Leuprolide acetate induces structural and functional recovery of injured spinal cord in rats
2015-02-07CarmenazGalindoBeatrizmezGonzlezEvaSalinasDenisseCalderVallejoIrmaHernndezJassoEduardoBautistaLuisQuintanar
Carmen Díaz Galindo, Beatriz Gómez-González, Eva Salinas, Denisse Calderón-Vallejo, Irma Hernández-Jasso, Eduardo Bautista, J Luis Quintanar,
1 Laboratory of Neurophysiology, Department of Physiology and Pharmacology, Universidad Autónoma de Aguascalientes, Aguascalientes, México
2 Area of Neuroscience, Department of Reproductive Biology, CBS, Universidad Autónoma Metropolitana, Unidad Iztapalapa, México
3 Laboratory of Immunology, Department of Microbiology, Centro de Ciencias Básicas, Universidad Autónoma de Aguascalientes, Aguascalientes, México
Leuprolide acetate induces structural and functional recovery of injured spinal cord in rats
Carmen Díaz Galindo1, Beatriz Gómez-González2, Eva Salinas3, Denisse Calderón-Vallejo1, Irma Hernández-Jasso1, Eduardo Bautista1, J Luis Quintanar1,*
1 Laboratory of Neurophysiology, Department of Physiology and Pharmacology, Universidad Autónoma de Aguascalientes, Aguascalientes, México
2 Area of Neuroscience, Department of Reproductive Biology, CBS, Universidad Autónoma Metropolitana, Unidad Iztapalapa, México
3 Laboratory of Immunology, Department of Microbiology, Centro de Ciencias Básicas, Universidad Autónoma de Aguascalientes, Aguascalientes, México
Gonadotropin-releasing hormone (GnRH) and its synthetic analog leuprolide acetate, a GnRH agonist, have neurotrophic properties. This study was designed to determine whether administration of leuprolide acetate can improve locomotor behavior, gait, micturition ref ex, spinal cord morphology and the amount of microglia in the lesion epicenter after spinal cord injury in rats. Rats with spinal cord compression injury were administered leuprolide acetate or saline solution for 5 weeks. At the 5thweek, leuprolide acetate-treated rats showed locomotor activity recovery by 38%, had improvement in kinematic gait and exhibited voiding ref ex recovery by 60%, as compared with the 1stweek. By contrast, saline solution-treated rats showed locomotor activity recovery only by 7%, but voiding ref ex did not recover. More importantly, leuprolide acetate treatment reduced microglial immunological reaction and induced a trend towards greater area of white and gray matter in the spinal cord. Therefore, leuprolide acetate has great potential to repair spinal cord injury.
nerve regeneration; spinal cord injury; leuprolide acetate; gonadotropin-releasing hormone; neurotrophic factor; microglia; micturition ref ex; gait; inf ammation; neural regeneration
Funding: This study was supported by a grant from CONACyT for scholarship 376921/246887. Díaz Galindo C, Gómez-González B, Salinas E, Calderón-Vallejo D, Hernández-Jasso I, Bautista E, Quintanar JL (2015) Leuprolide acetate induces structural and functional recovery of injured spinal cord in rats. Neural Regen Res 10(11):1819-1824.
Introduction
Spinal cord injury (SCI) causes dif erent and long-term neurological disorders and even morbidity in humans. There have been no ef ective interventions for any of the main types of neurological disorders after SCI, including motor and sensory def cits, bladder, bowel and sexual dysfunction, chronic pain, and autonomic dysref exia. SCI results in acute as well as progressive secondary tissue damage, and initiates a number of regenerative and neuroprotective responses within the damaged nervous system. Possible therapies are directed to generate neuroprotection, regeneration or an enhancement in plasticity of the uninjured tissues (Hagg and Oudega, 2006).
There are dif erent experimental approaches to counteracting the ef ects of SCI, such as the use of neurotrophic factors, Nogo neutralizing antibodies and N-metyl-D-aspartate receptor modulators (Schwab, 2004; Thuret et al., 2006). Neurohormone gonadotropin-releasing hormone (GnRH) has been recently shown to be neurotrophic. In vitro, GnRH administration induced changes in outgrowth, number and length of neurites in rat cortical neurons (Quintanar and Salinas, 2008). The receptor for this decapeptide has been identified in cerebral cortex neurons, spinal cord, bladder and other extra-pituitary tissues (Bahk et al., 2007; Quintanar et al., 2007, 2009). GnRH treatment improved locomotor activity and bladder function, and increased the expression of neurof laments in spinal cords in rats with SCI (Calderon-Vallejo and Quintanar, 2012).
