淮南采煤沉陷区内小型塌陷湖泊轮虫群落结构特征*
2015-02-25王晓萌易齐涛
王晓萌,易齐涛,2,徐 鑫,李 慧,章 磊
(1:安徽理工大学地球与环境学院,淮南 232001)
(2:中国科学院生态环境研究中心,北京 100085)
淮南采煤沉陷区内小型塌陷湖泊轮虫群落结构特征*
王晓萌1,易齐涛1,2**,徐鑫1,李慧1,章磊1
(1:安徽理工大学地球与环境学院,淮南 232001)
(2:中国科学院生态环境研究中心,北京 100085)
摘要:在淮南潘谢矿区内设置3个水文生态环境条件差异较大的小型煤矿塌陷湖泊研究站点,分别为潘谢潘集站(PXPJ)、潘谢顾桥站(PXGQ)和潘谢谢桥站(PXXQ),于2013-2014年4个季度分别对这3个湖泊的轮虫群落结构组成特征及水生态环境因子影响因素进行分析.共观察到轮虫15属32种,其中PXPJ站点13属25种,PXGQ站点13属24种,PXXQ站点11属20种.3个塌陷湖泊轮虫群落结构具有较大的时空分布差异,主要优势种为曲腿龟甲轮虫、罗氏异尾轮虫、前节晶囊轮虫、卜氏晶囊轮虫和针簇多肢轮虫.PXPJ、PXGQ、PXXQ站点轮虫丰度范围分别为500~2800、950~3350、400~3900ind./L,3个站点Shannon-Wiener多样性性指数的平均值分别为2.71、2.68和2.54,体现了塌陷湖泊β中污染型水质环境的特点.轮虫群落结构与环境因子的冗余分析结果表明:水温、电导率和营养盐等环境因子对淮南采煤塌陷湖泊的轮虫群落结构影响较大.
关键词:采煤沉陷区;塌陷湖泊;轮虫;水环境因子
*国家自然科学基金项目(41202242)和中国博士后科学基金项目(2014M560127)联合资助.2014-08-04收稿;2014-12-04收修改稿.王晓萌(1993~),女,硕士研究生;E-mail:wxsprout2014@sina.com.
安徽省淮南市是我国重要的煤炭基地之一,由于采煤造成了大面积土地沉陷和积水,调查资料表明淮南矿区2010年沉陷区面积约140km2,积水区面积占30%~50%,并呈快速扩张的趋势,预计至2020年淮南沉陷区面积将增至200km2以上,可储备约7×108~10×108m3的淡水资源,对区域社会、经济及环境的可持续发展意义重大[1].根据淮南市政府和淮南矿业集团规划,淮南采煤沉陷积水区将建成集防洪、渔业活动、工农业供水、生态湿地等综合功能于一体的大型平原水库或湖泊群.
受矿区煤炭开采边界限制和人为改造活动因素影响,沉陷区水域分别形成了水塘、湖泊、湿地或平原水库等不同景观类水体,其中以小型塌陷湖泊水体为主(单体面积范围一般为1~10km2).这些塌陷湖泊地处农业活动中心区域或城镇生态恢复带上,分别执行着水源保护地、渔业活动区、湿地生态恢复区等不同的水体功能,水质和营养状况差异较大[2],从而导致浮游生物群落结构对湖泊生态环境表现出不同的响应情况.从区域尺度来看,小型塌陷湖泊与矿区水文生态环境密切相关,各分矿区水文情势、生态环境与人为活动等相关特征很大程度将会在区内湖泊生态系统中得以体现,湖泊生态系统的结构与功能表征、其对水体富营养化进程响应过程和机制及相关生态环境效应,是矿区生态环境重建、恢复及评价理论中面临的重要科学问题.
轮虫作为浮游动物的重要组成部分之一,处于食物网的中间环节,起着承上启下的作用.它一方面可以作为捕食者调控浮游植物、原生动物和细菌等生物的数量,另一方面可以作为鱼类的开口饵料和食物来源,同时轮虫繁殖迅速,物质循环和能量流动效率较高,在水生态系统营养结构中起着重要作用[3-4].目前,国内已有不少有关轮虫与环境因子关系的研究,从中可以发现在不同湖泊中影响轮虫群落结构的环境因子主要包括水体理化特征、营养盐和甲壳类浮游动物等,如夏品华等[5]研究表明水温、氨氮、电导率和pH是影响红枫湖水库冬、春季轮虫分布的主要因子,舒婷婷等[6]在对玄武湖的研究中发现pH、浮游甲壳动物丰度、总悬浮物和总溶解性氮是影响夏季轮虫水平分布的重要因子,其他湖泊中也有诸多相关研究.有关塌陷湖泊轮虫方面的研究仍然较为欠缺,此前,金显文等[7]研究发现淮北煤矿塌陷水体的轮虫群落结构变化特征与其他富营养化水体显著不同.由于区域地理气候方面的差异,淮南与淮北矿区塌陷湖泊水化学特征存在显著差异[8],进而可能造成水生态系统包括轮虫在内的生物群落结构方面的差异.此外,淮南采煤塌陷湖泊相比淮北具有更大的水域规模和初级生产力[9],具有较大的渔业生产潜力,轮虫数量的变动很大程度上会影响鱼类的存活率,通过研究湖泊中轮虫群落结构与水质环境因子之间的关系,可以对淮南塌陷湖泊渔业资源的合理开发和管理利用提供科学依据.与此同时,轮虫对水质变化反应敏感,常作为水质变化的指示因子,监测和评价水体的污染程度和营养状况.因此对淮南矿区塌陷湖泊轮虫群落结构进行研究不仅可以深入理解这一区域性水域生态系统结构特征及其对矿区水生态环境因子响应情况,而且可以比较两淮矿区的区域性差异,为矿区水生态环境重建、恢复与保护提供基础资料和相关理论指导.
