Repair, protection and regeneration of peripheral nerve injury
2015-02-07Readingguide
Reading guide
SPECIAL ISSUE
Repair, protection and regeneration of peripheral nerve injury
Reading guide
1778 Repair of long-segment peripheral nerve defects
1779 Bionic reconstruction of hand function after adult brachial plexus root avulsion
1780 Optimized design of regeneration material for the treatment of peripheral nerve injury
1781 Synergism of electroactive polymeric materials and electrical stimulation promotes peripheral nerve repair
1783 Schwann cell ef ect on peripheral nerve repair and regeneration
1785 Biomaterials with specif c topological structures for the repair and regeneration of injured peripheral nerves
1787 Drug control of Schwann cells during peripheral nerve repair
1788 In vivo transplantation of green f uorescent protein-labeled neural stem cells delays denervation-induced muscle atrophy
1789 Key issues of stem cell therapy for peripheral nerve injury
1790 Repair, regeneration and remodeling of injured peripheral nerves
1792 Molecular mechanism of the regulation of neuron-intrinsic regeneration
1793 Neurotropism in peripheral nerve regeneration
1794 Intervention, repair, and protection of nerve injury in carpal tunnel syndrome
1795 Signif cance and strategy of promoting seed cell survival in tissue-engineered artif cial nerve construction
1796 Challenges for repairing peripheral nerve defects using 3D nerve grafts with nerve tissue-derived extracellular matrix and basement membrane tube-like conduits
Cite this article: Chen SL, Chen ZG, Dai HL, Ding JX, Guo JS, Han N, Jiang BG, Jiang HJ, Li J, Li SP, Li WJ, Liu J, Liu Y, Ma JX, Peng J, Shen YD, Sun GW, Tang PF, Wang GH, Wang XH, Xiang LB, Xie RG, Xu JG, Yu B, Zhang LC, Zhang PX, Zhou SL (2015) Repair, protection and regeneration of peripheral nerve injury. Neural Regen Res 10(11):1777-1798.
Shan-lin Chen*Department of Hand Surgery, Beijing Jishuitan Hospital
Zeng-gan Chen*Department of Orthopedic Surgery, Zhongshan Hospital, Fudan University
Hong-lian Dai#,*State Key Laboratory of Advanced Technology for Materials Synthesis and Processing, Wuhan University of Technology; Biomedical Materials and Engineering Research Center of Hubei Province
Jian-xun Ding#,*Key Laboratory of Polymer Ecomaterials, Changchun Institute of Applied Chemistry, Chinese Academy of Sciences Jia-song Guo*Department of Histology and Embryology, Southern Medical University; Key Laboratory of Tissue Construction and Detection of Guangdong Province; Institute of Bone Biology, Academy of Orthopedics
Na Han#,*Peking University People’s Hospital
Bao-guo Jiang*Peking University People’s Hospital
Hua-jun Jiang#,*Department of Orthopedics, First Af liated Hospital of Dalian Medical University
Juan Li#Department of Orthopedic Surgery, Zhongshan Hospital, Fudan University
Shi-pu Li*State Key Laboratory of Advanced Technology for Materials Synthesis and Processing, Wuhan University of Technology; Biomedical Materials and Engineering Research Center of Hubei Province
Wen-jun Li#Department of Hand Surgery, Beijing Jishuitan Hospital Jing Liu#,*The First Af liated Hospital of Dalian Medical University Yang Liu#Laboratory of Biotechnology, Dalian Institute of Chemical Physics, Chinese Academy of Sciences
Jun-xiong Ma#Department of Orthopedics, General Hospital of Shenyang Military Area Command of Chinese PLA, Rescue Center of Severe Wound and Trauma of Chinese PLA
Jiang Peng#,*Institute of Orthopedics, Chinese PLA General Hospital
Yun-dong Shen#Department of Hand Surgery, Huashan Hospital Af liated to Fudan University
Guang-wei Sun*Laboratory of Biotechnology, Dalian Institute of Chemical Physics, Chinese Academy of Sciences
Pei-fu Tang*Department of Orthopedics, the PLA General Hospital Gu-heng Wang#Department of Hand Surgery, Af liated Hospital of Nantong University
Xiang-hai Wang#Department of Histology and Embryology, Southern Medical University
Liang-bi Xiang*Department of Orthopedics, General Hospital of Shenyang Military Area Command of Chinese PLA, Rescue Center of Severe Wound and Trauma of Chinese PLA
Ren-guo Xie*Trauma Center, Department of Orthopedic Surgery, Shanghai First People’s Hospital, Shanghai Jiao Tong University School of Medicine
Jian-guang Xu*Department of Hand Surgery, Huashan Hospital Af liated to Fudan University
Bin Yu*Jiangsu Provincial Key Laboratory of Nerve Regeneration of Nantong University, Nerve Regeneration Collaborative Innovation Center
Li-cheng Zhang#Department of Orthopedics, the PLA General Hospital
Pei-xun Zhang#,*Peking University People’s Hospital
Song-lin Zhou#Jiangsu Provincial Key Laboratory of Nerve Regeneration of Nantong University, Nerve Regeneration Collaborative Innovation Center
Note: Arranged vertically in alphabetical order by their last name; #First author; *Corresponding author
Repair of long-segment peripheral nerve defects
The repair of long-segment peripheral nerve defects is obviously a dif cult clinical problem. For short-segment nerve defects (less than four times the diameter of the nerve), direct end-to-end neurorrhaphy can be applied by dissociating tissues surrounding the nerve or by adjacent joint f exion. However, for long-segment nerve defects that cannot be repaired by direct end-to-end neurorrhaphy, autologous nerve grafting is currently a preferred method of treatment. Simple nerve anastomosis may inhibit axon regeneration probably due to excessive tension. Moreover, direct nerve anastomosis can have certain error matching ratio and autologous nerve grafting also has some shortcomings such as lack of donors, increases trauma and needs multiple surgeries (Sabongi et al., 2015). Recently, the development of tissue-engineered nerve grafts and three-dimensional (3D) printing technology provides further possibilities for the treatment of peripheral nerve defects (Figure 1).