Leuprolide acetate (LA) is a synthetic agonist analog of GnRH. It is less susceptible to proteolysis and has a greater binding affinity to GnRH receptors than the natural hormone, increasing its biological activity (Periti et al., 2002). LA is used for disorders of the reproductive system such as prostate cancer. However, LA treatment has been shown to decrease the severity of clinical signs related to locomotion of rats with experimental autoimmune encephalomyelitis. It also induces a signif cant body weight gain together with increases in neurof lament and myelin basic protein expression and axonal diameter in the spinal cord (Guzmán-Soto et al., 2012).
One of the processes that occur in the injured spinal cord area is activation of the immune response. SCI leads to an early inf ltration of macrophages/microglia. The initial inf ammatory response is followed by an active phase ofresolution which is necessary to repair the injury. Inf ammatory response has undesirable ef ects and produces a greater neurological damage and neurodegeneration (Raposo and Schwartz, 2014).
The aim of this study was to determine whether administration of LA in rats with SCI can improve locomotion behavior, kinematic gait and maturation, promote the recovery of morphology of injured spinal cord, and increase the number of microglial cells.
Materials and Methods
Animals
Female Wistar rats, aged 8 months, weighing 250–330 g, from Universidad Autónoma de Aguascalientes, Mexico were included in this study. They were treated according to Institutional Welfare Regulations of The University Autonomous of Aguascalientes. This study was approved by Ethics Committee of Laboratory Animals Care, Universidad Autónoma de Aguascalientes, Mexico. All ef orts were made to minimize animal discomfort and reduce the number of animals used. Animals were kept under 12-hour light/dark cycle and at controlled temperature. Purina chow and tap water were provided ad libitum.
Surgery
One month before SCI, both ovaries of rats were surgically removed through a dorsal incision under deep anesthesia with methyl ether, in order to avoid the ef ects of ovarian hormones. Ovariectomized animals were randomly divided into three groups: Sham SCI (sham; n = 9), SCI treated with physiological saline solution (SS; n = 11) and SCI treated with LA (LA; n = 10). Rat models of spinal cord compression injury were established according to a previously described method (Vanicky et al., 2001). A 2-French Fogarty catheter was inserted at T12level and then advanced 1 cm cranially to the site where laminectomy was performed. The catheter balloon was inf ated with 20 μL of 0.9% sodium chloride for 5 minutes and then def ated and removed. In the sham group, the catheter was introduced without inf ating the balloon. From the day of surgery, manual bladder emptying was performed at least twice a day until ref ex bladder control was re-established. All animals were injected with penicillin (Penprocilin; 5,000 IU, i.m.) once a day for 7 days and Neomelubrin (15 mg/kg; i.m.) once daily for 3 days. All animals were sacrif ced 5 weeks later.
LA administration
Rats were administered LA (Sigma, St. Louis, MO, USA; 10 μg/kg, i.m.) once a day after SCI for 3 consecutive days. Thereafter, only one injection was given at the dose the same as that used in our previous experiments (Guzmán-Soto et al., 2012) every 4 days for 5 weeks. Rats in the SS group were identically given physiological saline solution (0.9% sodium chloride).
Locomotor behavior and kinematic gait evaluation
Locomotor activity evaluation was performed using the Basso, Beattie, and Bresnahan (BBB) locomotor rating scale (Basso et al., 1995), beginning 1 day after SCI and later once a week. Six evaluations were performed starting from the 1stday after SCI (week 0). Two independent examiners blinded to therapy rated movement capacities during the tests. On the same days, kinematic gait analysis was also performed. To visualize iliac crest, greater trochanter, knee, external malleolus and f fth metatarsal correctly, rats were marked on these f ve points with a permanent marker in the left leg. A clear rectangular rigid structure delimited the path of the animals. The walking activity was videotaped. The video was divided according to the principle of 15 pictures per second and the abovementioned f ve points were identif ed on every picture. Using MacBiophotonics Image J software (National Institute of Health, MD, USA), these points were transformed into coordinates, which were interpreted by the software developed in our laboratory to establish the step cycle duration and the stance and swing phases. To obtain values for all the variables plotted for step cycle duration, swing, and stance phase, the average of three steps performed for each rat was calculated. In rats that had a complete step cycle, the distance development between each stance phase plotted as stride distance was measured (cm) and the stride speed (cm/sec) was calculated using the stride distance and the swing phase duration. In some rats, SCI caused locomotor inability that restricted them to develop the path. The maximum waiting time to carry out the path was 10 seconds. The rats that did not move were retired, and the step cycle duration was rated as 10 seconds.
