槲寄生与4种寄主叶表皮微形态特征及生态适应性
2015-02-24李霞徐增奇任皓蔡可心谢晓东王长宝
李霞,徐增奇,任皓,蔡可心,谢晓东,王长宝*
(1.佳木斯大学生命科学学院,黑龙江佳木斯 154007;2.杜尔伯特伊利乳业有限责任公司,黑龙江杜尔伯特 166200)
槲寄生(ViscumcoloratumNakai)是桑寄生科(Loranthaceae)半寄生常绿小灌木,其根部特化为寄生根并直接与寄主木质部导管相连,从宿主身上获得水和无机盐。主要分布在东北、华北、朝鲜、日本、苏联[1]。国内外学者对槲寄生的研究多集中在化学成分、生物活性及药理作用等方面,但有关槲寄生与寄主植物之间生态关系的研究鲜有报道[2-3]。本文通过比较槲寄生与4种寄主叶表皮微形态特征,探讨两者间的相关性和差异性,进而解释它们的生态适应性。
1 材料与方法
1.1 实验材料
实验所用寄主植物分别为山杨(Populusdavidiana)、白桦(Betulaplatyphylla)、黑桦(Betula dahurica)、蒙古栎(Quercusmongolica)。实验材料全部采自黑龙江省桦川县老平岗林场(130°30′35″~130°43′03″E,46°31′37″~46°37′59″N),并在FAA固定溶液中保存。
1.2 实验方法
将固定液中的叶片取出,切成1cm2的小块,放入50%NaCl O溶液中浸泡12h后用蒸馏水冲洗干净,撕取上、下叶表皮制成水封片。在Olympus 1X51显微镜下分别观察上、下表皮细胞形态、气孔器类型、垂周壁式样,并拍照。同时测定每种植物叶片的气孔密度SD、气孔指数SI及气孔长度SL。SD是随机观测20个视野后取平均值;SI=[S/(S+E)]×100,其中,S为单位视野内气孔数目,E为单位视野内表皮细胞数目;SL即两个保卫细胞连接点间的距离,为20个视野中随机测量5个气孔长度后取平均值。采用SPSS19.0软件对数据进行处理。本文所用术语参考陈机[4]所述。
2 结果与分析
2.1 槲寄生与寄主叶表皮细胞特征比较
槲寄生与黑桦的上、下叶表皮细胞形状均为多边形,垂周壁式样同样均为弓形。蒙古栎上叶表皮细胞形状为多边形,而下表皮却为无规则形,垂周壁式样也不相同:上表皮为弓形,下表皮为波状。山杨上、下叶表皮均为无规则形,垂周壁式样均为波形。
在叶表皮细胞体积大小方面,槲寄生的上、下叶表皮细胞体积大小与4种寄主植物存在明显差异,槲寄生上、下叶表皮细胞大于所有寄主植物叶表皮细胞(表1)。

表1 槲寄生与4种寄主植物叶表皮细胞特征