1 材料与方法
1.1 研究区域概况与站点
选取潘谢矿区内塌陷湖泊为研究对象.潘谢矿区也是淮南煤矿基地中沉陷面积最大、积水最为集中的区域,其位于安徽省淮河北岸的冲击平原,矿区东西长近58km,南北宽为6~25km,面积约为865km2.矿区内地表水系较为发达,天然河道有西淝河、港河、黑河、济河、泥河和架河,人工河道有永幸河,均由西北向东南流入淮河.分别在矿区潘谢东部的潘集矿区(PXPJ)、中部的顾桥矿区(PXGQ)和西部的谢桥矿区(PXXQ)内各选1个小型塌陷湖泊站点为研究对象进行相关研究(图1).

图1 淮南潘谢矿区沉陷积水区及研究湖泊站点分布Fig.1 Distribution of the subsidence areas and studied lakes around the Panxie coal mine area around Huainan
3个小型塌陷湖泊沉陷积水历史及水文条件均存在较大差异,其中PXPJ站点位于潘集矿区西北侧,与流经矿区的河流泥河常年相通;PXGQ站点位于顾桥矿西北侧,水体较为封闭(无入湖河流);而PXXQ站点位于谢桥矿西北侧沉陷区内,其通过节制闸与其南边的济河连通,在汛期接纳济河季节性引水.3个塌陷湖泊均为失地农民所利用进行渔业活动,每年有鱼苗投放,但一般无饵料和肥料施加,3个湖泊面积、水深、容积及形成年龄等具体信息见表1.
1.2 样品采集与测定
于2013-2014年分别对PXPJ、PXGQ和PXXQ站点分4个季度进行采样和分析,在每个研究站点设置7~9个水质采样点,每个水质采样点用有机玻璃采水器采取表层水样保存待分析.水质调查及分析的指标包括:水温(Tem)、电导率(EC)、透明度(SD)、pH、溶解氧(DO)、碱度(Alk)、总悬浮颗粒物(TSS)、化学需氧量(CODCr)、总氮(TN)、总磷(TP)、叶绿素a(Chl.a).水温、pH、DO用pH仪(YSI pH100)现场测定,SD用赛氏盘测定.水质分析方法主要参照相关国家标准进行[10],其中碱度用酸碱指示剂滴定法测定,TN浓度用碱性过硫酸钾消解紫外分光光度法测定,TP浓度用钼酸铵分光光度法测定,COD浓度采用重铬酸钾法测定,Chl.a浓度用分子荧光法分析测定.
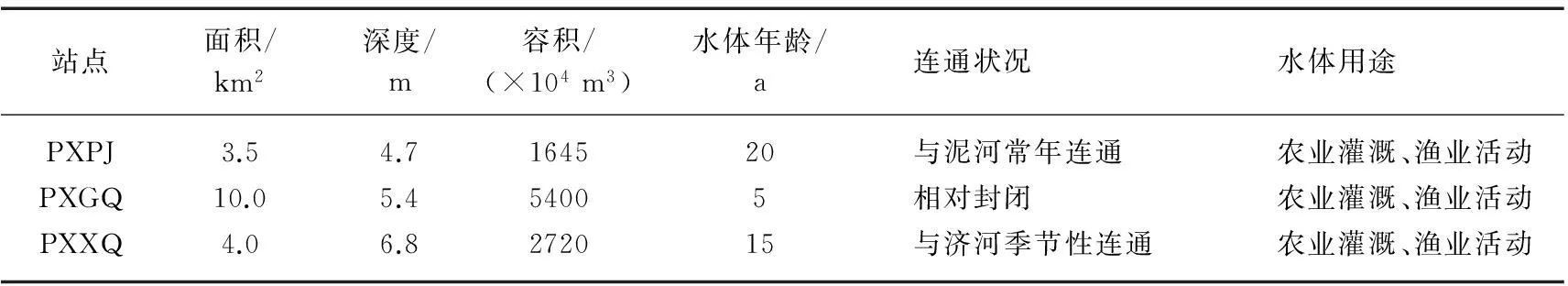
表1 淮南潘谢矿区3个塌陷湖泊水体的主要特征
在各站点湖心附近设置3个浮游生物采样点,采集浮游植物和浮游动物.浮游植物和轮虫定性样品采集用25#(网孔直径64μm)浮游生物网在水体表层中以“∞”字型来回捞取,加福尔马林固定,部分样品作活体观察,用于鉴定种类.对于定量样品,用5L有机玻璃采水器根据水深在上、中、下层采集进行混合,然后装入1000ml的采集瓶,分别加入鲁哥试剂和少量福尔马林,沉淀48h后浓缩定容至50ml.甲壳类定性样品采集用13#(网孔直径112μm)浮游生物网在水体表层中以“∞”字型来回捞取,加福尔马林固定,部分样品作活体观察,用于鉴定种类.对于定量样品,用5L有机玻璃采水器根据水深在上、中、下层采取水样进行混合,经25#(64μm)浮游生物网过滤,加福尔马林固定.浮游生物种类鉴定及观察计数参考文献[11].
1.3 数据分析方法
运用浮游动物Jaccard相似性指数(X)[12]、Shannon-Wiener多样性指数(H′)[13]、均匀度指数(J)和优势种的优势度(Y)对塌陷湖泊轮虫进行生态学分析评价,各指数计算公式分别为:
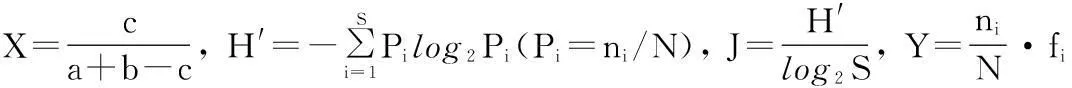
式中,ni为站位中i物种的个数,N为站点中轮虫总个数,fi为i物种在各站点中出现的频率,S为站点中轮虫总种数,a为站点A中出现的轮虫种类数,b为站点B中出现的轮虫种类数,c为站点A和站点B中都出现的轮虫种类数.