Micturition ref ex
Data on micturition was obtained from a daily record about the presence or absence of distended bladder and the requirement of manual bladder empty. It was expressed as the average number of days that rats from each group required assistance.
Histology of white and gray matter and microglial cells
Five weeks after SCI, all rats were sacrificed by exposing them to a deep anesthesia with methyl ether. To achieve a positive pressure inside the left ventricle, physiological saline solution was f lled through the use of a syringe. The spinal cords were removed, and 1 cm2of epicenter of the lesioned area was obtained. The harvested tissue was f xed with 10% neutral formalin and then sliced into 5 μm-thick sections on a microtome for morphological and immunohistochemical analysis. For morphological analysis, quantification of white and gray matter of spinal cord was performed using Klüver-Barrera staining (Prophet et al., 1994). Five rats from each group were used for light microscopy examination (Zeiss, Axiostar plus, Carl Zeiss Microscopy, Thornwood, NY, USA) at 4× magnif cation. The area (mm2) was determined using SigmaScan Pro software (version 4.01, Cranes Software, 512093 Bangalore, Karnataka, India). For microglial cells analysis, immunohistochemistry was performed. Slices were incubated with primary polyclonal antibody against ionized calcium binding adapter molecule 1 (IBA-1; goat polyclonal to IBA-1; Abcam, USA, Ab48004) (marker to microglia)1:500, in a wet chamber at 4°C overnight. Labeling was made according to the kit instruction (Dako Cytomatio LSAB + System HRP, K0690 Kit, USA). Color was developed by reaction with 3,3′-diaminobenzidine (Sigma, USA). The nuclei were counterstained with hematoxylin. The immunoreactivity was determined using AxioVision40V4.8.2.0. (Carl Zeiss Microimaging, Inc., Thornwood, NY, USA). Microglial cells were quantif ed as density of immunoreactive cells per 100 μm2of tissue. The average of microglial cells across three spinal cords of each group was calculated. All microglial cells in one cross-section were evaluated. At least 10 microphotos were taken from each section at 40× magnif cation for histological observation.
Statistical analysis
One-way analysis of variance (ANOVA) with Dunnett’s post hoc test was performed using GraphPad Prism version 5.00 for Windows (GraphPad Software, San Diego, CA, USA). Data are expressed as the mean ± SEM. Statistical signif -cance of locomotor behavior and gait analysis was evaluated at each week during the evaluation period. For microglia, white and gray matter, and micturition ref ex, analysis was made at the end of the evaluation period (5thweek). A level of P < 0.05 was considered statistically signif cant.
Results
Locomotor behavior
The rats that showed hind limb paralysis after SCI were included in this study. Initially, there was no signif cant dif erence in BBB scores between SS and LA groups. Rats in these two groups exhibited similar locomotion characteristics, with a score close to 0 (Figure 1). However, rats in the sham group showed slightly reduced locomotor activity followed by an immediate and progressive recovery close to level 21 corresponding to normal locomotor activity (Figure 1). A more signif cant recovery of locomotor activity was observed in the LA group than in the SS group from the 2ndweek to the 5thweek. During this time period, the locomotor activity in the LA group improved by 38% of that in the 1st week. At the same time, spontaneous recovery was enhanced by 7% in the SS group. Although recovery in the LA group was signif -cantly greater than that in the SS group, it did not reach the level of recovery in the sham group (Figure 1).
Kinematic gait
Gait analysis was divided into three sections: step cycle duration, stance phase duration, and swing phase duration. Step cycle duration in the LA group decreased from 9 to 4.6 seconds (initial and f nal evaluations) and it was signif cantly dif erent compared with the SS group in the 2nd, 3rdand 5thweeks (Figure 2A).
In the LA group, a reduction in stance phase duration was observed from 8.8 seconds at initial evaluation to 4.1 seconds at f nal evaluation. Swing phase duration also diminished in the LA group along weeks, from 8.3 to 3.5 seconds. In the SS group, reduction in stance phase duration was signif cant at the 1st, 2nd, 3rdand 5thweeks, and as for swing phase duration, signif cant reduction lasted from the 1st week until the 5thweek (Figure 2B and C).