Figure 1 Treatment of long-segment peripheral nerve defects.
Allogeneic nerve has the advantage of having natural 3D nerve structure and is the preferred material used in grafts. However, it often results in immunological rejection. The major histocompatibility antigen of the allogeneic nerve is mainly present in Schwann cells, the myelin sheath and interstitial cells. Of these, the antigenicity of Schwann cells is the strongest and the current processing method of allogeneic nerves is to remove or deactivate cells so as to reduce the antigenicity of the allogeneic nerve. Usually, the immunogenic cells in grafts can be destroyed by freezing, radiation or chemical methods thus removing immune components and protecting the extracellular matrix structure. These methods can apparently decrease immunological rejection in allografts or xenografts and expand the source of nerve grafting. Clinical and experimental reports have shown that long-segment nerve defects have been successfully repaired, and various products have been used in the clinic (Brooks et al., 2012). However, the regeneration and conduction velocity of treated allogeneic nerves are still inferior to autologous nerve grafting. Current reports have conf rmed that the ef cacy of acellular nerve allografts (> 5 cm) is poor. Acellular nerve allograft scaf olds combined with Schwann cells and/ or stem cell transplantation has achieved good ef ects in animal experiments in vivo and in vitro, and is better than acellular nerve allografts alone. This method could be applied to clinical repair of long-segment peripheral nerve injury. Stem cells have multiple dif erentiation potentials and can dif erentiate into Schwann-like cells, promote the repair of peripheral nerve injury, and have a great application value. Many stem cells have been investigated, including embryonic stem cells, induced pluripotent stem cells, bone marrow mesenchymal stem cells, adipose-derived stem cells, olfactory ensheathing cells, and hair follicle stem cells. Stem cells can be isolated from embryonic tissue or autologous tissue. Schwann-like cells derived from dif erent stem cells have similar ef ects on nerve repair to Schwann cells.
Applied research of tissue-engineered nerve grafts provides more options for treatment of peripheral nerve defects (Li et al., 2013). Nerve conduits made of biomaterials or synthetic materials can only repair nerve defects less than 3 cm in size, and their ef ects are poor in the repair of nerve defects greater than 3 cm. Nerve conduits combined with nerve growth promoting factors such as Schwann cells, stem cells, small block muscle or nerve, neurotrophic factors and a variety of chemical and biological drugs may help to repair nerve defects greater than 3 cm in size. It is becoming the common view in the repair of long-segment nerve defects to select suitable nerve conduits according to the type of injured nerve while Schwann cells or stem cells can be used to promote axon regeneration simultaneously and it would be better to use local injection of various factors and drugs or additional electrical stimulation at the same time to promote nerve growth. Hydrogel and various composites formed by biological f brin, polysaccharides and proteoglycans are natural materials used to construct artif cial nerve grafts. However, natural materials used in the repair of nerve defects have some limitations on nerve defect length. Construction of synthetic materials can change material composition, structure and contents of various constituents, mechanical properties, degradation rate, porosity, permeability and surface shape, and can therefore meet the requirement for dif erent types of nerve injury. Synthetic materials combined with natural materials can make full use of the advantages of natural materials, alter the internal environment of the conduit using dif erent production processes, and better contribute to axon regeneration. Therefore, tissue-engineered nerve conduits combined with Schwann cells and stem cells is a possible direction for the repairment of long-segment peripheral nerve defects.
3D printing is a new type of digital prototyping technology which was gradually used in the late 1980s. In accordance with computer aided design models or tomography data and under computer control, layered printing which superimposes layer by layer, 3D printing precisely accumulates materials and rapidly makes 3D complex objects of arbitrary shape. 3D bioprinting technology can individually replicate in vitro the tissues and organs needed for patients or biomedicine. Currently, few studies have investigated the construction of artif cial nerve grafts using 3D printing technology. There was a study which tried to reconstruct the 3D structure of human brachial plexus (Chen et al., 2011). The internal structure of the brachial plexus was found to be very complicated. The fascicles bifurcated and recombined with one another with no f xed rules. So it is believed that with the development of image-reconstruction technique, the f ne structure of human nerves can be clearly revealed and reconstructed. 3D printing technology can help to construct individualized artificial nerves with a 3D structure and function similar to natural nerves, and also can even combine with dif erent cells and nerve growth promoting factors in a single process. This will help to further improve the therapeutic ef ects of long-segment nerve defects.
The repair of long-segment peripheral nerve defects is a very challenging task and requires continuous research and exploration. With the ongoing developments in science and technology, we believe that nerve repair ef ect will be further improved in the near future.
Juan Li, Zeng-gan Chen*
Department of Orthopedic Surgery, Zhongshan Hospital, Fudan University, Shanghai, China
*Correspondence to: Zeng-gan Chen, M.D., Ph.D.,
马克思主义认为,无产阶级及其政党必须具备无产阶级革命立场。否则,它就肩负不了实现共产主义的历史使命。列宁是在马克思主义指导下创造性地运用马克思主义取得俄国革命胜利的,他坚信马克思主义。他说:“严格的无产阶级世界观只有一个,这就是马克思主义。”[10]271因此,他没有任何理由不站在无产阶级立场上观察、思考和解决问题。列宁关于批评要有内容的思想正是站在无产阶级革命立场上提出来的。所以,他对任何非无产阶级立场都是反对的。列宁说:“如果批评自由是指维护资本主义的自由,那我们就要取消这种自由。”[6]36显然,在列宁那里,只要是有损无产阶级革命利益的批评,都是不允许的。
chen.zenggan@zs-hospital.sh.cn.
