基于结构方程模型的云南松次生林林木多样性影响因子1)
2015-03-07舒树淼赵洋毅段旭胡慧蓉熊好琴
舒树淼 赵洋毅 段旭 胡慧蓉 熊好琴
(西南林业大学,昆明,650224)
责任编辑:潘 华。
现代森林经营方法指出,森林的林分结构是生长过程中的驱动因子,对森林发展具有决定性作用[1]。其中林分结构包括了林分非空间结构以及林分空间结构,并受到立地条件的影响。于是森林在林分结构和立地条件的共同作用下呈现出不同状态。如表现出不同的多样性[2-3]。虽然长期以来对多样性的形成原因是森林生态学研究的主要内容之一[4]。但大多都集中在生境因子等角度,而从林分特征因子角度对多样性影响的研究相对较少[5]。特别是在对林木大小多样性上的研究更是少有报道。同时,对林分因子作用的研究大多集中在一对一的直接关系上,因此并不能较全面概括出生森林生态系统作为系统的固有复杂性[6],也不能为研究的系统提供全面信息[7]。故需从系统的结构层次入手,综合考虑这些因子的相互作用关系是描述系统行为的必要基础。
结构方程模型是广泛应用于社会学的因果检验模型[8]。可通过较易数量化的外显变量来估计无法直接观测的潜变量,进而分析潜变量之间的联系。因此,结构方程模型可以揭示系统的内在因果联系,并且对这种联系的描述不仅包含直接作用,还包括间接作用。在本文中,林分非空间结构、林分空间结构、立地条件均是不能直接观测的潜变量,故可通过其他若干较易数量化的外显变量进行估计,这满足结构方程模型的计测思想,进而使得分析这些潜变量之间的相互作用成为可能。
云南松作为滇中高原地区典型乡土树种,对滇中地区生森林生态环境有着至关重要的作用,并以同龄纯林为代表[9]。因此,本文选择林龄相似的云南松次生纯林为研究对象,探讨不同立地条件下林分结构对林木大小多样性的影响。且林木大小多样性包括了胸径多样性、树高多样性等内容。故本研究在一定程度上也反应了林分结构对胸径、树高分布的影响。即通过结构方程模型,分析了立地条件、林分结构对林木胸径、树高多样性或分布的影响,可对云南松的经营和管理提供理论依据。
1 研究区概况
