东海营养盐结构的时空分布及其对浮游植物的限制*
2015-03-08刘艳云崔永平王泽寿叶仙森
叶 然 刘艳云 崔永平 王泽寿 叶仙森
(国家海洋局宁波海洋环境监测中心站 宁波 315012)
东海是我国陆架最宽的边缘海, 海域面积宽阔,与太平洋及南海水交换方便, 环流发育相当充分。受到以长江、钱塘江、瓯江、闽江四大水系为代表的主要陆源径流入海物质通量的强烈影响及多种来源不同水团交汇的共同作用, 东海营养盐循环特征复杂多变, 含量较为丰富, 利于浮游植物的生长和繁殖,是我国海洋生产力最高的海域(洪华生, 2012)。近年来, 随着沿海地区海洋经济的高速发展, 人类社会活动的高密度投入导致氮(N)、磷(P)等特征污染物大量入海, 水体富营养化程度严重, 赤潮等海洋灾害频发,海洋生态环境面临巨大压力(周名江等, 2003)。
浮游植物是海洋生态系统中的初级生产者, 而营养盐作为其赖以生存的物质基础, 在海水中的输入、扩散的物理和生化过程对浮游植物的群落演替起着重要作用(王江涛等, 2012), 且不同环境海域的营养盐含量和结构比例会对浮游植物的生长发育造成一定差异(李京, 2008)。有研究指出, 浮游植物并非完全按照一定比例吸收海水中的营养物质, 其生长将受到含量相对较少的营养盐的限制(Hecky et al,1988)。一般来说, 河口沿岸海域多以P限制为主, 而开阔海域及大洋水多发生N限制(Ryther et al, 1971)。
关于东海营养盐的分布特征及其对浮游植物生长限制因子和受限机理, 已有许多国内学者在长江口及其邻近海域(王修林等, 2004; 赵卫红等, 2006;米铁柱等, 2012)以及受黑潮影响的远海区域(王芳等,2008; 张建平等, 2010; 刘超等, 2013)作了相关研究,对于东海大范围内的研究也有报道(王保栋, 2003;Wang et al, 2003), 但多数缺乏与过往历史数据的比较, 难以看出变化趋势。本文基于2013年春、夏、秋、冬四个航次的调查数据, 分析东海海域不同季节营养盐结构的时空分布, 讨论不同季节浮游植物生长的营养盐限制情况, 同时结合历史资料, 将本文研究的结果与近10年东海区夏季营养盐的限制区域进行比较, 为进一步研究东海海域营养盐结构的时空变化对东海生态环境的影响提供基础资料。
1 材料与方法
1.1 调查时间与站位布设
国家海洋局宁波海洋环境监测中心站于2013年2 月(春季)、5 月(夏季)、8 月(秋季)、11 月(冬季)分别对东海海域(120°—128°E、25°—33°N)进行了 4 个航次的现场综合调查。每个航次调查均布设3条断面和15个辅助站, 共计41个站位。调查站位见图1。
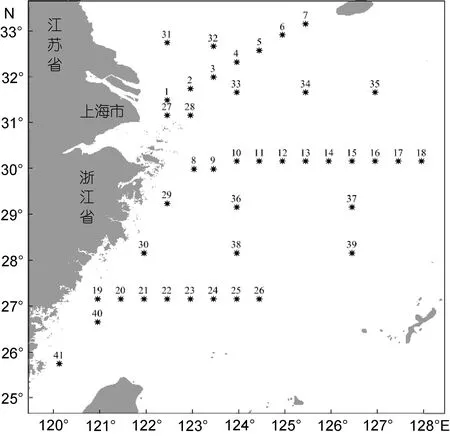
图1 现场调查站位图Fig.1 Sampling sites in the East China Sea
1.2 样品采集与分析
采用CTD (美国Seabird)获取现场温度、盐度及深度数据, CTD自容式采样器采取水样。营养盐、溶解氧均在现场测定, 无机氮(DIN)为硝酸盐(NO3–-N)、亚硝酸盐(NO2–-N)及铵盐(NH4+-N)的浓度之和, 分别用锌-镉还原分光光度法、萘乙二胺分光光度法和次溴酸钠氧化分光光度发测定, 磷钼蓝分光光度法测定磷酸盐(PO43–-P), 硅钼黄分光光度发测定硅酸盐(SiO32–-Si)。叶绿素 a样品经量取 300mL水样, 用孔径为 0.45μm的纤维素酯微孔滤膜过滤, 用滤纸包好并干燥, 于冷冻(-20°C)环境下保存, 带回陆基实验室, 用荧光分光光度法测定。