The distance traveled in the LA group was greater in f nal evaluation than in initial evaluation (6.1 to 1.9 cm respectively) and it was significantly higher than that in the SS group at the 1st, 2ndand 3rdweeks (Figure 2D). The stride speed of rats in the LA group increased from 1.3 cm/sec as initial rate to 9.0 cm/sec in f nal evaluation. The increase of stride speed in the LA group was signif cantly greater than that in the SS group at the 1st, 2ndand 3rdweeks (Figure 2E). In the sham group, there were no signif cant dif erences in these variables even at the beginning of evaluation, and locomotor activity was slightly altered with time.
Micturition control
According to the data daily collected during the experiment, only four out of ten rats in the LA group needed manual bladder emptying at the end of the experiment. In the SS group, all rats required assistance. The number of the days in which rats in each group needed assistance before retrieval of urination ref ex in the LA group was signif cantly less than in the SS group (19 vs. 35 days)( Figure 3).
Morphology of injured spinal cord
In the LA group, the conf guration of white and gray matter of spinal cord was more similar to that in the sham group and it was better than that in the SS group. After LA administration, a trend towards a slight, but not signif cant, increase in the area of white and gray matter was observed (Figure 4B). Spinal cord sections from the SS group showed many large cavitations without nervous tissue, trabecular structures or cellular infiltrates, and in each cavitation, a small number of motorneurons with poor morphology were observed. However, similar to the sham group, spinal cord sections in the LA group showed fewer cavitations in which a greater number of neurons with better morphology were observed than in the SS group (Figure 4A).
Microglial immunoreactivity
In order to analyze the amount of microglia present in the epicenter of the lesion, immunohistochemistry was performed and the density of microglial cells was measured. Results showed that the area of microglia in the epicenter lesion in the LA group was signif cantly minimized than that in the SS group (1,080 ± 270 vs. 2,842 ± 434 μm2), but it was similar to that in the sham group (1,421 ± 101 μm2) (Figure 5).
Discussion
The most common consequence of SCI is the paralysis due to neural circuit interruptions. Although there is some degree of spontaneous recovery, lost connections are hardly restored. It is an option to consider the use of neurotrophic factors to induce nerve regeneration. Neurotrophic factors have a key role in the modulation of neuronal survival, neurite outgrowth, synaptic plasticity and neurotransmission in both intact and injured nervous system.
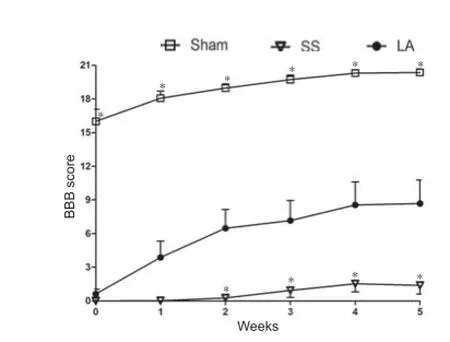
Figure 1 Locomotor ability of rats with spinal cord injury after LA treatment.
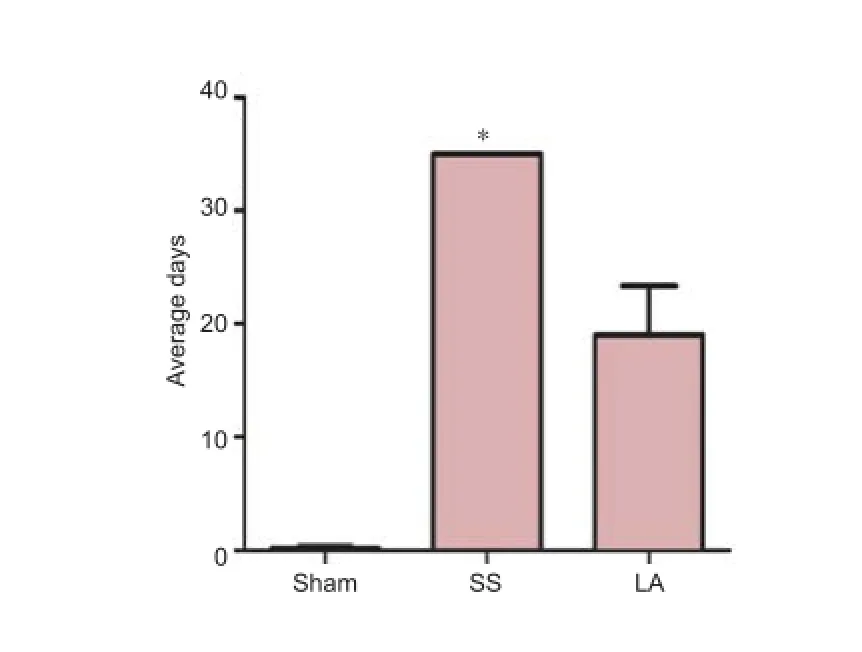
Figure 3 The micturition in rats with spinal cord injury after LA treatment.