云南磨盘山国家森林公园地处云贵高原、横断山地和青藏高原南缘的地理结合部。地理位置为北纬23°46'~23°54',东经101°16'06″~101°16'12″,海拔1 260.0~2 614.4 m,是云南亚热带北部与亚热带南部的气候过渡地区,又有着典型的山地气候特点,年平均气温15 ℃,年平均降水量为1 050 mm,极端最高气温33.0 ℃,极端最低气温2.2 ℃,全年日照时间2 380 h。磨盘山土壤以第三纪古红土发育的山地红壤和玄武岩红壤为主,高海拔地区有黄棕壤分布,土壤厚度以中厚土壤层为主,局部为薄土层,土层厚度1 m 左右。
2 研究方法
2014年9—11月选择磨盘山林龄相近的典型云南松天然次生纯林固定大样地8 个(每个100 m×100 m),立地条件存在差异,各样地基本情况见表1。在每块大样地中设置若干临时标准小样地(20 m×20 m),共计100 个。并在各个小样地中进行每木检尺,调查树木的胸径、树高、位置、年龄等。采用Shannon-Wiener 指数计算小样地内胸径多样性、树高多样性[10]。公式为:
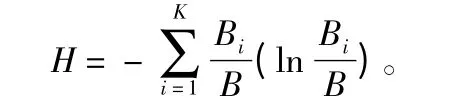
其中,H 为胸径多样性(Hd)或树高多样性(Hh);B为胸径断面积总和;Bi为不同径阶等级或树高等级下的胸径断面积总和;K 为径级或树高等级数目。胸径以2 cm 为一个等级,树高以2 m 为一个等级。若将样地内林木的分布结构视为分形体,则可采用分形维数表征小样地中林木对空间的占据情况[11]。公式为:
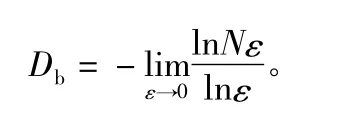
其中,Db为分形维数;ε 为样地中格子的边长,N(ε)表示不同尺度格子中树木的个数,体现了尺度划分。本文中将每个小样地从2 等分至20 等分,即格子尺度从10 m×10 m 到1 m ×1 m。同时采用平均大小比数、平均角尺度衡量林分空间结构特征,公式为:
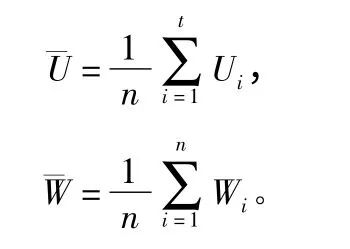
其中,Ui为大小比数,为结构单元中kij的平均值,即如果相邻木j 比参照木比参照木i 胸径小,则kij=1,否则kij=0。Wi为角尺度,为结构单元中Zij的平均值,即如果第j 个α 角小于标准角α0,Zij=1,否则Zij=0。平均大小比数和平均角尺度分别衡量了林木个体大小和分布格局等情况[12],对复杂森林的空间结构有很强的解析能力[13]。
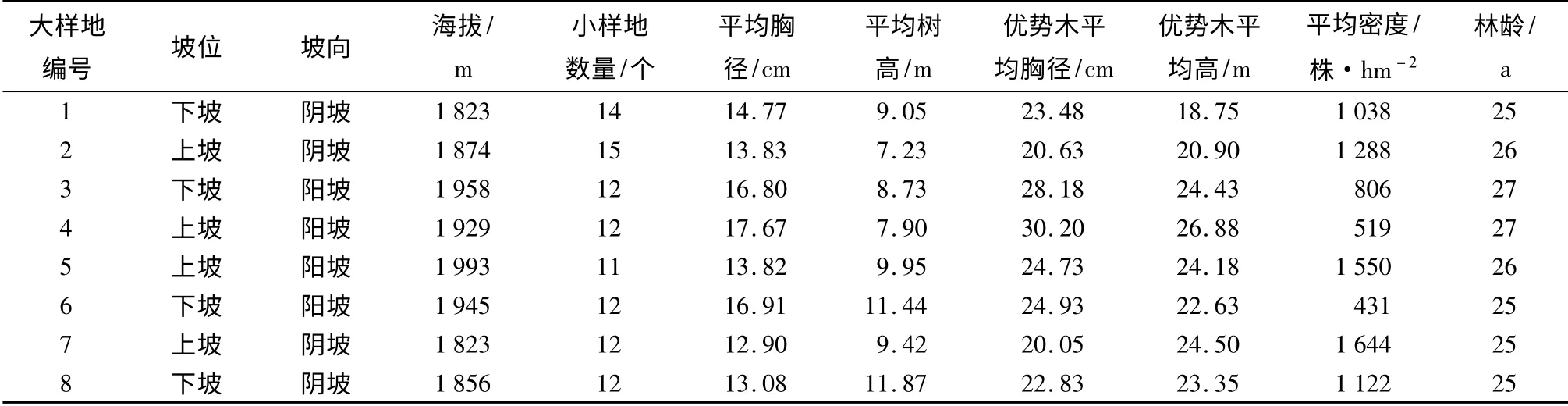