采用SPSS 16.0软件对不同湖泊站点及不同季节数据进行差异显著性分析;选择3个站点中相对密度>2%的轮虫代表种进行环境因子影响分析,将轮虫丰度值和理化数据通过lg(x+1)转换使数据趋于正态分布,用Canoco 4.5软件对该数据进行去趋势对应分析(detrended correspondence analysis,DCA),得出第一轴的梯度长为2.109,因此选择典型冗余分析(redundancy analysis,RDA)并通过蒙特卡罗置换检验(monte carlo permutation test)对环境因子进行筛选,绘制物种与环境因子关系图[14-15].
2 研究结果
2.1 调查期间水质特征
调查期间PXPJ站点TP、TN和Chl.a浓度的季节变化范围分别为0.085~0.120mg/L、0.79~2.08mg/L和8.5~40.8mg/m3;PXGQ站点TP、TN和Chl.a浓度均比PXPJ站点低,TP季节变化范围为0.038~0.140mg/L,TN浓度季节变化范围为0.63~1.44mg/L,Chl.a浓度季节变化范围为12.2~38.7mg/m3;PXXQ站点TP浓度相对较低,季节变化范围为0.010~0.076mg/L,TN浓度相对较高,季节变化范围为1.73~5.37mg/L,春、夏季从济河引水带来了丰富的营养盐,其Chl.a浓度高达60mg/m3.总体上3个湖泊水体Chl.a浓度均较高,体现了富营养湖泊的典型特征.PXPJ由于泥河外源负荷的长期输入,使其TP浓度要远高于PXGQ和PXXQ站点,而TN浓度大小顺序为PXXQ站点>PXPJ站点>PXGQ站点(表2).

表2 淮南潘谢采煤沉陷区内3个小型塌陷湖泊水质指标
2.2 3个塌陷湖泊轮虫种类组成特征
3个塌陷湖泊4个季度调查期间总共观察到轮虫15属32种,其中PXPJ站点13属25种,PXGQ站点13属24种,PXXQ站点11属20种(表3).PXPJ站点主要由多肢轮属、龟甲轮属和晶囊轮属构成,春、秋、冬季针簇多肢轮虫最多,优势度分别为0.42、0.25和0.42,夏季罗氏异尾轮虫最多(Y=0.29);PXGQ站点主要为多肢轮属、龟甲轮属和晶囊轮属,春、夏和秋季针簇多肢轮虫最多,优势度分别为0.53、0.31和0.31,冬季螺形龟甲轮虫最多(Y=0.51);PXXQ站点主要为多肢轮属、龟甲轮属和臂尾轮属,春、夏、秋和冬季分别以曲腿龟甲轮虫、前翼轮虫、针簇多肢轮虫和螺形龟甲轮虫为主,优势度依次为0.50、0.42、0.30和0.32.
3个塌陷湖泊轮虫优势种组成的季节分布如图2所示.PXPJ站点轮虫第1优势种为针簇多肢轮虫、罗氏异尾轮虫,分别占年平均丰度的34.7%、12.9%,其中针簇多肢轮虫在秋季丰度较高(950ind./L),罗氏异尾轮虫在夏季丰度较高(750ind./L);PXGQ站点轮虫第1优势种为针簇多肢轮虫、螺形龟甲轮虫,分别占年平均丰度的31.0%、14.2%,其中针簇多肢轮虫在夏季丰度较高(1100ind./L),螺形龟甲轮虫在冬季丰度较高(950ind./L);PXXQ站点轮虫第1优势种为曲腿龟甲轮虫、针簇多肢轮虫和前翼轮虫属,分别占年平均丰度的24.4%、20.2%和13.5%,其中曲腿龟甲轮虫在秋季丰度较高(500ind./L),针簇多肢轮虫在秋季丰度较高(1050ind./L),前翼轮虫属在夏季丰度较高(1650ind./L).

表3 研究区内3个小型塌陷湖泊鉴定的轮虫种类*
*+++为优势种,++为常见种,+为少见种.
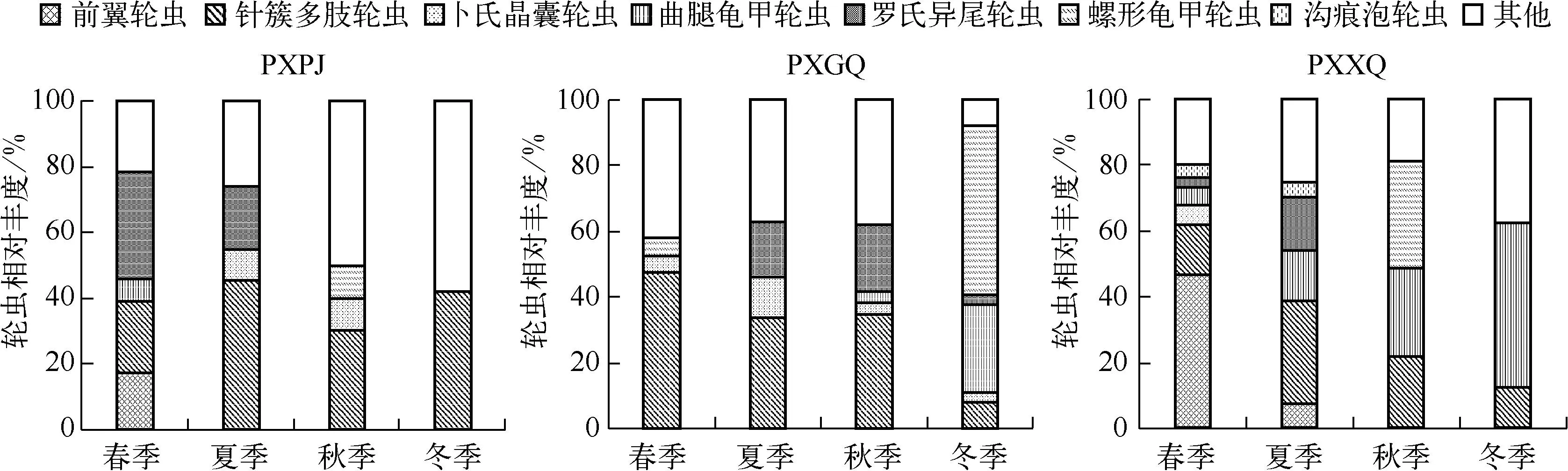
图2 3个塌陷湖泊轮虫优势种组成的季节动态Fig.2 Seasonal dynamics of rotifer dominant species in the three subsided lakes in the research area
2.3 3个塌陷湖泊轮虫丰度季节变化
3个塌陷湖泊4个季度轮虫丰度的变化趋势一致,均值差异分析结果表明,轮虫丰度在不同季节之间的差异显著(P<0.05),夏、秋季轮虫丰度要远高于冬、春季(图3),而3个湖泊站点之间的差异不显著(P>0.05).具体来看,PXPJ站点轮虫丰度范围为500~2800ind./L,夏季轮虫丰度最大,冬季轮虫丰度最小,多肢轮虫属在春、秋和冬季均占较大比例,丰度分别为3500、1050和150ind./L,占轮虫总丰度的58.3%、37.7%和30.0%,异尾轮虫属在夏季占较大比例,丰度高达1100ind./L,占轮虫总丰度的39.3%.