Bionic reconstruction of hand function after adult brachial plexus root avulsion
The brachial plexus is the web connection center for connecting the limb ef ector to motor neurons of the brain and anterior horn of the spinal cord. Brachial plexus injury can result in disability, typically known as “upper limb paralysis”. Through the 1960s, brachial plexus injury remained an incurable disease. With advances in medical technology and the application of extraplexal nerve transfer, the therapeutic ef cacy of brachial plexus root avulsion had gradually improved. In particular, the use of a contralateral C7nerve root transfer, created by Gu et al. (1989) brought a novel and ef ective treatment for brachial plexus avulsion. After unremitting ef orts for more than half a century, there is much improvement in the recovery of shoulder and elbow joint function (Dy et al., 2015). Muscle strength of the deltoid muscle in 82.9% of patients, the biceps brachii in 85.4% of patients, and the clavicular head of the pectoralis major in 92.7% of patients has reached grades 3–4, although there are still many problems to be solved regarding recovery of hand function (Wang et al., 2013). Even if the muscle strength of the hand recovers to grade 3 or greater, the hand remains handicapped with regard to voluntary movement, which greatly af ects daily life. The current techniques used for reconstruction of hand function are not very successful following brachial plexus injury, in particular with complete brachial plexus or middle and inferior trunk, or inferior trunk avulsion injury.
Present reconstruction situation for hand function following brachial plexus avulsion: The biggest dif erence between man and animal lies in the hands and the brain. The brain is responsible for thinking and coordinating, while the hands can be used to create and work. It has been hypothesized that the human brain likely evolved to employ creative thinking mainly through the use of hands. Why is the human hand so f exible? Because the hand has 19 intrinsic muscles and 23 extrinsic muscles. Simply speaking, complete hand function not only needs voluntary brain regulation, but also requires good shoulder and elbow function, stable wrist function, f nger f exion and extension, intrinsic hand muscle function, and sensitivity in the hand. The loss of control function of extrinsic and intrinsic muscles may appear after avulsion injury of the middle and inferior trunks of the brachial plexus. Because of axonal interruption, target organs lose the neurotrophic ef ect of motor cells from the anterior horn of the spinal cord, which ultimately results in hand muscle atrophy and f brosis, followed by loss of hand function, coordination, and brain-controlled function to the hand. Clinical research has, to date, been unsuccessful with the reconstruction of hand function following brachial plexus avulsion.
The methods of hand function reconstruction mainly include: (1) nerve transfer (a) reconstruction of f nger f exion; it is generally considered that ulnar nerve function is irreparable after brachial plexus injury, so median nerve repair is mainly used to reconstruct f nger f exion. According to dif erent types of injury, the following methods are commonly used: transfer of brachialis muscle branch, full-length phrenic nerve transfer, contralateral C7nerve root transfer, ulnar nerve bridging two-stage repair and reconstruction, intercostal nerve transfer, and contralateral C7nerve root transfer through anterior vertebral body to directly repair the inferior trunk. Among these methods, contralateral C7nerve root transfer through anterior vertebral body route obtains good therapeutic effects due to the shortest nerve regeneration distance and least number of anastomosis site. The muscle strength of f nger f exion of 64% patients, thumb f exion of 53% patients, and wrist joint f exion of 72% patients reaches a grade 3 or greater. (b) Reconstruction of f nger and thumb extension: the source of brachial plexus-innervated f nger and thumb extension has always been controversial. Clinical and basic research studies have verif ed that the source of f nger and thumb extension originates from the posterior division of the middle and inferior trunks, in particular the posterior division of the inferior trunk. On this anatomical basis, phrenic nerve transfer can be used to repair posterior division of the inferior trunk. Muscle strengths of extensor digitorum communis of 80% patients and extensor hallucis longus of 70% patients reach a grade 3 or greater. (c) Reconstruction of sensory function: sensory function of the hand is dominated by the radial nerve, ulnar nerve, and median nerve. The median nerve is the most important nerve for pulp of thumb, index, middle and radial side of ring f nger. Currently, the reconstruction of sensory functions primarily focuses on the median nerve. After contralateral C7transfer and two-stage repair of ulnar nerve bridging, sensory function recovery at S1–3can reach 61.29%. After contralatera C7transfer through anterior vertebral body route to directly repair the inferior trunk, sensory function recovery at S2–4reaches 88.6%, which primarily involves sensory in the middle f nger, ring f nger, and the hypothenar site. (2) Free functional muscle transplantation: Doi et al. (2013) from Japan highly recommends muscle graft reconstruction for brachial plexus avulsion, and reports 36 cases of gracilis-free muscle grafts. The recovery function outcomes were useful in 17 patients, who were followed up for at least 2 years. Barrie et al. (2004) used gracilis and intercostal nerves to reconstruct elbow f exion and wrist extension, f nger f exion, and sensory function of f ngers, and obtained functional recovery to varying degrees.