表1 样地基本情况
根据结构方程模型,潜变量是指那些不能被直接观测的变量,往往通过显变量对其进行估计。在森林生态系统中,如胸径、树高、冠幅等指标都是可被直接测量的指标,而通过对这些显性指标的了解便可以估计出林木大小,于是便能基于林木大的意义来进行后续讨论。结构方程模型分为测量模型部分和结构模型部分。测量模型是建立潜变量与外显变量之间关系的模型。
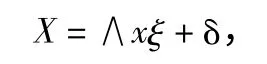
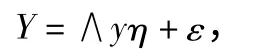
式中:X 为外生显性变量向量为观测值;Y 为内生显性变量向量为观测值;∧x 和∧y 为指标变量(X,Y)的因素负荷量;δ、ε 为外生显性变量与内生显性变量的测量误差;ξ 为外生潜变量表示原因,η 为内生潜变量表示结果。而结构模型则可反映各潜在变量之间的关系。
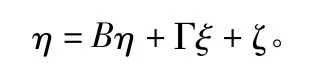
式中:B 为内生潜在变量之间关系的结构系数矩阵;Γ 为内生潜在变量与外生潜在变量之间关系的结构系数矩阵;ζ 为结构模型中干扰因素或残差值。根据此模型和最大似然法便可建立结构方程模型路径图。这里将林分立地条件视为外生潜变量(ξ1)。因各样地林分密度较小且与优势木胸径之间无显著负相关(P=1.15 >0.05),可以认为密度尚未明显影响优势木胸径。故立地条件由优势木胸径(x1)和优势木树高(x2)计测[6,14]。将林分非空间结构(ξ2)、空间结构(ξ3)视为外生潜变量。其中与之相关的观察变量分别为:大小比数(x3)、角尺度(x4)、分形维数(x5);平均胸径(x6)、平均树高(x7)、每个小样地中林木个体数量(x8);将多样性(η)视为内生潜变量,其中胸径多样性(η1)、树高多样性(η2)与之相关。δ、ε、ζ 均为对应残差。使用SPSS 和AMOS 进行结构方程路径分析。
3 结果与分析
3.1 林分非空间结构与空间结构相关性
