低,这可能是树种不同造成的。从另外一方面说明植物并不是消极被动地适应环境,它们可以通过自身形态、生理等方面的不断调整来适应环境中生态因子的变化,将其限制作用减小。因此在不同的环境中,植物会产生不同的适应性变异[17-18]。
4.2 幼苗在不同光环境下对土壤水分梯度的响应
不同的遮阴条件下,滇南山杨幼苗的生长参数和气体交换参数变化无规律,有的参数甚至在土壤体积含水量20%或28%水分条件下最大,表明在适度缺水环境下,滇南山杨有一定的适应性和抵抗效应,适度缺水反而更有利于其生长,也从另外一方面说明,滇南山杨幼苗对水分变化并不敏感,这对滇南山杨幼苗在强光及干旱环境中生长和生存具有重要意义。
4.3 幼苗对极端干旱条件(土壤含水量20%)的响应差异
在1层和2层遮阴网条件下,与其它水分条件相比,土壤含水量为20%时,滇南山杨幼苗的生长参数均表现出较高水平,说明不耐阴的树种滇南山杨对水分变化不敏感,而且有较高的耐旱性。李晓清等[19]的研究认为,干旱胁迫时,植物幼苗更倾向于将资源分配给根系,但限制叶片生长,使光合总面积减小,光合总产物也减少,从而提高了根冠比。但滇南山杨幼苗的根冠比在干旱条件时并不是最大,说明在干旱条件下,幼苗对水分胁迫可能是长期的适应过程,其形态结构和生理生态功能也是对干旱长期适应的结果,换句话说,由于该研究区域降雨充足,温度较低,蒸发较小,滇南山杨幼苗受干旱胁迫的程度相对较小,所以其幼苗在一定范围内对土壤水分的变化均不敏感。也表明干旱胁迫下,植物幼苗生物量分配特征在不同树种之间存在着较大差异,并不能一概而论[20]。在3层遮阴网条件下,滇南山杨幼苗的生长参数大多不是最低值,也说明在较低光照的条件下,土壤水分条件对幼苗生长的影响不大。不同光照条件下,与其它水分条件相比,土壤含水量为20%时,滇南山杨幼苗的气体交换参数无明显的规律,说明干旱条件下,水分亏缺可能降低了一些气体交换参数,但并不明显。
5 结论
在人工遮阴不同的光照梯度下,滇南山杨幼苗的基径增量、株高增量、叶片增量、质量相对生长速率、总生物量、最大净光合速率和各项气体交换参数均随光照强度增加而增大,而比叶面积随光照强度增大而减小,可见,光照强度是影响植物生长的重要环境因子,在一定光照强度范围内,幼苗生长都随光照的增强逐渐加快。在1层和2层遮阴网条件下,与其它水分条件相比,土壤含水量为20%时,滇南山杨幼苗的生长参数均表现出较高水平,说明不耐阴的树种滇南山杨对水分变化不敏感,而且有较高的耐旱性。
[1] Huston M A, Smith T M. Plant succession: life history and competition[J].The American Naturalist,1987,130(2):168-198.
[2] Kozlowski T T. Physiological ecology of natural regeneration of harvested and disturbed forest stands: implications for forest management[J]. Forest Ecology and Management,2002,158(1/3):195-221.
[3] Burslem D F R P, Grubb P J, Turner I M. Responses to simulated drought and elevated nutrient supply among shade-tolerant tree seedlings of lowland tropical forest in Singapore[J]. Biotropica,1996,28(4b):636-648.
[4] Lambers H, Poorter H. Inherent variation in growth rate between higher plants: a search for physiological causes and ecological consequences[J]. Advances in Ecological Research,1992,23:187-261.
[5] Thomas S C, Bazzaz F A. Asymptotic height as a predictor of photosynthetic characteristics in Malaysian rain forest trees[J]. Ecology,1999,80(5):1607-1622.
[6] Osunkoya O O, Ash J E, Hopkins M S, et al. Influence of seed size and seedling ecological attributes on shade-tolerance of rain-forest tree species in northern Queensland[J]. Journal of Ecology,1994,82(1):149-163.
[7] 于洋,曹敏,郑丽,等.光对热带雨林冠层树种绒毛番龙眼种子萌发及其幼苗早期建立的影响[J].植物生态学报,2007,31(6):1028-1036.
[8] Liu Wenyao, Fox J E, Xu Zaifu. Biomass and nutrient accumulation in montane evergreen broad-leaved forest (Lithocarpus xylocarpus type) in Ailao Mountains, SW China[J]. Forest Ecology and Management,2002,158(1/3):223-235.
[9] 邱学忠.哀牢山森林生态系统研究[M].昆明:云南科技出版社,1998:12-27.
[10] 闫兴富,曹敏.不同光照梯度的遮荫处理对绒毛番龙眼幼苗生长的影响[J].热带亚热带植物学报,2007,15(6):465-472.
[11] 张光飞,欧阳志勤,苏文华.生境光强对云南金钱槭幼苗生长及光合速率的影响[J].湖北农业科学,2009,48(11):2780-2782.
[12] 金鑫,胡万良,丁磊,等.遮阴对红松幼苗生长及光合特性的影响[J].东北林业大学学报,2009,37(9):12-13,47.
[13] 阴卓越.4种乔木幼苗对遮荫与芒萁水提液的响应机制[D].杭州:浙江农林大学,2013.
[14] 闫兴富,周立彪,李静,等.遮阴对紫荆种子萌发和幼苗生长的影响[J].江苏农业科学,2010(4):176-179.
[15] 方江保,殷秀敏,余树全,等.光照强度对苦槠幼苗生长与光合作用的影响[J].浙江林学院学报,2010,27(4):538-544.
[16] 姚志刚,王中生,颜超,等.濒危植物银缕梅幼苗对不同光强的光合响应[J].南京林业大学学报:自然科学版,2010,34(3):83-88.
[17] 刘志发,申卫军,温达志.南澳岛光裸地和次生林下6种乡土树幼苗生长与光合特征[J].生态环境学报,2009,18(2):621-628.
[18] 李佳杏,黄小辉,刘芸,等.模拟三峡库区消落带土壤水分条件下的桑树幼苗生长状况[J].蚕业科学,2012,38(2):210-215.
[19] 李晓清,胡学煜,左英强,等.水分胁迫对希蒙得木幼苗生物量的影响[J].西南林学院学报,2009,29(5):19-22,27.
[20] 李芳兰,包维楷,吴宁.白刺花幼苗对不同强度干旱胁迫的形态与生理响应[J].生态学报,2009,29(10):5406-5416.
Habitat Preference Characteristics ofPopulusrotundifoliaSeedlings in the Ailao Mountains
Gong Hede, Peng Yihang, Yan Teng(Southwest Forestry University, Kunming 650224, P. R. China); Zheng Li(Yunnan Academy of Scientific & Technical Information)//Journal of Northeast Forestry University,2015,43(1):1-4.
Shading experiment; Soil moisture; Seedlings ofPopulusrotundifolia
1) 国家自然科学基金项目(31200482)。
巩合德,男,1978年8月生,西南林业大学生态旅游学院,副教授。E-mail:gonghede3@163.com。
2014年4月28日。
S718.51
责任编辑:程 红。