2 结果与讨论
2.1 营养盐含量分布
研究海域四季表层营养盐含量、变化范围及平面分布如表1、图2所示。由此可见, 三种营养盐浓度大体呈现自西向东递减的变化趋势, 春、夏季DIN、含量的高值区均集中在 123°E以西的长江口海域, 此处等值线分布密集, 形成营养盐锋面, 自此向外开始迅速递减, DIN和PO43–-P在东海中部混合水区域几乎被消耗殆尽; 而秋、冬季则基本南迁至123°E以西、28°N附近的浙南沿岸海域,和还在 124°—125°E、30°—31°N 附近的东海北部区域存在部分高浓度现象。显然, 秋、冬季黄海沿岸流的南下入侵是造成上述分布格局的主要原因(王正方等, 1983; 王保栋, 2003)。总体看来, 秋、冬季营养盐的含量要高于春、夏两季, 且高值区(特别是的东扩现象更为明显。
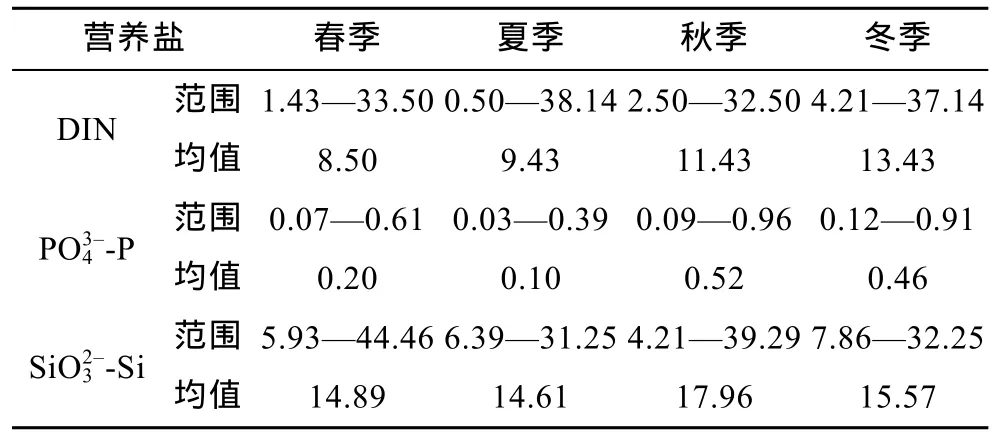
表1 东海四季表层营养盐含量(μmol/L)Tab.1 Contents of nutrients at surface water in 4 seasons overthe East China Sea
2.2 营养盐结构分布
表2和图3分别显示了研究海域表层营养盐四个航次中DIN/P、Si/DIN、Si/P比值的变化范围、均值及平面分布, 图4为表层叶绿素a含量的平面分布。

春季航次DIN/P比值呈现长江口门处较高, 其它海域相对较低的分布格局。在 123°E以西、31°N以北的长江入海口门处形成一个以 1号站为中心的舌状高值区, 最高至44.7。长江口门处是赤潮高发地区,初级生产力水平较高(周伟华等, 2003), 本航次调查中的1号站DIN浓度高达33.50μmol/L, PO43–-P含量仅为0.34μmol/L, 与表层叶绿素a的峰值分布有较好的对应。这表明了以高DIN/P比值为特征的长江冲淡水等陆地径流向东海近岸海区的输入, 同时由于近岸海域相对受到沿岸流的限制作用使得入海营养盐难以向外扩散, 易于积累(Liu et al, 2003), 因而在长江口门处形成了高DIN/P比值分布区。相比受到光照条件限制的长江口内部的浑浊水域(赵卫红等, 2006),在长江口羽状锋区, 海水透明度开始增大, 加之春季适宜的水温使浮游植物大量繁殖并摄取营养盐(朱建荣, 2004), PO43–-P含量被消耗殆尽, 其生长活动显然受到PO43–-P的限制(Harrison et al, 1990)。其他区域DIN/P比值基本低于 35, 部分海域还出现低于Redfield的16∶1比值现象。Si/DIN在研究海域的比值均大于 1, 在济州岛以南的东海东北部海域有一个涡状分布的高值区, 中心值可达19.8, 而叶绿素a均值浓度仅在 1.5μg/L左右, 由此表明在寡营养盐的黑潮表层水影响下, 外海浮游植物的生长可能相对受限于DIN, 同时也体现了以SiO32–-Si为主的九州岛入海径流对东海东北部高Si/DIN比值的贡献(张建平等,2010)。Si/P比值介于28.1—373之间, 其高值区集中在浙南近岸水域, 与高生泉等(2004)的研究结果相一致。对比长江口海域, 该区域的PO43–-P被浮游植物大量消耗, 含量更低(0.07—0.21μmol/L), 增高了Si/P比值。总体看来, 春季航次DIN/P、Si/DIN及Si/P比值的平面分布均表现为高值区域较为集中、向外扩展影响较小的特征。