将林分空间结构与非空间结构进行基于结构方程模型的相关分析,结果如图1、表2。卡方自由度之比:X2/df=9.8/8 <3,且P =0.270 >0.05,故接受虚无性假设,即观察数据与模型适配。模型的拟合结果均达到各项检验要求。由图1可知,林分空间结构和林分非空间结构存在负相关的路径系数-0.63,P=0.046 <0.05,为显著关系。这表明随着林分空间结构的变化,非林分空间结构将发生一定负相关变化。在该模型的关系下,角尺度、大小比数的因子载荷量分别为:-0.30、-0.47。其系数大小表明了它们受到林分空间结构的影响大小且为负。而林分空间结构对分形维数为正影响。当林分空间变化1 时,三者分别变化-0.30、-0.47、0.77。即三者对空间结构的估计有明显贡献。同理,就在载荷量来看,平均胸径、平均树高和个体数量也能较好的估计林分非空间结构。
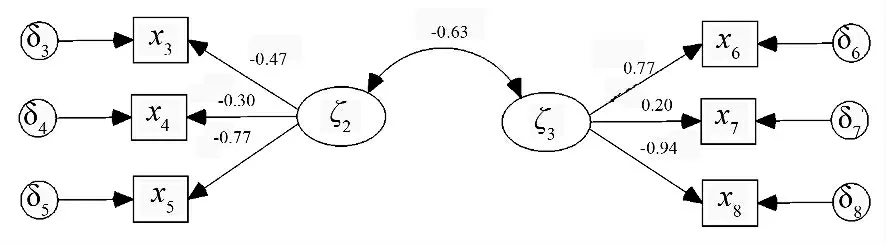
图1 结构模型方程模型1
3.2 林分结构对林木大小多样性结果的影响
立地条件直接影响林分结构和林分多样性[15]。而林分结构作为长期驱动森林生长的重要因子,在响应立地条件,影响林分生长的同时也影响林分多样性[16]。因而可以建立立地条件、林分结构影响林分多样性中林木大小多样性的结构方程模型。结果如图2所示。其卡方自由度之比为:X2/df =37.5/26<3,P=0.067 >0.05,故接受虚无假设,即模型与观测数据适配。其他拟合指标见表2。
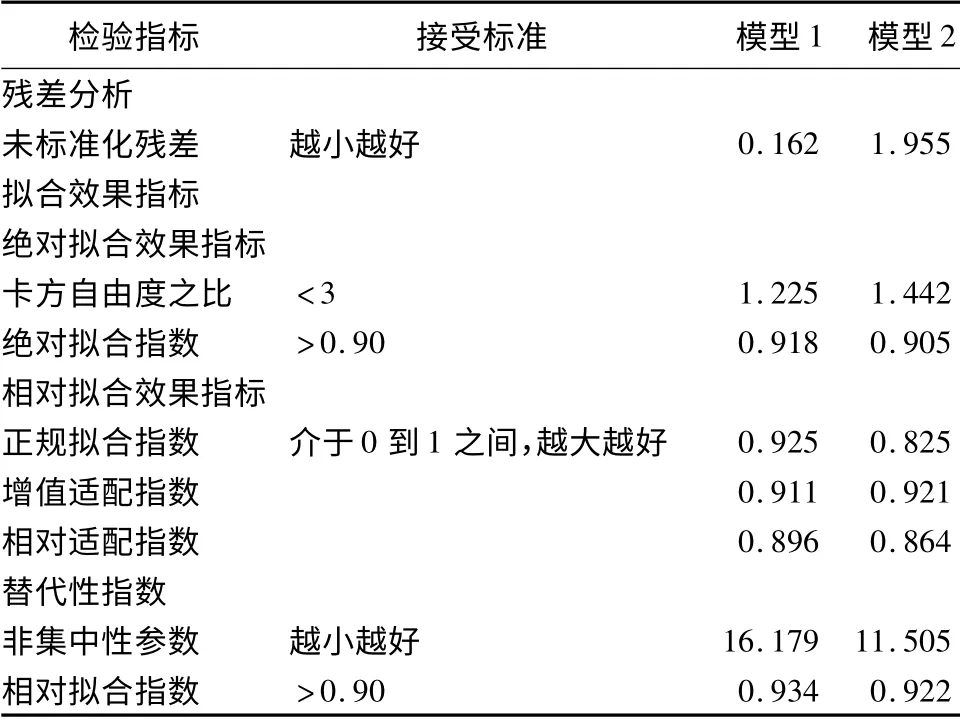
表2 结构方程模型拟合结果检验
由图2知,立地条件对林空间结构、非空间结构的影响系数为:-0.35、0.54,即立地条件变化1,林分空间结构变化-0.35,林分非空间结构变化0.54。直接对多胸径样性、树高多样性的影响系数分别为:0.23、0.77,均为正影响。可知环境因子直接对树高的多样性的影响较胸径多样性大。这表明树高对环境的响应更为明显。这与王树立等的研究相似[6]。在王树力等的结构方程模型中树高多样性对林分多样性的因子载荷量为1.044,大于胸径多样性0.540。而立地环境对林分结构的影响系数为0.657,于是当立地环境变化1 时,则树高多样性变化0.657×1.044 =0.686,胸径多样性变化0.657 ×0.540 =0.355。
林分结构中,空间结构对胸径多样性的影响系数为0.52,对树高多样性的影响系数为0.81。即空间结构更容易影响树高多样性的变化,为正影响。另外,非空间结构对胸径的影响系数为-0.44,对树高多样性的影响系数为-0.18,意味着非空间结构更易影响胸径多样性的变化,为负影响。