Bionic reconstruction of hand function: (a) Progress of bionic manipulator: previously, many amputees used a decorative prosthetic hand to cover the defect and to play a help in body balance. Since then, many amputees have used the rope-controlled prosthetic hand that is driven by shoulder, upper arm, and trunk movement. This prosthetic hand has the advantage of a simple design, low maintenance cost, durability, and can be used under moisture and dust conditions. Presently, electromyographic signal-controlled prosthetic hands are the f rst choice for amputees (Behrend et al., 2011). They can be controlled by surface electromyography signals on the arm stump, are feature-rich, and exhibit homologous control with the real hand. Recent trends show that the prosthetic hand has made great progress worldwide. The bionic prosthetic hand is close to the real hand in many aspects and can replace the human hand to some extent. Although research development for the prosthetic hand is rapid, the development of prosthetic hand control method is lagging and deserves further investigations. (b) Bionic hand reconstruction: the brachial plexus injury is involved in the middle and inferior trunks, will lead to loss of hand function. The long distance for brachial plexus nerve innervates of intrinsic muscles which antidenervation time is shorter than extrinsic muscle. Although great ef orts continue, muscle strength of useful f nger f exion has only reached 64% after nerve transfer or free-functional muscle transplantation, and the recovery of motor function is poor in approximately 40% of patients. Because recovery of sensory function remains poor, the random movement of limbs requires further research. These patients lose limb innervations, so they do not have indications for hand allotransplantation. Aszmann et al. (2015) from the Medical University of Vienna, Austria, f rst used bionic reconstruction in 2015 to reconstruct hand function after brachial plexus injury for three selected patients. Selective free-functional muscle transplantation, selective amputation, and bionic hand rehabilitation were used to restore hand function. The treatment consisted of two stages. During the f rst stage, ef ective electromyographic signals were identif ed and constructed to control the bionic hand. During the second stage, the injured hand was amputated, and the bionic hand was f xed. The Disabilities of Arm, Shoulder & Hand scores improved in three patients from 46.5 preoperatively to 11.7 postoperatively.
Prospects: Brachial plexus injury can result in serious dysfunctions. Presently, the reconstruction of random hand function continues to be dif cult. It is believed that a better understanding of the overall function of the nervous system and technological developments will provide novel treatments for hand function reconstruction after brachial plexus avulsion.
Department of Hand Surgery, Beijing Jishuitan Hospital, Beijing, China
*Correspondence to: Shan-lin Chen, M.D., Ph.D., drcsl@qq.com.
Optimized design of regeneration material for the treatment of peripheral nerve injury
Peripheral nerve defects are a common disabling injury. Peripheral nerve injuries arise from trauma, cancer, or congenital defects, and are challenging clinical issues to address. According to statistical data from the World Health Organization, there are 10–15 million new trauma cases reported worldwide every year, of which 15–40% involve peripheral nerve injury. In China, 600,000–900,000 new cases of peripheral nerve injury are reported annually, approximately half of which require nerve grafting. However, long-segment peripheral nerve defects and injury to multiple nerves cannot be repaired in the clinic, and neuromas form readily during the process of repair. The aim of treatment for peripheral nerve defects is the recovery not only of structure, but also sensory and motor function. Therefore, nerve regeneration and functional recovery after peripheral nerve damage remain important topics in the f eld of neuroscience. In this review, we highlight a recent development in nerve regeneration surgery.
Materials used in the repair of peripheral nerves include autologous and allogeneic nerve grafts, autologous non-neural tissue grafts and biomaterials. Autologous nerve grafting is the gold standard for nerve repair; however, its source is limited, and it can damage the donor site. Allogeneic nerve grafts require concurrent treatment with immunosuppressants, and the technique has a low success rate. Autologous non-neural tissue grafts are insuf ciently stable and collapse easily. Inert nerve conduits can delay the recovery of the regenerating nerve, causing long-term complications; furthermore, they require a second surgical procedure for their removal. To date, the main focus of studies in this f eld has been on absorbable nerve conduits, such as polyglycolic and polylactic acids, polylactic acid-poly(ε-caprolactone) copolymers, chitosan and collagen. These conduits provide a temporary environment for nerves undergoing regeneration. They are degraded and absorbed in vivo, avoiding compression and inf ammation on newly formed nerves and further invasive surgery for their removal. However, the degradation products of some polymers used in these nerve conduits are acidic, and not conducive to the growth and proliferation of nerve cells. This reduces the speed and quality of repair, rendering nerve conduits insuf ciently ef ective in the repair of long-segment peripheral nerve defects.
When a nerve f ber breaks, the distance between the two stumps might be too great for unaided repair, or the severed nerves may fail to connect properly. This can obstruct axonal growth in all directions, inducing f exion, axonal wrapping, connective tissue hyperplasia and neuroma formation, preventing the restoration of nerve function. Nerve conduits can provide a relatively isolated microenvironment for nerve regeneration, rich in neurotrophic factors, and with reduced f broblast inf ltration. Although this can contribute to axon growth, the conduit is only a physical channel for nerve growth and cannot induce nerve regeneration or provide a microenvironment that supports repair (Moore et al., 2009). Furthermore, tissue f uid in the conduit is not conducive to nerve growth, and has the risk of neuroma formation (Moore et al., 2009).
Improvement in the speed and quality of nerve repair, and inhibition of neuroma formation, are the core aims of treatment strategies for peripheral nerve injury. The axonal growth cone in a damaged nerve detects the surrounding environment and guides axon regeneration. Therefore, the conduit plays an important role in nerve repair, not only acting as a temporary scaf old and providing a good microenvironment for nerve growth, but also promoting axon regeneration. Successful functional regeneration depends on three elements: (1) survival and functional status of neuronal cell bodies; (2) rate of axonal regeneration; (3) nerve growth in the target area. Nerve growth factor is essential to all three elements. However, the main problems arising from the addition of exogenous nerve growth factor to conduits are that its half-life is very short, and the dose is dif cult to determine. Therefore, this raises the question of how to promote the secretion of bioactive factors that induce nerve f ber growth towards the target area, by changing the composition of the conduit material.