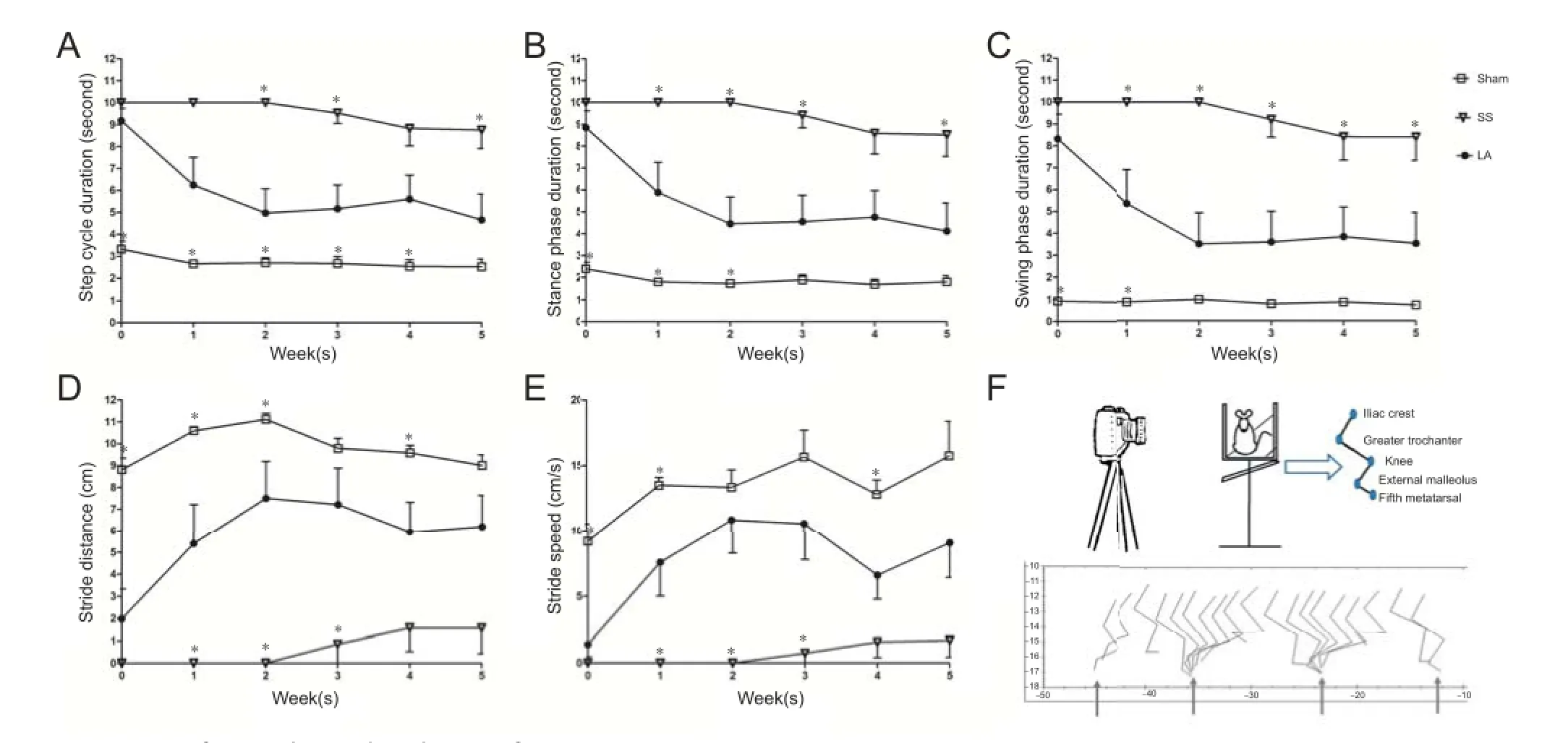
Figure 2 Gait of rats with spinal cord injury after LA treatment.