标准化总影响系数表明,胸径多样性主要受到立地条件的影响,影响系数为0.46,而树高多样性主要受到林分结构的影响,影响系数为0.63。另外,立地条件对多样性的总影响系数为0.85。林分结构对多样性的总影响系数为0.71(表3)。
最后需要指出的是对于森林生态系统而言许多特征因子之间并不孤立。如本文中云南松个体数量的多少还表现出对林木对林分空间的填充程度,直接影响分形维数。在图2中表示了这种关系,即x8对x5存在影响,其系数为0.40。若立地条件变化1,则分形维数将变化-0.27。这可能表明了在较好的立地条件下,云南松倾向于增加现有个体的大小。而当立地条件不太适宜的时候,云南松开始倾向于r 对策即采取繁殖策略使得个体数量增加,至使分形维数变大。事实上云南松因种子繁殖的方式为r对策生物[17]。另外,这一现象还可能和种内竞争存在关系,即因为立地条件较好,聚集分布并没导致强烈竞争,使得聚集可以维持。可见分形维数还可综合反映林分结构、立地条件等综合指标。
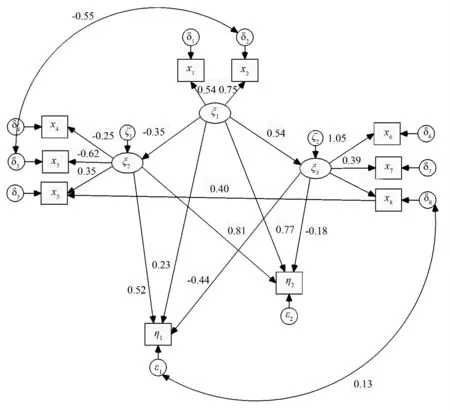
图2 结构模型方程模型2
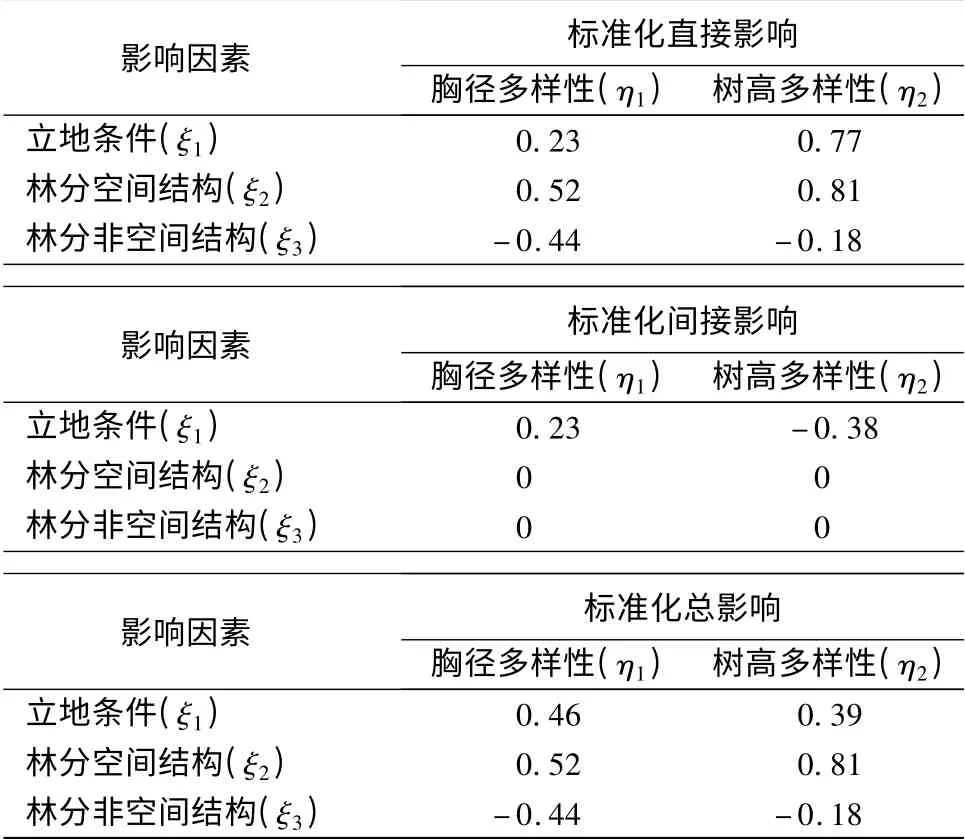
表3 云南松次生林结构方程模型标准化影响系数
4 结论与讨论
生物多样性被认为与生态系统的功能与稳定有密切关系[18-19],已成为生态学领域研究的一个重大科学问题。包括了物种数目、个体数量、分布、相互关系等内容[20]。在云南松纯林中,个体差异的分布是多样性的主要内容。于是本文通过胸径、树高多样性衡量了这种复杂性的差异。并对磨盘山云南松天然次生纯林进行了基于结构方程模型的林分因子分析。测量模型的结果表明:林分空间结构和林分非空间结构存在显著负相关,相关系数为-0.63。各观测变量均能较好反反应隐变量。而结构模型则表明:立地条件对胸径多样性、树高多样性除了直接影响作用外还存在间接影响作用。其中对树高多样性的间接影响为负影响,影响系数为-0.38。总体而言,对胸径多样性影响最大的是立地条件;对树高多样性影响最大的是林分结构。林分结构又分为空间结构和非空间结构。