图2 研究海域四季表层营养盐含量的平面分布(μmol/L)Fig.2 Horizontal distributions of nutrients in the surface water in four season of the study area
夏季DIN/P比值达到全年最高, 平均43.6。与春季相似, DIN/P=16的等值线出现在125°E附近区域。有两个高于均值的高值区, 第1个高值区位于研究海域的东北方向(124.5°E, 32°N), 与夏季长江冲淡水入海后的主体流向相同(朱建荣等, 2003), 32号站和1号站分别达到最高值(97.0)和次高值(93.0), 叶绿素a含量也在32号站出现峰值(26.1μg/L)。与春季航次相比,夏季DIN/P比值即使是在济州岛附近海域, 依然维持在30以上的水平(春季航次只有10左右)。王保栋等(2002)的研究指出夏季长江冲淡水中的营养盐向东北最远可扩散至127°E, 33°N附近, 本航次6号和7号站表层DIN浓度依旧维持在高于10.71μmol/L的水平,但PO43–-P已降至0.10μmol/L左右。因此推断东海北部海域夏季 DIN/P比值异常偏高是在长江充沛的径流量影响下, 水体中的 DIN含量过剩且浮游植物的生长对 PO43–-P的过量消耗而引起的(宋秀贤等,2013)。第2个高值区分布在123°E以西、27°N以南的浙江沿岸上升流区(曹欣中, 1986), 随东海陆架抬升的上升流使底层的高营养盐水不断向上涌升, 与北上的浙江沿岸水交汇, 在此形成均值达 15.5μg/L的叶绿素a高值区, 表明浮游植物的生长活动异常旺盛, PO43–-P 含量更是被消耗至 0.03—0.09μmol/L; 但沿岸水带来的陆源农业排污补充了 DIN含量, 由此形成第2个高DIN/P比值区域, Si/P比值分布情况与之相似。Si/DIN的分布与DIN/P相反, 123.5°以西、28°N以北的浙江近岸海域和127°以西、31°N以北的长江口及其邻近海域的 Si/DIN比值多介于 1—5, 部分站位<1(27号站), 说明N 营养元素相对平衡, Si/DIN比值有利于浮游植物的生长; 台湾岛东北部黑潮区海域的含 量 介 于 9.57—13.39μmol/L之间, 同时 DIN 含量降至最低值(均值1.79μmol/L), 导致这一区域的 Si/DIN 比值高达10.1—20.6, 并向冲绳海槽的东南方向扩展。这与刘超等(2012)认为东海黑潮区表层水的 Si/DIN比值在7—8月达到最大值的研究结果相符。
秋季, 研究海域的 DIN/P比值达到全年最低(均值仅9.3), Si/DIN也相对较低(均值4.1), 分别接近于Redfield的16∶1和1∶1的比值。与DIN/P高值区相对应, 在124°E以西、30°N以北海域 DIN的含量依旧很高(1、2、3号站含量均高于21.43μmol/L), 使得这一区域的Si/DIN比值可降至1左右, 但在125°E以东的外部海域则相反, 暗示比其远海区域, 近岸水域浮游植物的生长可能更需要大量的硅元素(孙松,2012)。与春、夏两季相比, 秋季高 DIN/P区域主要集中在 123°附近的舟山群岛外部海域, 其周围海域亦维持在较高水平。而携带高浓度SiO32–-Si的苏北沿岸流受到东北季风的作用南下, 与少部分输送至东海东北部的长江冲淡水混合后向东南方向入侵, 从而形成124—125°E海域的Si/DIN高值区(王保栋等,2002; 王奎等, 2013)。Si/P比值较前两个季节变化幅度较小, 降至 11—60.7, 其高值区的分布分别与本航次的DIN/P和Si/DIN具有相同的成因机制。