In this study, we found that LA, a GnRH agonist, significantly regained locomotor activity of SCI rats. Calderon-Vallejo and Quintanar (2012) reported that GnRH treatment recovered locomotor activity with similar f ndings to ours. Both results are strongly tied, because these ef ects are probably mediated through activation of GnRH receptors, which have been described in the spinal cord motoneurons (Dolan et al., 2003; Quintanar et al., 2007). In an experimental autoimmune encephalomyelitis model, LA administration produces an increase in the axonal growth and in neurof laments and myelin basic protein expression (Guzmán-Soto et al., 2012), which could explain the improvement in locomotor activity found in our experiments. The degree of locomotor activity recovery in rats treated with LA was higher than in those treated with GnRH (Calderon-Vallejo and Quintanar, 2012), which suggests that LA has greater potential for clinical application than GnRH.
In kinematic analysis, we observed that LA treatment in SCI rats decreased step cycle, stance phase, and swing phase durations, but it resulted in increases in the stride distance and speed. LA rats spent less time in each step phase, traveled greater distance, and performed faster steps. These results can be considered as a sign of locomotor activity recovery, which were not observed in rats with physiological salinesolution treatment. These results are similar to the outcomes by Hamers et al. (2001) who studied gait parameters in two dif erent SCI types, i.e., transection of the dorsal half of the spinal cord and spinal cord contusion.

Figure 4 White and gray matter area in spinal cord of rats with spinal cord injury after LA treatment.
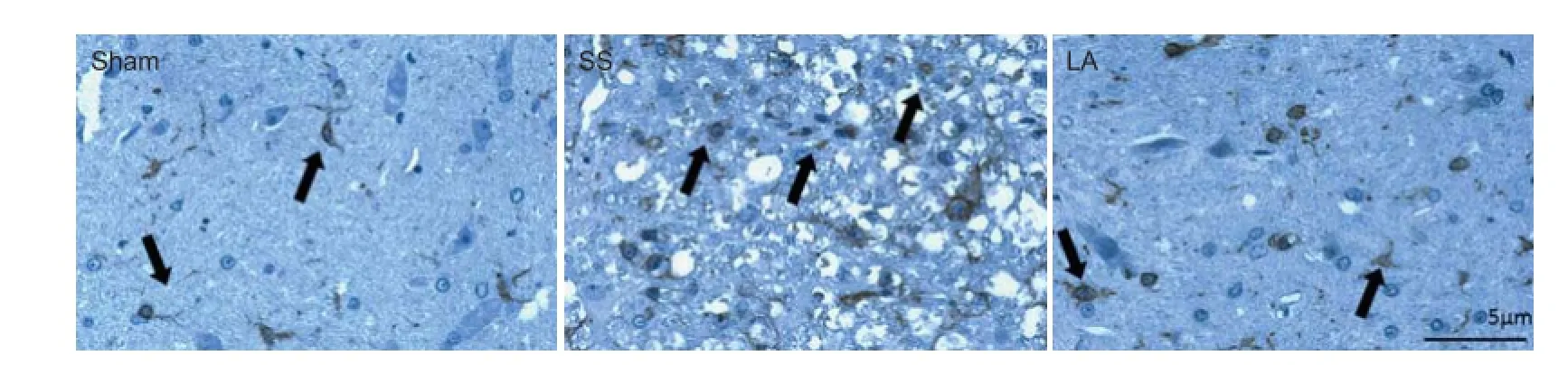
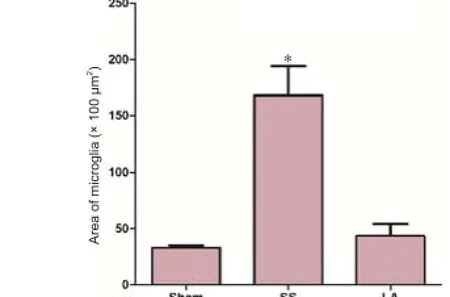
Figure 5 Density of microglial cells in spinal cord of rats with spinal cord injury after LA treatment.