图3 研究区内3个小型塌陷湖泊轮虫丰度季节动态Fig.3 Seasonal dynamic of rotifer abundance in the three subsided lakes of the research area
PXGQ站点轮虫丰度范围为950~3350ind./L,多肢轮虫属在春、夏和秋季均占较大比例,丰度分别为500、1250和1300ind./L,占轮虫总丰度的52.6%、37.3%和40.6%,龟甲轮虫属在冬季占较大比例,丰度高达1450ind./L,占轮虫总丰度的78.4%.
PXXQ站点轮虫丰度在400~3900ind./L之间,龟甲轮虫属在春、冬季占较大比例,丰度分别为200、1100ind./L,占轮虫总丰度的50.0%、59.5%,前翼轮虫属在夏季占较大比例,丰度高达1650ind./L,占轮虫总丰度的42.3%,多肢轮虫属在秋季占较大比例,丰度为1050ind./L,占轮虫总丰度的29.6%.
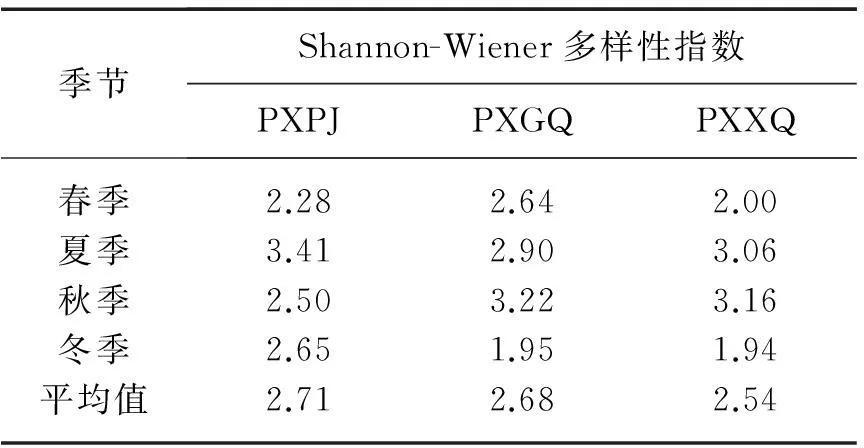
表4 研究区内3个小型塌陷湖泊
2.4 3个塌陷湖泊轮虫多样性与相似性
2.4.1 多样性比较3个站点Shannon-Wiener多样性指数均值分别为2.71、2.68和2.54(表4),站点之间差异较小.PXPJ站点多样性指数季节变化范围为2.50~3.41,夏季最高,而春季最低;PXGQ站点多样性指数季节变化范围为1.95~3.22,秋季最高而冬季最低;PXXQ站点多样性指数季节变化范围为1.94~3.16,夏、秋季较高,而春、冬季最低.
2.4.2 相似性比较调查期间3个站点4个季度轮虫相似性指数X变化范围为0.06~0.63(表5),PXPJ、PXGQ和PXXQ站点相似性指数变化范围分别为0.14~0.25、0.22~0.63和0.18~0.53,差别较大,相似性较差或一般;3个站点在各季节具有一定的相似性,春、夏季PXPJ和PXGQ站点相似性指数最高,相似性指数均为0.52,秋季PXGQ和PXXQ站点相似性指数最高(X=0.50),冬季PXPJ与PXGQ、PXGQ与PXXQ站点之间相似性指数最高(X=0.50).