冬季DIN/P以30号站(32.3)和26号站(28.5)为中心分别形成浙南近海和台湾岛东北部两个涡状高值中心。这一带的 DIN含量即使在冬季也可维持在14.07—47.64μmol/L 的水平, 但 PO43–-P 含量却只有0.20—0.60μmol/L, 主要是因为长江冲淡水的南下与入侵东海陆架区的高温高盐、低低的黑潮表层营养盐水汇合, 而在西侵过程中黑潮次表层水的涌升作用又为东海陆架区带来了高含量营养盐, 由此形成了高DIN/P区域(陆赛英, 1996; Chen,1996), 对应此处比值介于1.2—2.1的Si/DIN低值区。调查海域的Si/P和Si/DIN比值的分布相似, 中部的东海混合水区域相对较低, 二者在 127—128°E、29—30°N 均有一个相对高值区, 这可能与 SiO32–-Si本身在海水中的高含量以及冬季外海硅藻的生物活动量较低有关。

表2 东海四季DIN/P、Si/DIN及Si/P比值Tab.2 Ratio of DIN/P, Si/DIN and Si/P in surface water in four seasons of the East China Sea
2.3 营养盐对浮游植物限制的季节变化
关于营养盐对浮游植物生长限制的标准, 已有许多国内外学者做过相应研究(胡明辉等, 1989; Justić et al, 1995), 但这些标准或不够全面或只能判断营养盐的潜在限制作用而不能说明限制是否会发生。本文选用 Fisher等(1992)提出的营养盐半饱和常数(Ks)为评价标准, 判断研究海域的浮游植物生长是否受到营养盐的限制, 当Si/DIN>1, DIN/P<10, 且 DIN<2.0μmol/L时, 为N限制; 当Si/P>22, DIN/P>22, 且P<0.20μmol/L 时, 为 P 限制; 当 Si/P<10, Si/DIN<1, 且 Si<2.0μmol/L 时, 为 Si限制。

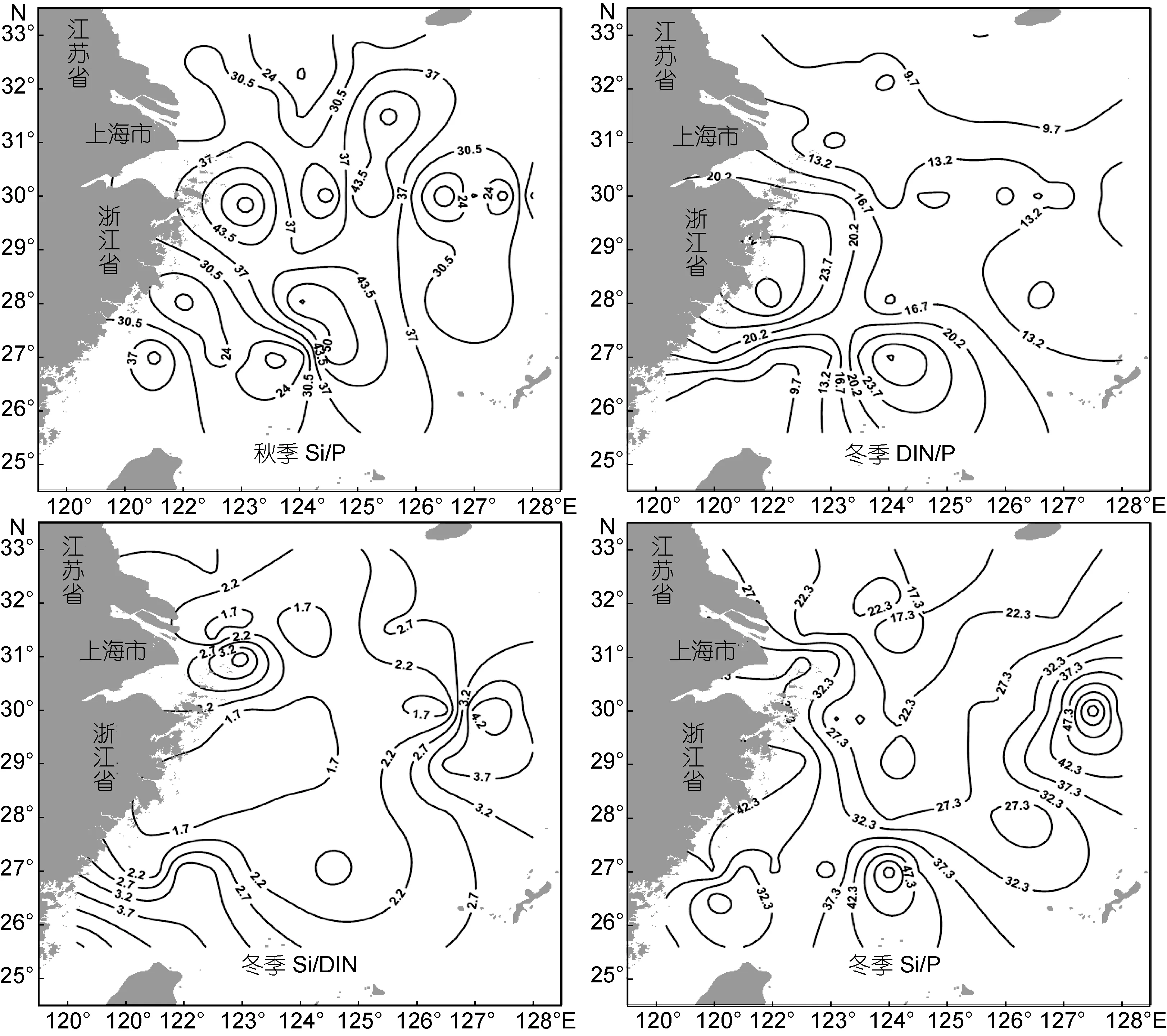
图3 研究海域四季表层营养盐DIN/P、Si/DIN、Si/P比值平面分布Fig.3 Horizontal distributions of DIN/P, Si/DIN and Si/P in surface water in four seasons of the study area

图4 研究海域四季表层叶绿素a含量平面分布(μg/L)Fig.4 Horizontal distributions of chlorophyll-a in surface water in four seasons of the sea area
根据本文的调查结果(图5), 春季航次N限制和P限制分别占总调查站位数的百分比分别为 9.8%和22%; 无Si限制站位。夏季调查海域以 P限制为主,60%的调查站位受到P限制; N限制站位占到19.5%;同样, 本航次无Si限制情况。秋季营养盐限制站位较前两航次大幅度减少, 全航次仅长江口海域的7个站位受到P限制, 无N和Si限制情况, 而冬季由于海水垂直运动强烈, 研究海域水体营养盐平衡状况较好,浮游植物的生长不受营养盐限制的。进一步分析发现受 N限制的站位基本出现在 126°E以东、受黑潮影响强烈的海域。