The locomotor activity recovery in LA-treated animals is due to dif erent mechanisms including remyelination (Guzman-Soto et al., 2012) or increases in microf lament protein expression, neurite outgrowth (Calderon-Vallejo and Quintanar, 2012), and axonal diameter (Quintanar et al., 2011). These results can be explained by reorganization of local networks via propriospinal circuitry. These local networks control the involuntary movements, which do not require cortico- or rubrospinal tracts, proposed as an explanation for the ability of spinal cord injured animals to produce walking movements (Ek et al., 2010). However, supraspinal tracts were likely involved in the control of voluntary movements observed in animals treated with LA. It could be related to reconnections going through injured area originated from supraspinal neurons.
Propiospinal circuits provide an explanation for the restorations of micturition ref ex in LA animals, in which the number of days that they need assistance of manual bladder emptying was reduced. This is possible due to the reorganization in ref ex pathways inside the spinal cord, as reported by Groat and Yoshimura (2012).
In this study, LA administration modif ed the gray and white matter areas, decreasing scar area and promoting thte recovery of spared tissue in SCI rats. High BBB scores were positively correlated with spared tissue in the spinal cord lesion (Basso et al., 1995). Structural conformation of thespinal cord is kept by both suf cient numbers of neurons and axons including their myelin. In traumatized spinal cord, only remnants of both gray and white matter exist due to the presence of hypercellularity, inflammation, cavitation, increased extracellular space and a loose f brous matrix (Totoiu and Keirstead, 2005). Our results showed that in the LA group, spared tissue was increased, BBB scores were higher, faster stride speed and longer stride distance were observed in the LA group than in the sham group. These results ref ect the new integration of spared descending and af erent-driven signals, as recovery after contusive SCI has been reported to be identified by changes in gait biomechanics and muscle activation patterns (Hansen et al., 2012).
In the LA group, signif cantly higher BBB score and improved gait were observed at the 1stand 2ndweeks during the evaluation period compared to the sham group. As a consequence of SCI, microglia considerably increased independently of spared tissue. An early infiltration of macrophages/microglia in traumatic lesions of spinal cords has been observed in a rat model of SCI. The immune cell cascade involves inf ltration of neutrophils and activation of resident microglia, followed by subsequent accumulation of monocyte-derived macrophages and the later entry of lymphocytes into the lesion site. The monocytes that inf ltrate the injured area acquire a pro-inf ammatory/classical prof le (Raposo and Schartz, 2014). The reduction in the density of microglial cells observed in the LA group is probably associated with the locomotor activity in these rats.
In conclusion, administration of LA partially improves lotomotor activity, gait, micturition ref ex, spinal cord morphology and decreases microglial area in a rat model of SCI. Promotion of neuronal survival by administration of neurotrophic factors and a possible immunomodulation to counteract secondary injury are a promising approach to repair of SCI. Additionally, even LA administered via intramuscular injection is able to cross the blood-spinal cord barrier as observed in previous studies on patients with prostate cancer. LA is a potential alternative treatment of SCI because of its safety and ease in use as well as few side ef ects.
Acknowledgments: We would like to express our sincere gratitude to Kalman Kovacs and Fabio Rotondo for paper review and Dra. Irene Guzmán Soto, Crhistian Jafet Pérez Ferrer, Biól. Araceli Adabache Ortíz, and Violeta Saraí Jiménez Hernández for methodological support.
Author contributions: CDG, BGG, ES, DCV and JLQ conceived and design this study, were responsible for def nition of intellectual content. CDG, DCV, IHJ and EB performed experiments. CDG, DCV, ES and JLQ wrote, revised, and reviewed the paper. IHJ and EB performed statistical analysis and retrieved the literature. All authors participated in data acquisition and data analysis and approved the f nal version of this paper. Conf icts of interest: None declared.
Plagiarism check: This paper was screened twice using Cross-Check to verify originality before publication.
Peer review: This paper was double-blinded, stringently reviewed by international expert reviewers.
Bahk JY, Kim MO, Park MS, Lee HY, Lee JH, Chung BC, Min SK (2008) Gonadotropin-releasing hormone (GnRH) and GnRH receptor in bladder cancer epithelia and GnRH ef ect on bladder cancer cell proliferation. Urol Int 80:431-438.
Basso DM, Beattie MS, Bresnahan JC (1995) A sensitive and reliable locomotor rating scale for open f eld testing in rats. J Neurotrauma 12:1-21.
Calderón-Vallejo D, Quintanar JL (2012) Gonadotropin-releasing hormone treatment improves locomotor activity, urinary function and neurof lament protein expression after spinal cord injury in ovariectomized rats. Neurosci Lett 515:187-190.