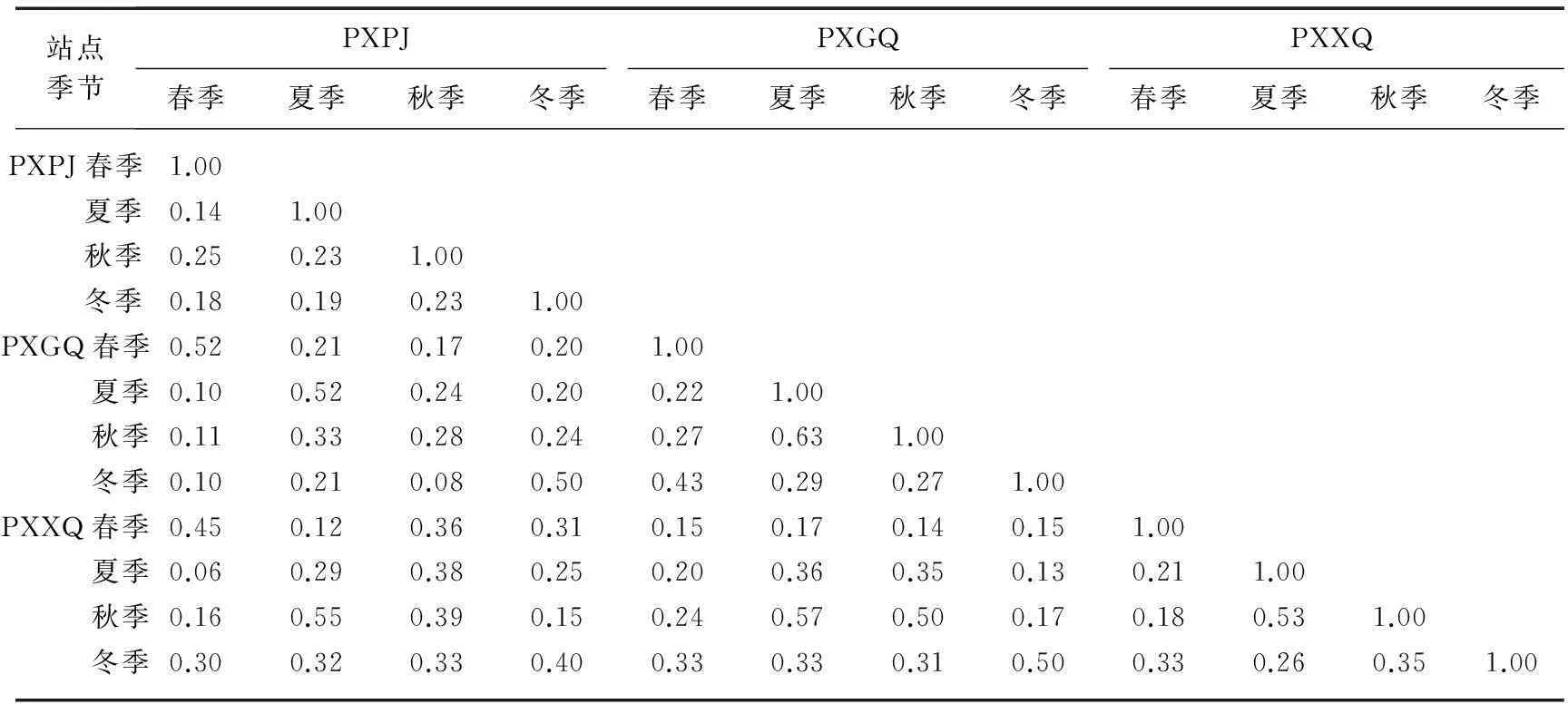
表5 研究区内3个小型塌陷湖泊不同季节轮虫相似性指数
2.5 轮虫群落结构与环境因子的相互关系
根据轮虫出现的频率,选取12种主要的轮虫进行典型冗余分析(RDA),分别为沟痕泡轮虫、罗氏异尾轮虫、前节晶囊轮虫、长三肢轮虫、针簇多肢轮虫、萼花臂尾轮虫、前额犀轮虫、裂痕龟纹轮虫、曲腿龟甲轮虫、前翼轮虫属、疣毛轮虫属、舞跃无柄轮虫.通过RDA前选和蒙特卡罗置换检验(Monte Carlo permutation test)确定Tem、EC、pH、Alk、TP、TN和Chl.a共7个环境因子.

表6 研究区内3个小型湖泊轮虫和
从RDA分析结果可以看出(表6),第1轴的特征值为0.416,说明环境梯度较大,适于排序分析.第1轴环境因子解释物种变异的49.5%,前4轴共解释物种变异的96.1%.
沟痕泡轮虫、罗氏异尾轮虫、裂痕龟纹轮虫以及前翼轮虫属与水温和电导率呈正相关,前节晶囊轮虫、针簇多肢轮虫和舞跃无柄轮虫主要受TP浓度的影响,萼花臂尾轮虫、曲腿龟甲轮虫与TN浓度、Alk呈正相关,长三肢轮虫则主要受SD的影响(图4).总体来看,影响湖泊中轮虫群落结构的主要环境因子为Tem、EC、TN浓度和TP浓度.
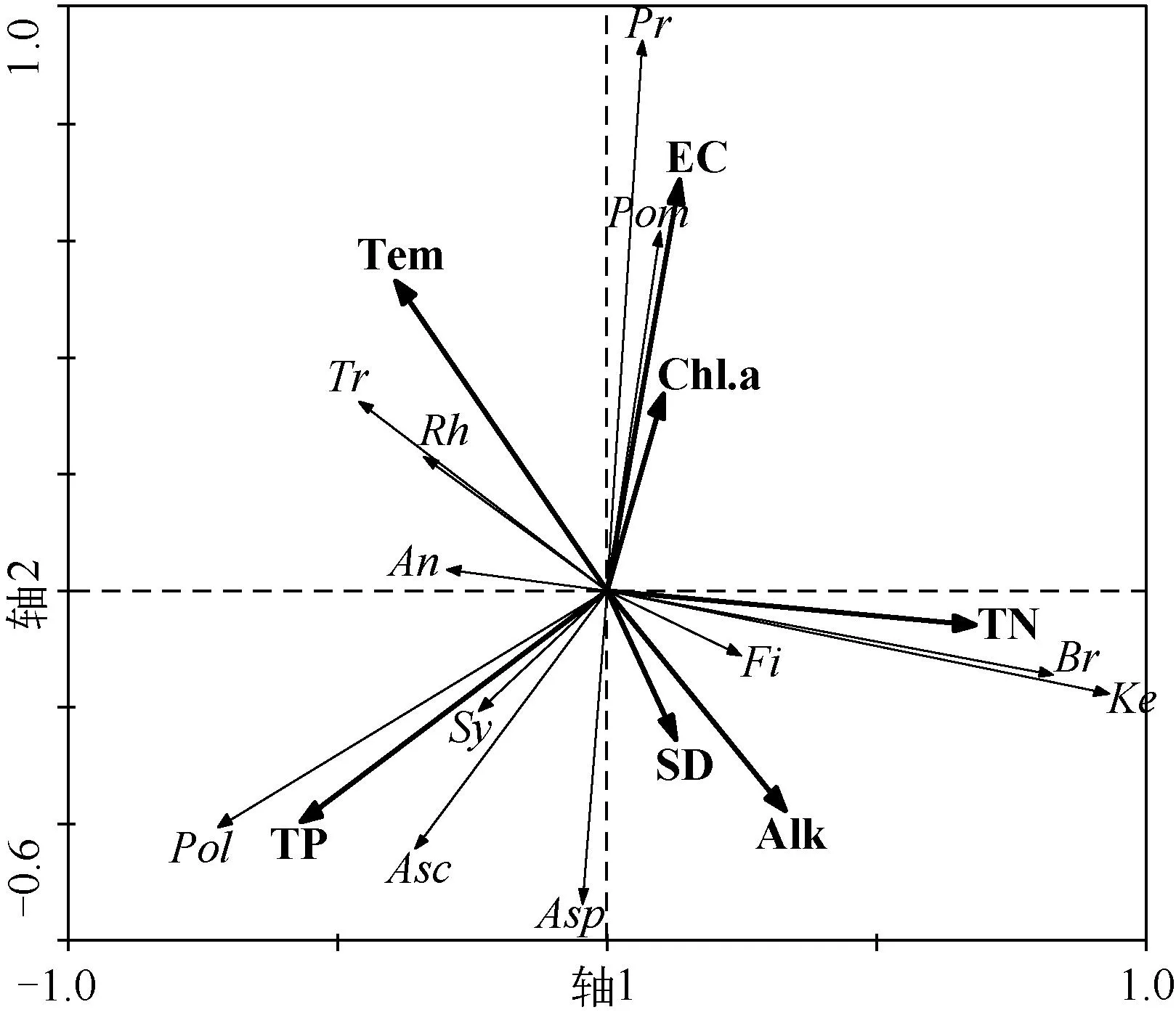
图4 轮虫优势种与环境因子的冗余分析(Pom:沟痕泡轮虫,Tr:罗氏异尾轮虫,Asp:前节晶囊轮虫,Fi:长三肢轮虫,Pol:针簇多肢轮虫,Br:萼花臂尾轮虫,Rh:前额犀轮虫,An:裂痕龟纹轮虫,Ke:曲腿龟甲轮虫,Asc:舞跃无柄轮虫,Sy:疣毛 轮虫属,Pr:前翼轮虫属)Fig.4 Redundancy analysis between dominate species of rotifer and environmental factors
3 讨论
3.1 轮虫群落结构对淮南塌陷湖泊生态环境的指示作用
轮虫对环境反应敏感,常常作为水体的指示生物,因此在评价水体营养状态和污染状况等方面受到广泛的关注[16-17].首先,一般湖泊中常用生物多样性指数来评价水体的污染情况,Shannon-Wiener多样性指数评价标准分为3级:0~1表示重度污染;1~3表示中度污染,其中1~2为α中污染,2~3为β中污染;>3表示轻度污染或无污染[18].PXPJ、PXGQ和PXXQ站点轮虫多样性指数指示为β中污染,表明淮南塌陷湖泊水体已经受到了一定程度的污染,这与水质理化指标表征的结果一致.