以往的研究表明东海黑潮区全年50m以浅水域的DIN/P比值基本低于10(项有堂等, 1994),而春季正值浮游植物生长繁殖的旺季, 虽然黑潮区高温、高盐的性质使得浮游植物生物量全年差异并不大, 但该区域的DIN含量春、夏季(0.50—1.43μmol/L)明显低于秋、冬两季(2.50—4.21μmol/L), 基本低于本文所采用的浮游植物生长所需 N元素的阈值(2.0μmol/L), 故春、夏季 N限制站位多集中在此。P限制站位大都位于 200m等深线以西的东海陆架区,但夏季受限范围要比春、秋季更大, 除126°E以东的黑潮区站位外, 近岸及中部海域的大部分站位为P限制。夏季台湾暖流和黑潮在该区域势力的强盛, 使低营养盐含量区域扩大(王芳, 2006), 且温跃层的形成阻碍了下层水体营养盐对上层水体的补充, 导致浮游植物生长受P限制的区域面积扩大(谢琳萍, 2012)。Si在全年均无限制情况。由此看来, 研究海域浮游植物的生长主要受到N和P的限制, 126°以西的近岸及大陆架海域以P限制为主, 而126°以东的黑潮区受N限制; 在季节变化上又以夏季受到营养盐的限制最明显, 冬季则无营养盐限制情况。
2.4 与历史资料相比
根据本文的分析, 2013年东海四季营养盐对浮游植物的限制以夏季最为明显, 为达到与历史资料比对的时空一致性, 本文选取 2001—2010年东海海域夏季航次(内部资料, 尚未发表)与 2013年夏季航次的营养盐限制情况作比对分析(表3)。
由表3可知, 2001—2010年近10年间东海海域夏季受到P限制的站位数量较少、范围较小, 均位于123°E以西的长江口及浙北沿岸海域, 除了2004年夏季航次所有站位均未受到营养盐的限制之外,2001—2003年受到 P限制的站位比例逐年小幅度上升, 自2005年起至2010年受P限制的站位数又开始缓慢增加。本次调查的结果显示, 研究海域P限制站位数比过往 10年间有所增加, 且 P限制范围由28°—32°N、123°E 以西的长江口及浙北沿岸海域扩展到了125°E以西的东海中部及浙江近岸水域, 这主要与东海沿岸人类经济活动产生的高 DIN/P比值的农业废水随着陆源径流不断入海密切相关。N限制情况仅在 2007年发生, 与本次调查的结果相同, 受到限制的站位也位于126°—127°E以东的黑潮区, 但本航次的N限制范围要大于往年同期。

表3 东海2001—2010年夏季航次表层营养盐限制站位及范围Tab.3 Patterns of nutrient limitation in the surface water in summer of 2001—2010 in the East China Sea
3 结论
(1)东海 DIN/P、Si/DIN及Si/P比值受各种水团及浮游植物生长周期的影响较为明显, 长江冲淡水与沿岸水的交汇作用控制着全航次DIN/P比值, 基本呈近岸高、远海低的分布规律, 而冬季由于黑潮次表层水的涌升向大陆架的入侵作用, 在东海南部也有个涡状高值中心; 受近岸高DIN/P比值陆源径流入海及浮游植物生长周期的影响, 近岸 Si/DIN高值区基本对应DIN/P低值区, 而受黑潮影响的远海区域则相反; 春、夏季Si/P高值区主要分布在近岸, 而秋、冬季则开始由中部海域向远海扩展。
(2)研究海域浮游植物的生长主要受到N和P的限制, 126°以西的近岸及中部海域以 P限制为主, 而126°以东的黑潮区受 N 限制; 在季节变化上又以夏季受到营养盐的限制最明显, 冬季则无营养盐限制情况。
(3)与2001—2010年同期历史资料相比, 2013年夏季航次受 P限制站位数量比过往 10年有所增加,限制范围由 28°—32°N、123°E以西的长江口及浙北沿岸海域扩展到了 125°E以西的东海中部及浙江近岸水域; 受N限制站位基本集中在126°—127°E以东黑潮区海域, 但受限范围更大。