After nerve injury, alterations in the local microenvironment and release of proinf ammatory cytokines cause f broblast activation and collagen hyperplasia. Scar formation and f brosis occurs, resulting in the development of neuromas. Therefore, neuroma growth can be avoided by maintaining the concentration and activation of nerve growth factor and inf ammatory factors, promoting axon regeneration, avoiding f broblast activation, maintaining the balance of collagen synthesis and degradation and ensuring the correct anastomosis of the nerve. The development of a biochemical signaling molecule that can induce nerve regeneration, and a biomaterial that provides a good microenvironment for nerve growth, is therefore needed in the f eld of nerve repair.
The ideal nerve conduit meets following requirements: (1) good cytocompatibility; (2) biodegradability (rates of degradation and metabolism should match that of nerve repair); (3) suitable mechanical properties, providing a three-dimensional space for nerve growth; (4) materials that can interact with cells, improving cell viability and inducing nerve regeneration; (5) high porosity, oriented pore structure, and a connected and uniform conduit interior, convenient for transmission and distribution of nutrients and cellular metabolites.
Accordingly, we have invented a biomimetic nanocomposite nerve repair material with three components: composite-introduced RGD peptide modif ed poly{(lactic acid)-co-[(glycolic acid)-alt-(L-lysine)]} (PRGD) promotes cell migration, adhesion and proliferation, and induces nerve regeneration; poly-D,L-lactide acid (PDLLA) meets the requirements for mechanical strength and degradation rate to match the rate of nerve regeneration in nerve defects by regulating molecular weight; β-tricalcium phosphate (β-TCP) neutralizes the acidic environment produced by PDLLA degradation products, prevents aseptic inf ammation, and provides calcium ions (Pachuau et al., 2014) that aid synthesis and secretion of neurotrophic factor, to promote the growth of regenerating nerves. Animal and clinical studies conf rmed that the material, PRGD/PDLLA/β-TCP, has good biocompatibility and biodegradability, and induces nerve regeneration, reaching an excellent rate of nerve repair (73.7%; Huang et al., 2013) comparable to the clinical performance of autologous nerve grafts (Figure 1). Furthermore, no evidence of neuroma formation was found in sciatic nerve repair tests in approximately 300 rats and dogs, and over 40 clinical tests. In nerve regeneration chambers, PRGD/PDLLA/β-TCP-constructed cytokines and degradation products reduced the inf ammatory response, inf uenced the matrix-degrading enzymes and the transforming growth factor-β/Smad signaling pathway, reduced the synthesis, secretion and excessive deposition of collagen, and suppressed scar formation. Growth factor enrichment promoted myelin repair, increased the ratio of myelinated to unmyelinated nerves, reduced pain, improved the quality of nerve regeneration, and decreased the occurrence of neuroma. PRGD/PDLLA/β-TCP composites induced nerve regeneration and inhibited neuroma formation.
In summary, optimizing design of the composition and structure
Synergism of electroactive polymeric materials and electrical stimulation promotes peripheral nerve repair
Electroactive materials are mixed ionic-electronic conductors that can become electrical conductors by reversible ion exchange in the oxidation-reduction state. Electroactive materials contain inorganic transition metal complexes, inorganic/organic hybrid materials, electroactive polymeric materials (EPMs), and so on.
EPMs are an important component of electroactive materials. Due to their unique conjugate structure and doping mechanism, EPMs exhibit superior physicochemical properties, including adjustable electrical response, good biocompatibility, and easy processing. Thus, EPMs have the great potential for broad application in the f elds of catalysis, anti-static electricity, selective electrodes, gas separation membranes, electrochemical sensors, metal corrosion, electromagnetic shielding, optoelectronic devices, stealth technology, molecular wires, biomedicine, and so on. As shown in Table 1, EPMs mainly comprise polyacetylene (PA), polypyrrole (PPy), polythiophene (PTh), poly(p-phenylene) (PPP), poly(phenylene vinylene) (PPV), and polyaniline (PANi) (Guimard et al., 2007). Simultaneously, various derivatives are gradually developed based on these basic structures to elevate EPMs performances and expand their applications.
In the f eld of biomedicine, PPy, PTh, and PANi are widely used in anti-oxidation and anti-bacterial applications, biological imaging, controlled drug release, artif cial muscle, and bone and nerve repair. PPy was deployed early and extensively due to its advantages, including simple synthesis, high conductivity, easy f lm formation, softness, oxidation resistance and good biocompatibility. PTh has a similar chemical structure of aromatic ring to PPy. PTh is easily synthesized, has good stability, and exhibits adjustable conductivity from insulation to metal characteristics. Among them, PANi is the most promising EPM, because of its simple synthesis, stable performance, good conductivity, and easy processing.
Electrical stimulation has been shown to induce adhesion, migration, proliferation, and dif erentiation of f broblasts, osteoblasts, and nerve cells. EPMs have been used as tissue-engineered materials to ef ectively promote tissue repair by transmitting electrical stimulation to a lesion site. Due to the electrical signal-related physiological structures and functions of nerve cells and tissue, electroactive polymers in combination with electrical stimulation have been used for nerve repair (Figure 1).