其次,随着水体富营养化程度的加剧,采煤塌陷区水体中浮游动物的种类呈明显下降趋势,PXXQ站点轮虫种数低于PXPJ和PXGQ站点,PXXQ站点在引水期间由周围济河引入了污染物浓度较高的河水,特别是有机物和氨氮浓度较高,可能对轮虫的群落结构产生了一定的影响.
再次,从具体的指示物种来看,富营养化湖泊的轮虫主要指示种包括臂尾轮属、裂痕龟纹轮虫、沟痕泡轮虫、扁平泡轮虫、圆筒异尾轮虫、暗小异尾轮虫、长三肢轮虫、螺形龟甲轮虫、矩形龟甲轮虫和针簇多肢轮虫[19-22],而3个小型塌陷湖泊的优势种多为多肢轮属、异尾轮属和龟甲轮属,进一步指示出塌陷湖泊水体的富营养化特征.湖泊富营养化进程能够显著提高轮虫第1优势种的优势度[23],PXPJ站点第1优势种为多肢轮属(Y=0.42),PXGQ站点第1优势种为多肢轮属(Y=0.53),PXXQ站点第1优势种为龟甲轮虫(Y=0.50),在群落中占较高比例.
从两淮区域角度来看,淮北2个采煤塌陷湖泊南湖、乾隆湖一年内共分别鉴定到轮虫9属14种、15属27种,第一优势种均为多肢轮虫属和异尾轮虫属[24],这与本研究的结果基本一致,可能体现了塌陷湖泊富营养化的影响;南湖、乾隆湖轮虫年平均丰度分别为198.4、1916ind./L,其Shannon-Wiener多样性指数平均值分别为0.67、1.99,远低于本研究的3个湖泊,可见采煤塌陷湖泊轮虫的种类和多样性随着水体营养盐浓度水平的升高呈现上升趋势.有研究者曾经提出,在不同水体中任何一个因素的差异都将导致现存轮虫群落结构的不同[25].除了两淮区域地理气候具有差异性,南湖TN、TP浓度分别为0.56、0.06mg/L, 乾隆湖TN、TP浓度分别为0.98、0.15mg/L,营养盐浓度和比例与本研究中的湖泊差异也较为明显.此外,淮北塌陷湖泊碱度比较大,也可能影响轮虫群落结构变化[26].在以上多种因素的影响下,淮南与淮北塌陷湖泊轮虫群落结构既表现出一定的共性,又有所差异.
3.2 影响淮南塌陷湖泊轮虫群落结构的主要因素
作为生态系统营养结构中承上启下的重要功能群,轮虫群落结构主要受生物因素和非生物因素的影响.生物因素主要包括浮游植物、甲壳类浮游动物和鱼类,非生物因素主要包括水温、溶解氧及其它水质指标等.生物因素中,浮游植物作为轮虫的食物之一,其对轮虫的影响不容忽视.