其中非空间结构对胸径、树高多样性为负影响,并对胸径影响较大;空间结构对胸径、树高多样性为正影响,并对树高影响较大。陈学群[21]的研究指出随着林分密度的增加,林分平均胸径减小,直径分布的离散程度就越大,树高却不易受到这种影响。这表明非空间结构易影响胸径,而非树高。在本研究中认为林分密度受林分非空间结构的影响,而当林分密度变化1 时,林分非空间结构需变化-1.45。其对胸径多样性、树高多样性的影响系数为:0.64、0.26。这表密度使胸径分布更加离散。
郑景明[22]等的研究表明垂直复杂性和树高多样性存在显著正相关,显然空间结构更易影响垂直复杂性。与本文结论类似。另外,立地条件对多样性的影响大于林分结构,这与王树力[6]等的研究一致。并由此猜测上述林分特征因子的变化关系可能并不受树种的影响。
生态系统是一个拥有耗散结构的非线性开放系统[23],拥有固有复杂性[24]。可见,为了描述系统复杂行为并不能单靠一对一的简单关系来直接描述。本文从云南松的立地条件、林分结构等特征因子出发,通过结构方程模型探究了二者对林木大小多样性的影响。和以往的研究相比,细化了林分空间结构和非空间结构,分析了上述因子对多样的直接和间接作用。并没有直接讨论平均胸径、平均树高、大小比数、角尺度等林分因子对多样性的影响。而是将它们视为受较高系统层次影响的观察变量。即可以通过观测变量来估计位于系统主要结构位置的隐变量,进而探讨隐变量之间的联系。这正是结构方程模型的重要思想[8]。其对系统的解构和分析都有较强能力。然而结构方程模型还没有广泛运用到自然科学领域。目前国内外在草地、湿地、森林生态方面有一定应用[6,25-27]。
本文选择的研究对象为滇中地区典型乡土树种云南松,且80%为纯林[28]。故存在组成简单、生物多样性低、林分稳定性差等特点。虽然对于云南松已有大量研究,但对云南松纯林中特别是林木大小多样性与林分因子关系的基础研究却少有报道[28]。本文从不同立地出发,就典型云南松纯林进行了基于结构方程模型的林分因子分析,以期较全面地揭示立地条件、林分结构与林木大小多样性之间的耦合关系。
[1] 惠刚盈,赵中华,胡艳波.结构化森林经营技术指南[M].北京:中国林业出版社,2010.
[2] 雷相东,唐守正.林分结构多样性指标研究综述[J].林业科学,2002,38(3):140-147.