EPMs were f rst researched for peripheral nerve repair in 1997 when Schmidt et al. (1997) reported that the oxidation state of PPy doped with poly(styrene sulfonate), as a matrix, had favorable cell adhesion performance, strengthened cell interaction, and promoted axon growth of PC12 cells, that is, a rat adrenal pheochromocytoma cell line from a tumor of the rat sympathetic nervous system. In addition, all cell behaviors on the electroactive polymeric matrix could be further promoted by the supplementation of electrical stimulation. Subsequently, Liu et al. (2011) synthesized an oxidation state of PPy substrate doped with chondroitin sulfate conjugated with type I collagen. PC12 cells grown on the above substrate demonstrated improved cell dif erentiation and axon growth, and the combination with electrical stimulation demonstrated even greater improvement. Recently, Forciniti and colleagues investigated the interaction mechanisms of Schwann cells and PPy substrate in of nerve conduit materials will improve the repair of long-segment peripheral nerve defects, prevent neuroma formation, and restore peripheral nerve function. However, the mechanisms underlying material-induced nerve regeneration, and the development and prevention of neuroma formation during nerve repair, remain poorly understood. A number of questions remain unanswered, including which properties or factors of the material regulate neuronal behavior and specif c gene expression, which components of the material are most important in the inhibition of neuroma development, and how specif c factors in the material convert to signaling molecules that the body recognizes. In addition, the relationships between the properties, composition and structure of the material, and its ef ects on cytokine secretion, nerve regeneration and neuroma formation, warrant further investigation.
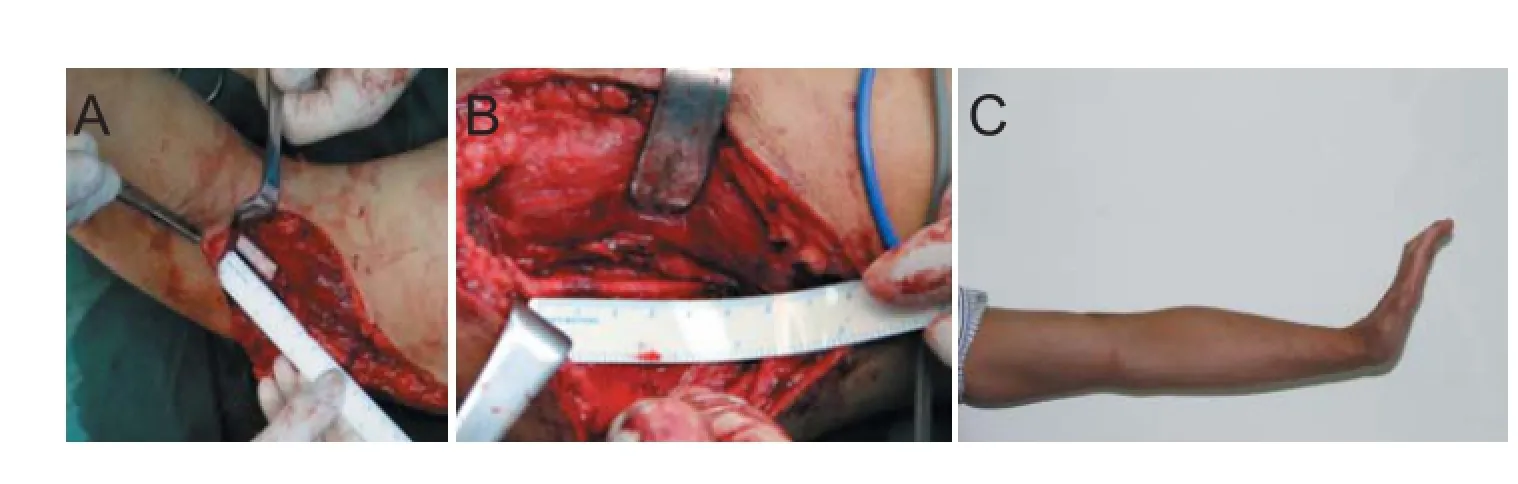
Figure 1 PRGD/PDLLA/β-TCP nanocomposite in the repair of a 2 cm-long radial nerve defect.
Hong-lian Dai*, Shi-pu Li*
State Key Laboratory of Advanced Technology for Materials Synthesis and Processing, Wuhan University of Technology, Wuhan, Hubei Province, China (Dai HL, Li SP)
Biomedical Materials and Engineering Research Center of Hubei Province, Wuhan, Hubei Province, China (Dai HL, Li SP)
*Correspondence to: Hong-lian Dai or Shi-pu Li, daihonglian@wuht.edu.cn or lishipu46@126.com.an electric f eld. The authors found that the PPy substrate absorbed multiple serum proteins in the electric f eld and could contribute to the net migration of Schwann cells toward the anode (Forciniti et al., 2014). Furthermore, the speed of Schwann cell migration was controlled by an integrin-receptor-mediated mechanism, and the direction of Schwann cell migration was dominated by the phenomenon of electrical conductivity.