PXPJ站点浮游植物细胞丰度范围为443×104~984×104cells/L,春、夏、秋季较高而冬季最低,以绿藻、硅藻和隐藻门内的种类为主;PXGQ站点浮游植物细胞丰度范围为190×104~1267×104cells/L,蓝藻细胞丰度在夏季和秋季均占据绝对优势,分别占藻类细胞总丰度的61.5%和46.2%,冬、春季浮游植物细胞丰度最小;而PXXQ站点浮游植物细胞丰度范围为557×104~3192×104cells/L,从春季到冬季逐渐减小.总体上浮游植物和轮虫丰度在各站点和季节上变化均具有一致性,即各湖泊春夏季浮游生物丰度相对较高、冬春季相对较低的特点,PXXQ和PXGQ站点浮游植物和轮虫的丰度均高于PXPJ站点(图5).

图5 研究区内3个小型塌陷湖泊浮游植物细胞丰度的季节分布Fig.5 Seasonal distribution of phytoplankton cell abundance in the three subsided lakes of the research area
尽管PXPJ站点水体营养盐浓度较高,但个体较小的绿藻、硅藻和隐藻在浮游植物群落结构中占据较大比例,轮虫或甲壳类浮游动物的捕食压力可能使浮游植物丰度降低.此外,PXPJ、PXGQ和PXXQ站点甲壳类浮游动物年平均丰度分别为8.7、25.95和39.2ind./L,3个塌陷湖泊每年投放大量的滤食性鱼类(鲢、鳙鱼),可能给甲壳类浮游动物带来较大的扑食压力,加上湖泊水体中蓝藻细胞丰度占据优势,其较大个体对甲壳类浮游动物来说并不是适合的食物,从而导致枝角类和桡足类丰度较小,使得轮虫食物竞争或捕食压力较弱,塌陷湖泊浮游植物和浮游动物群落结构体现了富营养化湖泊营养结构的典型特征[27].
从环境因素来看,3个湖泊轮虫丰度和优势种随着季节变化明显,且各湖泊之间相似性较低,可能主要体现了淮南塌陷湖泊水环境因子在时空分布上的差异.RDA分析结果表明,影响轮虫群落的主要环境因子为水温、电导率、TN和TP.许多研究均指出水温是影响轮虫生长、发育、群落组成和丰度变化等极为重要的环境因子[28],水温升高,卵的发育时间缩短,种群周转加快等能够在短时间内形成轮虫的丰度高峰[29],加上食物的充足(由浮游植物细胞密度等表征),塌陷湖泊轮虫丰度及群落结构季节组成很好地体现了这些水质因子的影响.此外,营养盐浓度和比例可能对轮虫群落结构具有较大影响,主要表现为N、P可以通过影响浮游植物从而影响轮虫[30].此前相关研究表明[2],3个塌陷湖泊中PXPJ和PXXQ站点浮游植物生长主要受P限制,而PXGQ站点主要受N限制,这种差异会导致各湖泊浮游植物群落结构组成变化进而影响初级消费者轮虫的群落结构,RDA分析中主要表现为前节晶囊轮虫、针簇多肢轮虫和舞跃无柄轮虫主要受TP浓度影响,而萼花臂尾轮虫、曲腿龟甲轮虫主要受TN浓度影响.值得注意的是,生物因素和水环境因子对轮虫群落结构的影响相互交叉,即通过营养结构中的上行效应(食物或环境等相关因素)或下行效应(捕食压力)来共同调控轮虫的物种组成和丰度变化,主要的调控过程与机制需要进行更加深入的调查和研究.
4 结论
1) 淮南3个塌陷湖泊中共鉴定到轮虫15属32种,其中潘集站点13属25种,顾桥站点13属24种,谢桥站点11属20种.3个湖泊站点轮虫群落结构显示出较大的时空变化特征,轮虫丰度夏、秋季要高于冬、春季,不同站点群落结构差异较大.PXPJ站点第1优势种季节变化为:前翼轮虫属(春季)→针簇多肢轮虫(夏季)→罗氏异尾轮虫(秋、冬季);PXGQ站点第1优势种季节变化为:针簇多肢轮虫(春、夏、秋季)→螺形龟甲轮虫(冬季);PXXQ站点第1优势种季节变化为:曲腿龟甲轮虫(春季)→前翼轮虫属(夏季)→针簇多肢轮虫(秋季)→曲腿龟甲轮虫(冬季).
2) 3个塌陷湖泊由于水文生态环境条件的不同,轮虫多样性、丰度和相似性体现出一定的差异.3个站点Shannon-Wiener多样性指数均值分别为2.71、2.68、2.54,指示β中污染的塌陷湖泊生境.其中PXXQ站点轮虫丰度明显高于PXPJ和PXGQ站点,3个湖泊水体富营养化程度一致;RDA分析表明,淮南煤矿塌陷湖泊轮虫群落结构的主要影响因素是水温、电导率、TN和TP浓度.
参考文献5
[1]易齐涛,孙鹏飞,谢凯.区域水化学条件对淮南采煤沉陷区水域沉积物磷的吸附特征的影响研究.环境科学,2013,34(10):3894-3903.
[2]王婷婷,易齐涛,胡友彪等.两淮采煤沉陷区水域水体富营养化及氮、磷限制模拟实验.湖泊科学,2013,25(6):916-926.
[3]董丽丽,谢力慧.不同法度下新安江水域(屯溪段)轮虫种群动态研究.生态科学,2012,31(2):187-191.
[4]Offem BO,Samsons YA,Omoniyi ITetal. Dynamics of the limnological features and diversity of zooplankton populations of the Cross River System SE Nigeria.KnowledgeandManagementofAquaticEcosystems, 2009, 393(2):1-19.
[5]夏品华,马健荣,李存雄等.红枫湖水库冬春季浮游生物群落与环境因子的典范对应分析.环境科学研究,2011,24(4):378-386.