王 芳, 2006. 东海营养盐分布特征及季节变化研究. 广州:华南师范大学硕士学位论文, 1—21
王 芳, 康建成, 周尚哲等, 2008. 东海外海海域营养盐的时空分布特征. 资源科学, 30(10): 1592—1599
王 奎, 陈建芳, 金海燕等, 2013. 长江口及邻近海区营养盐结构与限制. 海洋学报, 35(3): 128—136
王正方, 姚龙奎, 阮小正, 1983. 长江口营养盐(N, P, Si)分布与变化特征. 海洋与湖沼, 14(4): 324—332
王江涛, 曹 婧, 2012. 长江口海域近 50a来营养盐的变化及其对浮游植物群落演替的影响. 海洋环境科学, 31(3):310—315
王修林, 孙 霞, 韩秀荣等, 2004. 2002年春、夏季东海赤潮高发区营养盐结构及分布特征的比较. 海洋与湖沼, 35(4):323—331
王保栋, 2003. 黄海和东海营养盐分布及其对浮游植物的限制.应用生态学报, 14(7): 1122—1126
王保栋, 战 闰, 藏家业, 2002. 长江口及其邻近海域营养盐的分布特征和输送途径. 海洋学报, 24(1): 53—58
朱建荣, 2004. 长江口外海区叶绿素a浓度分布及其动力成因分析. 中国科学D辑 地球科学, 34(8): 757—762
朱建荣, 丁平兴, 胡敦欣, 2003. 2000年8月长江口外海区冲淡水和羽状锋的观测. 海洋与湖沼, 34(3): 249—254
刘 超, 康建成, 王国栋等, 2012. 东海黑潮区营养盐及其限制作用的月际空间分异. 资源科学, 34(7): 1375—1381
刘 超, 康建成, 王国栋等, 2013. 东海黑潮区硝酸盐含量及其高值区的季节分布特征. 海洋与湖沼, 44(5):1216—1222
米铁柱, 姚庆祯, 孟 佳等, 2012. 2011年春、夏季黄海、东海营养盐分布特征研究. 海洋与湖沼, 43(3): 678—688
孙 松, 2012. 中国区域海洋学—生物海洋学. 北京: 海洋出版社, 204—238
李 京, 2008. 东海赤潮高发区营养盐结构及对浮游植物优势种演替的作用研究. 青岛: 中国海洋大学硕士学位论文,15—32
宋秀贤, 俞志明, 殷克东等, 2013. 香港近岸海域营养盐结构特征及其对浮游植物生长的影响. 海洋与湖沼, 44(4):846—852
张建平, 康建成, 任惠茹等, 2010. 东海北部营养盐分布的季节变化及成因探讨. 海洋科学, 34(1): 35—43
陆赛英, 1996. 东海陆架水域营养盐的季节变化和物理输运的规律. 海洋学报, 18(5): 41—51
周伟华, 霍文毅, 袁翔城等, 2003. 东海赤潮高发区春季叶绿素 a和初级生产力的分布特征. 应用生态学报, 14(7):1055—1059
周名江, 颜 天, 邹景忠, 2003. 长江口邻近海域赤潮发生区基本特征初探. 应用生态学报, 14(7): 1031—1038
项有堂, 辛士河, 王东衬等, 1994. 东海黑潮区海水无机氮的分布特征及成因探讨. 海洋通报, 13(1): 35—43
赵卫红, 王江涛, 李金涛等, 2006. 长江口及邻近海域冬夏季浮游植物营养限制及其比较. 海洋学报, 28(3): 119—126