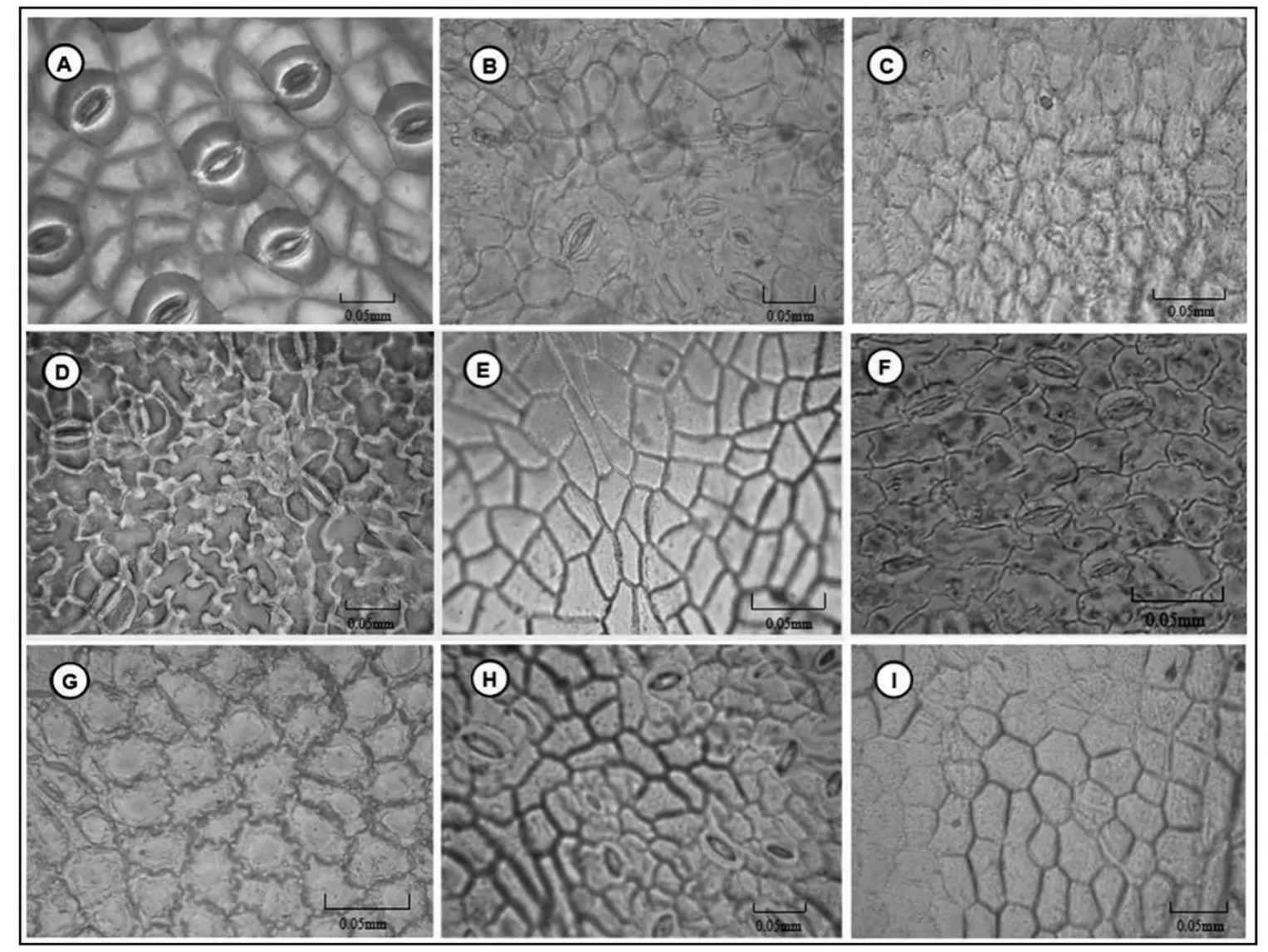
图1 槲寄生与4种寄主的上、下叶表皮
2.2 槲寄生与寄主叶表皮气孔特征比较
槲寄生叶片为等面叶,上、下叶表皮差异不明显(图1 A);4种寄主植物的叶片为异面叶,上、下叶表皮存在较大差异,上表皮没有或极少分布气孔(图1 B-I)。在气孔密度方面,槲寄生与4种寄主植物差异显著:槲寄生上表皮的气孔密度为16.6个/mm2,下表皮为16.3个/mm2;山杨叶片下表皮气孔密度最大达到83.8个/mm2;白桦次之,为37.8个/mm2;黑桦34.6个/mm2;蒙古栎气孔密度最小,是24.1个/mm2(表2)。在气孔指数方面,槲寄生与4种寄主植物的存在一定差异。在这5种植物中,仅山杨叶下表皮的气孔指数较大达到24.4。白桦下表皮的气孔指数为18.4,蒙古栎为16.4,槲寄生为14.2,黑桦最小为13.0(表2)。
在气孔器类型方面,槲寄生与4种寄主植物的气孔器类型都为无规则型(表2)。
从图2中可以看出,在气孔长度方面,槲寄生与它的寄主植物具有较大的差异,槲寄生叶下表皮的气孔长度远大于4种植物达到71.3μm,并且气孔长度大小较为稳定。在寄主植物中蒙古栎的气孔长度略大为41.8μm,但气孔长度大小较为不稳定。黑桦的气孔长度略小些为32.7μm,同样气孔长度大小较为不稳定。白桦的气孔长度更小为28.4μm,气孔长度大小较为稳定。山杨的气孔长度最小为24.5μm,气孔长度大小相对稳定。

表2 槲寄生与4种寄主植物叶表皮气孔特征
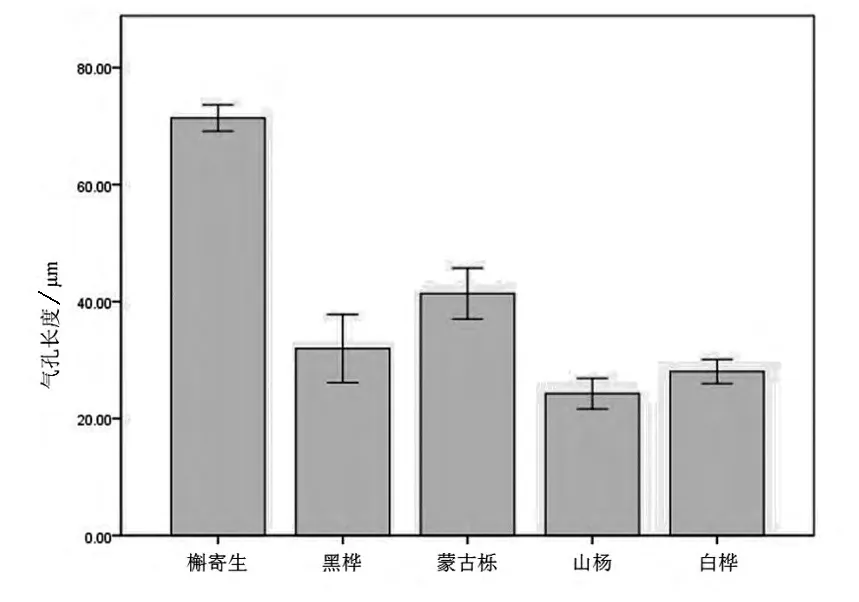
图2 槲寄生与4种寄主叶下表皮气孔长度的比较
3 讨论
植物体是一个开放的体系,叶片是在地面环境中暴露最多的器官,因此,植物对环境的适应性通常较多的反映在叶的形态结构上[5]。槲寄生是一种半寄生植物,在生理生化方面与其寄主植物必然存在较大的联系,槲寄生的叶表皮与寄主植物的叶表皮应具有很大相关性。
众多研究结果表明,小而数目众多的气孔、较小的细胞体积被认为是植物抵御不良环境的影响[6-8]。气孔数目多反映了较高的光合速率,同时在控制蒸腾作用方面具有两重性:水分充足时气孔打开,促进蒸腾作用和防止过热灼烧;水分紧张时气孔关闭,抑制蒸腾作用[9]。虽然气孔密度小于4种寄主,由于槲寄生的叶片为等面叶,上、下叶表皮的相关特征均相同,在叶面积上槲寄生的气孔数量与4种寄主植物应相差不多。