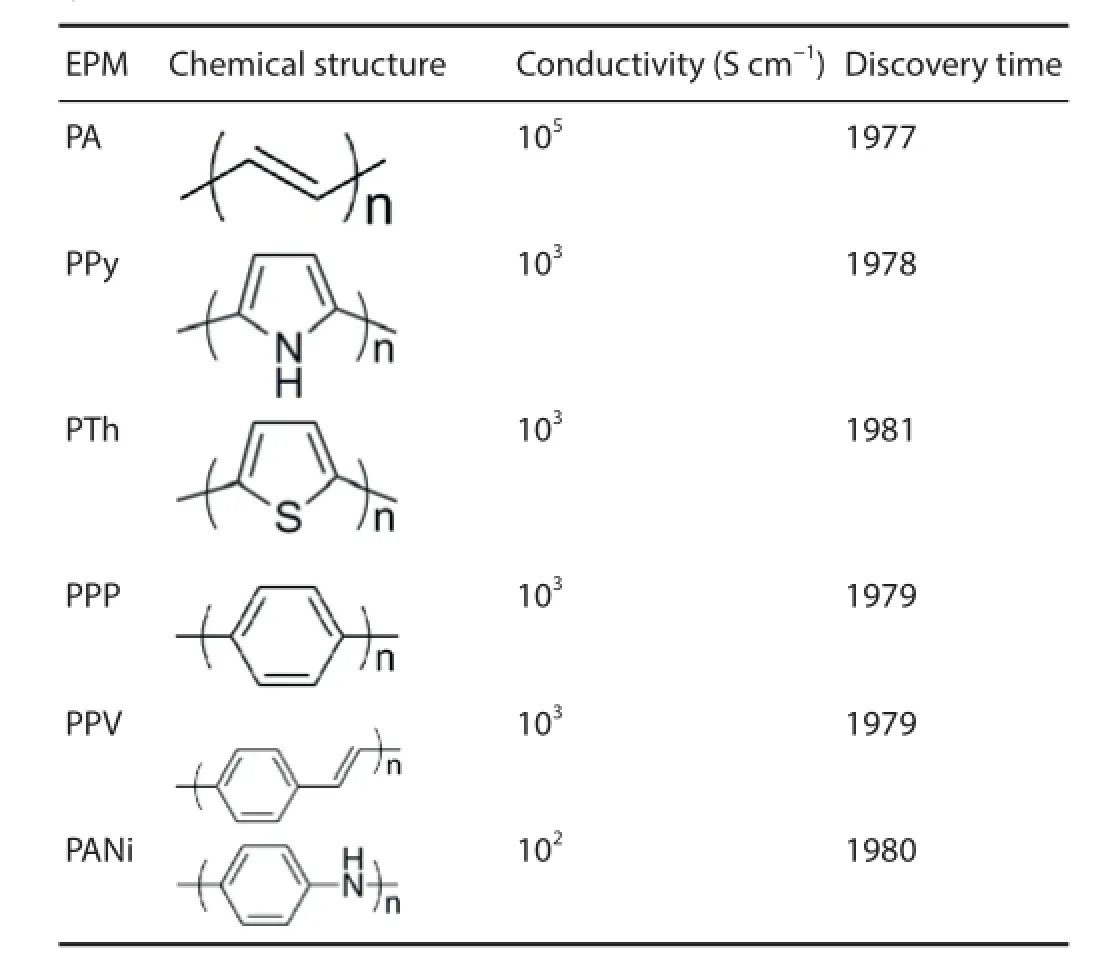
Table 1 Chemical structures, conductivities, and discovery years of typical EPMs
Thus far, investigators have mostly explored the synergistic effects of EPMs and electrical stimulation on the dif erentiation and axon growth of nerve cells. The non-biodegradability of EPMs limits their applications in living organisms. To address this limitation, some EPMs were synthesized based on electroactive oligomers and applied to enhance the behaviors of nerve cells. In 2002, a polymer was synthesized from conducting oligomers of pyrrole and thiophene linked by ester linkages (Rivers et al., 2002). The resulting polymer is conductive, degradable, and biocompatible (Rivers et al., 2002). Researchers from Changchun Institute of Applied Chemistry, Chinese Academy of Sciences, designed and synthesized a series of EPMs based on aniline tetramer and pentamer. For example, the multiblock copolymers composed of aniline pentamer and polylactide (PLAAP) were synthesized by condensation reaction in this research group (Huang et al., 2008). Combined with electrical stimulation, the PLAAP multiblock copolymer doped with camphorsulfonic acid promoted dif erentiation and axon growth of PC12 cells, which are proven to be a potential candidate for use in the repair of peripheral nerves.
Figure 1 Peripheral nerve reconstruction through synergism of EPMs and electrical stimulation.
The studies addressing the synergistic use of EPMs and electrical stimulation in the repair of peripheral nerves after injury are currently ongoing, which are only investigating ef ects on the behavior of nerve cells. However, EPMs have obvious advantages and great potential for repair of peripheral nerves after injury: (1) Electroactive oligomers linked to dif erent small molecules or polymers can assign dif erent performances to EPMs, such as biodegradability and stimuli-responsiveness; (2) EPMs are f exible and provide varying surface properties by doping with dif erent ions; (3) electrical stimulation can be concentrated in areas around the polymer by using EPMs, allowing precise behavior regulation of nerve cells; (4) nerve cell matrix analogs and growth factors can bind to the surface of EPMs by physical interactions and further enhance nerve regeneration.
In summary, the synergism of EPMs and electrical stimulation for treatment of peripheral nerve injury is expected to develop in the following four directions: (1) Developing multifunctional copolymers based on electroactive oligomers instead of EPMs of high molecular weight; (2) constructing EPM nerve conduits using tissue engineering methods for application in vivo as soon as possible; (3) fabricating a device that can ef ectively and precisely provide electrical stimulation in vivo; (4) combining EPMs to bioactive molecules and electrical stimulation to ef ectively achieve peripheral nerve repair. Although the emerging tissue engineering protocols from EPMs and electrical stimulation in the repair of peripheral nerves after injury have not been tested clinically and even in vivo, the combination of methods would exhibit good clinical viability.
Jian-xun Ding*, Wei-guo Xu
Key Laboratory of Polymer Ecomaterials, Changchun Institute of Applied Chemistry, Chinese Academy of Sciences, Changchun, Jilin Province, China
*Correspondence to: Jian-xun Ding, Ph.D., jxding@ciac.ac.cn.