[6]舒婷婷,孙颖,李静.玄武湖轮虫夏季水平分布及其与环境因子的关系.长江流域资源与环境,2012,21(1):40-45.
[7]金显文,杨文展,邓道贵.淮北煤矿塌陷区水域轮虫的初步研究.生物学杂志,2007,24(5):50-51.
[8]孙鹏飞,易齐涛,许光泉.两淮采煤沉陷积水区水化学特征及控制因素分析.煤炭学报,2014,39(7):1345-1353.
[9]易齐涛,徐鑫,曲喜杰等.两淮采煤沉陷区水体初级生产特征.中国环境科学,2014,34(8):2101-2110.
[10]尹航.环境监测方法标准汇编:水环境.北京:中国标准出版社,2007.
[11]章宗涉,黄祥飞.淡水浮游生物研究方法.北京:科学出版社,1991.
[12]陈家长,孟顺龙,尤洋等.太湖五里湖浮游植物群落结构特征分析.生态环境学报,2009,18(4):1358-1367.
[13]Shannon CE,Weaver W,Blahut REetal. The mathematical theory of communication. Urbana: University of Illinois Press, 1949.
[14]van den Wollenberg AL. Redundancy analysis an alternative for canonical correlation analysis.Psychometrika, 1977, 42(2):207-219.
[15]Muller KE. Relationships between redundancy analysis, canonical correlation, and multivariate regression.Psychometrika, 1981, 46(2):139-142.
[16]宗志祥,许崇任,任久长等.洋河水库轮虫群落用于水质评价的研究.环境科学学报,1993,13(1):102-106.
[17]都雪,王齐东,张超文.洪泽湖轮虫群落结构及其与环境因子之间的关系.湖泊科学,2014,26(2):269-276.
[18]饶小珍,许友勤,陈寅山.福州内河的轮虫与水质污染评价.福建师范大学学报:自然科学版,2000,16(1):71-75.
[19]高原,李新辉,赖子尼等.珠江三角洲河网浮游轮虫的群落结构.应用生态学报,2014,25(7):1-12.
[20]王庆,刘丽君,杨宇峰.南方某水库轮虫群落特征及其对环境因子的响应.生态学报,2010,20(13):3385-3395.
[21]Gannon JE, Stemberger RS. Zooplankton(especially crustaceans and rotifers) as indicators of water quality.TransactionsoftheAmericanMicroscopicalSociety, 1978, 97(1):16-35.
[22]侯磊,王庆,杨宇峰.珠江广州河段轮虫群落结构特征与水质生态学评价.暨南大学学报:自然科学版,2011,32(3):312-317.
[23]钱方平,席贻龙,温新利等.湖泊富营养化对轮虫群落结构及物种多样性的影响.生物多样性,2007,15(4):344-355.
[24]邓道贵,金显文,葛茜等.淮北采煤塌陷区小型湖泊轮虫群落结构的季节变化.湖泊科学,2012,24(1):111-116.
[25]饶利华,吴芝瑛,徐骏等.杭州西湖的轮虫群落结构及与水体环境因子的关系.湖泊科学,2013,25(1):138-146.
[26]张根,席贻龙,薛颖昊.基于rDNA ITS序列探讨粉煤灰污染对萼花臂尾轮虫种群遗传多样性的影响.生物多样性,2010,18(3):241-250.
[27]谷孝鸿,刘桂英.滤食性鲢鳙鱼对池塘浮游生物的影响.农村生态环境,1996,12(1):6-10,41.
[28]陈光荣,雷泽湘,谭镇等.环境因子对广东城市湖泊后生浮游动物的影响.水生态学杂志,2010,3(4):28-32.
[29]郭凯,赵文,殷守仁等.北京轮虫群落结构与水体富营养化状况.湖泊科学,2010,22(2):256-264.
[30]邱小琮,赵红雪,孙晓雪.鸣翠湖轮虫群落特征及其与环境因子的关系.宁夏大学学报:自然科学版,2012,33(1):62-68.
J.LakeSci.(湖泊科学), 2015, 27(4): 657-666
©2015 byJournalofLakeSciences
Rotifer communities in the small lakes around Huainan coal mine subsidence areas
WANG Xiaomeng1, YI Qitao1,2, XU Xin1, LI Hui1& ZHANG Lei1
(1:SchoolofEarthandEnvironment,AnhuiUniversityofScienceandTechnology,Huainan232001,P.R.China)
(2:ResearchCenterforEco-environmentalSciences,ChineseAcademyofSciences,Beijing100085,P.R.China)
Abstract:Three small lakes at PXPJ site, PXGQ site and PXXQ site in the Huainan Panxie coal mine subsidence areas were chosen as research sites, which were characterized by different conditions in hydrology, ecological and environmental factors. The investigation of rotifer communities and aquatic environmental factors were conducted in the four seasons of 2013-2014 to address their relationship. A total of 32 species have been recorded, which belong to 15 genera. 25 species of the 13 genera at PXPJ site, 24 species of the 13 genera at PXGQ site, and 20 species of the 11genera at PXXQ site have been indentified. Keratella valga, Trichocerca rousseleti, Asplanchna priodonta, Asplanchna brightwelli, Polyarthra trigla Ehrenberg, Gosse were the dominate species in the lakes. Rotifer abundance were 500-2800ind./L at PXPJ site, 950-3350ind./L at PXGQ site, and 400-3900ind./L at PXXQ site. The average Shannon-Wiener diversity indexes for rotifer communities within the three studied sites were 2.71, 2.68 and 2.54 respectively, indicating β-type of medium pollution water quality in the aquatic zones.Redundancy analysis based on rotifer community and aquatic environmental factors showed that water temperature, conductivity and nutrient in the lakes are the main factors affecting the rotifer communities in those lakes.
Keywords:Coal mine subsidence areas; subsidence lakes; rotifer; aquatic environment factors
通信作者**;E-mail:yiqitao@163.com.
DOI10.18307/2015.0414