此外,槲寄生叶表皮细胞体积大于寄主植物,因此在叶表皮形态特征方面符合寄生植物的生理生态特征,即蒸腾速率高于寄主植物、光合速率低于寄主植物[10-11]。4种寄主植物气孔密度从大到小的排列顺序为:山杨>白桦>黑桦>蒙古栎,在气孔长度方面,4种寄主植物从小到大的排列顺序同样为:山杨<白桦<黑桦<蒙古栎,这两组数据均与槲寄生在不同寄主植物上的数量分布规律相一致[12]。气孔密度越大,气孔长度越小,反映出植物的光合速率和蒸腾速率越高,这可能更利于槲寄生养分和水分的获取。因此,在一定范围内,叶表皮气孔密度越大并且气孔长度越小的寄主植物更利于槲寄生的繁殖与生长。
[1]中国科学院植物志编辑委员会.槲寄生属,中国植物志.24卷[M].北京:科学出版社,1988:148-149.
[2]孔德云,罗思齐,李惠庭,等.槲寄生化学成分的研究Ⅰ[J].医药工业,1987,18(3):123-127.
[3]王庆端,刘梅筠,王东阳,等.槲寄生总生物碱的抗肿作用[J].中国中药杂志,1994,19(1):45-47.
[4]陈机.植物发育解剖学(上册)[M].济南:山东大学出版社,1992.
[5]张泓,陈丽春,胡正海.骆驼蓬营养器官的旱生结构[J].植物生态学报与地植物学报,1992,16(3):181-185.
[6]梁松洁,张金政,张启翔,等.北方地区藤本类忍冬叶表皮结构及其生态适应性比较研究[J].植物研究,2004,24(4):435-440.
[7]萨仁,苏德毕力格,陈家瑞.黄华属植物叶表皮特征及其生物学意义[J].草地学报,2000,8(1):65-76.
[8]李强,曹建华.黄荆叶表皮形态结构及其生态适应性研究[J].河南师范大学学报,2008,36(5):131-134.
[9]贺金生,陈伟烈,王勋陵.高山栎叶的形态结构及其生态环境的关系[J].植物生态学报,1994,18(3):219-227.
[10]Glatzel G.Mineral nutrition and water relations of hemiparasitic mistletoes:a question of portioning.Experiments with Loranthus europaeus on Quercus petraea and Quercus robur[J].Oecologia,1983,56:193-201.
[11]黄丽.槲寄生与寄主红树之间水分和营养研究[D].厦门:厦门大学,2009.
[12]王长宝,徐增奇,岳仁杰.完达山区槲寄生的种群特征研究[J].植物科学学报,2013(4):345-352.