[3] 郝凌颖.紫金山不同坡位林分结构与生物多样性研究[D].南京:南京林业大学,2012.
[4] Berger U,Adams M,Grimm V,et al.Modelling secondary succession of neotropical mangroves:causes and consequences of growth reduction in pioneer species[J].Perspectives in Plant Ecology Evolution & Systematics,2006,7(4):243-252.
[5] Kraft L S,Crow T R,Buckley D S,et al.Effects of harvesting and deer browsing on attributes of understory plants in northern hardwood forests,Upper Michigan,USA[J].Forest Ecology &Management,2004,199:219-230.
[6] 王树力,周健平.林用基于结构方程模型的林分生长与影响因子耦合关系分析[J].北京林业大学学报,2014,36(5):7-12.
[7] Grace J B.Structural equation modeling and natural systems[M].Cambridge:Cambridge University Press,2006.
[8] 吴明隆.结构方程模型—AMOS 的操作与应用[M].重庆:重庆大学出版社,2009.
[9] 廖声熙.滇中云南松林景观格局、林分结构及近自然经营模式研究[D].北京:中国林业科学研究院,2008.
[10] Wang S,Chen H Y H.Diversity of northern plantations peaks at intermediate management intensity[J].Forest Ecology & Management,2010,259(9):360-366.
[11] 马克明,祖元刚.兴安落叶松种群格局的分形特征:计盒维数[J].植物研究,2000,20(1):104-225.
[12] 周红敏,惠刚盈,赵中华,等.森林结构调查中最适样方面积和数量的研究[J].林业科学研究,2009,22(4):482-485.
[13] 惠刚盈,Gadow K V,胡艳波.林分空间结构参数角尺度的标准角选择[J].林业科学研究,2004,17(6):687-692.
[14] 马建路,宣立峰,刘德君.用优势树全高和胸径的关系评价红松林的立地质量[J].东北林业大学学报,1995,23(2):20-27.
[15] 闫淑英,席青虎,铁牛.寒温带杜香-兴安落叶松林林分健康评价研究[J].南京林业大学学报:自然科学版,2010,34(6):81-86.
[16] 赵洋毅.缙云山水源涵养林结构对生态功能调控机制研究[D].北京:北京林业大学,2011.
[17] 虞泓.云南松及其近缘种生态遗传与进化研究[D].北京:中国科学院植物研究所,1999.
[18] De G L,Bergeron Y,De G L.Diversity and stability of understorey communities following disturbance in the southern boreal forest[J].Journal of Ecology,2012,90(6):777-784.
[19] Grime J P.Benefits of plant diversity to ecosystems:immediate,filter and founder effects[J].Journal of Ecology,1998,86(6):902-910.
[20] 张金屯,范丽宏.物种功能多样性及其研究方法[J].山地学报,2011,29(5):513-519.
[21] 陈学群.不同密度30年生马尾松林生长特征与林分结构的研究[J].福建林业科技,1995,22(增刊):40-43.
[22] 郑景明,赵秀海,张春雨.北京百花山森林群落的结构多样性研究[J].北京林业大学学报,2007,29(1):7-11.
[23] 普里戈金.从混沌到有序[M].上海:上海译文出版社,2005.
[24] 武显微,武杰.从简单到复杂——非线性是系统复杂性之根源[J].科学技术哲学研究,2005,22(4):60-65.
[25] Laughlin D C,Abella S R,Covington W W,et al.Species richness and soil properties in Pinus ponderosa forests:A structural equation modeling analysis[J].Journal of Vegetation Science,2009,18(2):231-242.
[26] 徐咪咪.异龄林林分生长的结构方程模型分析研究[D].北京:北京林业大学,2010.
[27] 王酉石,储诚进.结构方程模型及其在生态学中的应用[J].植物生态学报,2011,35(3):337-344.
[28] 蔡年辉.云南松群落动态特征及其在近自然改造中的应用[D].昆明:西南林学院,2007.