Dolan S, Evans NP, Richter TA, Nolan AM (2003) Expression of gonadotropin-releasing hormone and gonadotropin-releasing hormone receptor in sheep spinal cord. Neurosci Lett 346:120-122.
Ek CJ, Habgood MD, Callaway JK, Dennis R, Dziegielewska KM, Johansson PA, Potter A, Wheaton B, Saunders NR (2010) Spatio-temporal progression of grey and white matter. Damage following contusion injury in rat spinal cord. PLoS One 5:e12021.
Groat WC, Yoshimura N (2012) Plasticity in reflex pathways to the lower urinary tract following spinal cord injury. Exp Neurol 235:123-132.
Guzmán-Soto I, Salinas E, Hernández-Jasso I, Quintanar JL (2012) Leuprolide acetate, a GnRH agonist, improves experimental autoimmune encephalomyelitis: a possible therapy for multiple sclerosis. Neurochem Res 37:2190-2197.
Hagg T, Oudega M (2006) Degenerative and spontaneous regenerative processes after spinal cord injury. J Neurotrauma 23:264-280.
Hamers FP, Lankhorst AJ, Laar TJ, Veldhuis WB, Gispen WH (2001) Automated quantitative gait analysis during overground locomotion in the rat: its application to spinal cord contusion and transection injuries. J Neurotrauma 18:187-201.
Hansen CN, Linklater W, Santiago R, Fisher LC, Moran S, Buford JA, Basso M (2012) Characterization of recovered walking patterns and motor control after contusive spinal cord injury in rats. Brain Behav 2:541-552.
Periti P, Mazzei T, Mini E (2002) Clinical Pharmacokinetics of depot leuprorelin. Clin Pharmacokinet 41:485-504.
Prophet EB, Mills B, Arrington JB, Sobin LH (1994) Laboratory methods in histotechnology. Armed forces institute of pathology. Washington DC. USA.
Quintanar JL, Salinas E, González R (2007) Expression of gonadotropin-releasing hormone receptor in cerebral cortical neurons of embryos and adult rats. Neurosci Lett 411:22-25.
Quintanar JL, Salinas E (2008) Neurotrophic ef ects of GnRH on neurite outgrowth and neurof lament protein expression in cultured cerebral cortical neurons of rat embryos. Neurochem Res 33:1051-1056.
Quintanar JL, Salinas E, González R (2009) Gonadotropin-releasing hormone receptor in spinal cord neurons of embryos and adult rats. Neurosci Lett 461:21-24.
Quintanar JL, Salinas E, Quintanar-Stephano A (2011) Gonadotropin-releasing hormone reduces the severity of experimental autoimmune encephalomyelitis, a model of multiple sclerosis. Neuropeptides 45:43-48.
Raposo C, Schwartz M (2014) Glial scar and immune cell involvement in tissue remodeling and repair following acute CNS injuries. Glia 62:1895-1904.
Schwab ME (2004) Nogo and axon regeneration. Curr Opin Neurobiol 14:118-124.
Thuret S, Moon LD, Gage FH (2006) Therapeutic interventions after spinal cord injury. Nat Rev Neurosci 7:628-643.
Totoiu MO, Keirstead HS (2005) Spinal cord injury is accompanied by chronic progressive demyelination. J Comp Neurol 486:373-383.
Vanicky I, Urdzíková L, Saganová K, Cízková D, Gálik J (2001) A simple and reproducible model of spinal cord injury induced by epidural balloon inf ation in the rat. J Neurotrauma 18:1399-1407.
Copyedited by Moon SM, Liu L, Zhang M, Li CH, Song LP, Zhao M
*Correspondence to: J Luis Quintanar, Ph.D., jlquinta@correo.uaa.mx.
orcid: 0000-0003-1257-7723 (J Luis Quintanar)
10.4103/1673-5374.170311 http://www.nrronline.org/
Accepted: 2015-08-20
杂志排行

中国神经再生研究(英文版)的其它文章
- Intracellular sorting pathways of the amyloid precursor protein provide novel neuroprotective strategies
- The role of the Rho/ROCK signaling pathway in inhibiting axonal regeneration in the central nervous system
- VEGF in the nervous system: an important target for research in neurodevelopmental and regenerative medicine
- Studying neurological disorders using induced pluripotent stem cells and optogenetics
- Ef cacy of glucagon-like peptide-1 mimetics for neural regeneration
- Compliant semiconductor scaf olds: building blocks for advanced neural interfaces