胡明辉, 杨逸萍, 徐春林等, 1989. 长江口浮游植物生长的磷酸盐限制. 海洋学报, 11(4): 439—443
洪华生, 2012. 中国区域海洋学—化学海洋学. 北京: 海洋出版社, 171—217
高生泉, 林以安, 金明明等, 2004. 春、秋季东、黄海营养盐的分布变化特征及营养结构. 东海海洋, 22(4): 38—50
曹欣中, 1986. 浙江近海上升流季过程的初步研究. 水产学报,10(1): 51—56
谢琳萍, 孙 霞, 王保栋等, 2012. 渤黄海营养盐结构及其潜在限制作用的时空分布. 海洋科学, 36(9): 45—53
Chen C T A, 1996. The Kuroshio intermediate water is the major source of nutrients on the East China Sea continental shelf.Oceanologica Acta, 19(5): 523—527
Fisher T R, Peele E R, Ammerman J W et al, 1992. Nutrient limitation of phytoplankton in Chesapeake Bay. Marine Ecology Progress Series, 82(1): 51—63
Harrison P J, Hu M H, Yang Y P et al, 1990. Phosphate limitation in estuarine and coastal waters of China. Journal of Experimental Marine Biology and Ecology, 140(1—2):79—87
Hecky R E, Kilham P, 1988. Nutrient limitation of phytoplankton in freshwater and marine environments: a review of recent evidence on the effects of enrichment. Limnology and Oceanography, 34(4): 796—822
Justić D, Rabalais N N, Turner R E, 1995. Stoichiometric nutrient balance and origin of coastal eutrophication. Marine Pollution Bulletin, 30(1): 41—46
Liu S M, Zhang J, Chen S Z et al, 2003. Inventory of nutrient compounds in the Yellow Sea. Continental Shelf Research,23(11—13): 1161—1174
Ryther J K, Dunstan W M, 1971. Nitrogen, phosphorus, and eutrophication in the coastal marine environment. Science,171(3975): 1008—1013
Wang B D, Wang X L, Zhan R, 2003. Nutrient conditions in the Yellow Sea and the East China Sea. Estuarine, Coastal and Shelf Science, 58(1): 127—136