Schwann cell ef ect on peripheral nerve repair and regeneration
Following peripheral nerve injury, distal and proximal fibers undergo a series of changes in cell and molecular biology that can result in hypoesthesia, neuropathic pain, loss of motor function and muscle atrophy in the target area. Nerve injury treatment and subsequent regeneration and functional recovery are historical clinical problems. Currently, studies concerning peripheral nerve repair mainly focus on anastomosis, nerve grafting and bridging nerve defects. However, the ef ect of repair is insuf cient and unsatisfactory; it is reported that after surgery, original functionality is recovered in only approximately 10% of patients. The largest dif erence between nerve tissue repair and other tissue repair is that nerve tissue cannot be replaced by scar tissue, and can only be restored by self-repair.
Morphological regeneration of peripheral nerves is not the same as functional recovery of target organs. Even if regenerating nerves recover well morphologically, the nerve often cannot exert its functions. Therefore, for peripheral nerve injury, the important aim is not to recover its structure, but to fully restore its function. Achieving peripheral nerve function regeneration depends on three basic elements: (1) survival and functional status of neuronal cells, ensuring the sprouting and extension of proximal axons of injured nerves; (2) ensuring specif c choice of distal and proximal nerve f -bers, and that axon regeneration is in the correct direction; (3) suff cient speed of nerve regeneration, and re-domination and functional reconstruction of the terminal ef ector. Schwann cells are glial cells in the peripheral nervous system and play an important role in peripheral nerve function regeneration. Following peripheral nerve injury, Schwann cells produce a series of physiological and biochemical changes. Regeneration can be observed 3 hours after axon injury and mainly appears in the nodes of Ranvier near to the damaged end. Two days later, Schwann cells grow from the proximal end and migrate along the direction of axon regeneration. The speed of some Schwann cell migration exceeds that of axons, forming Bungner bands to guide axon regeneration (Torigoe et al., 1996).
The survival and functional status of neuronal cells determine the potential for nerve regeneration after peripheral nerve injury; in early neural development, many processes form around neuronal cells. Ultimately, only one process can form an axon and other processes become dendrites. During neural development, to quickly and accurately perceive the external environment and result in a corresponding action, axons must continue to grow until they reach the dominant target. Neurons regulate axon growth at a certain rate and growth discontinues at the right time. Following peripheral nerve injury, a sudden interruption of axons will cause oxidative stress in neuronal cells. Intracellular calcium overload and reactive oxygen species interfere with mitochondrial function, and can even lead to neuronal apoptosis. The intensity of this reaction is associated with the degree of peripheral nerve injury and the distance between injury level and the parent cell body. That is, severe peripheral nerve injury indicates a close distance between injury level and the parent cell body and severe neuronal injury. A previous study conf rmed that degeneration of dopaminergic neurons was associated with inhibition of mitochondrial complex I in rat models of Parkinson’s disease induced by 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (Perier et al., 2005). In the above models, mitochondria release cytochrome C, and p53, caspase-3 and caspase-9 are simultaneously activated. Proapoptotic protein Bax expression is upregulated and leads to apoptosis after translocation to mitochondria. Nevertheless, if the injured nerve is immediately repaired, neuronal loss can be minimized to a large degree, which indicates that neurotrophic factors may play a protective role (Ma et al., 2003). Schwann cells play a crucial role in the above process, and the microenvironment surrounding axons changes. Schwann cells on the proximal end produce a large number of neurotrophic factors such as ciliary neurotrophic factor, neurotrophin 3 and f broblast growth factor, which can be transported into neurons by axoplasm. These neurotrophic factors trigger the protection mechanism of neurons. For example, ciliary neurotrophic factors that make contact with damaged axon receptors improve STAT3 phosphorylation levels. STAT3 retrograde transport to the nucleus triggers neural changes (Zhang et al., 2011). More importantly, after injury, Schwann cells can secrete abundant nerve growth factors that retrogradely transport to the cell body by endocytosis and maintain neuronal survival.
Specif c choice of nerve f bers in proximal and distal ends and correct direction of axon regeneration are important. Regenerated motor axons often tend to prefer reinnervated muscle, i.e., they have preferential motor reinnervation, which is also called selective motor nerve regeneration. When nerve regeneration occurs, regenerated motor neurons will be preferentially reinnervated in muscular branches (Redett et al., 2005). It is a trend that regenerated motor axons selectively redistribute in muscle tissue. However, regenerated motor axons can produce many lateral buds, which initially randomly distribute to Schwann cells of sensory or motor f bers. With prolonged time, lateral buds that falsely project to sensory pathways are trimmed, but those projecting to motor pathways are retained, resulting in a specif c projection (Witzel et al., 2005).
Clinically, if the continuity of the endoneurial tube is still in existence (as is the case with crush injury), and only axon injury is detected, the functional recovery is frequently good. If the nerve is completely transected, it is very dif cult to repair, because axonal mismatch occurs. That is, regenerated motor axons reach sensory end organs along a catheter formed by Schwann cells in the sensory branch. The mismatched axons cannot exercise their normal functions, but also obstruct the growth of axons because of its mass ef ect. Thus, regenerated nerves cannot exert their normal function and prognosis is poor.
Traditionally, there are two types of Schwann cells (myelinated and unmyelinated). Höke et al. (2006) considered that Schwann cells in motor pathways and sensory pathways have dif erent phenotypes, and that the secreted neurotrophic factors are also dif erent. Jesuraj et al. (2012) verif ed that sensory neurons and motor neurons present noticeable dif erences in the expression of certain proteins. Thus, Schwann cells can be divided into motor and sensory types, which play dif erent regulatory roles in neuronal growth. Moreover, the direction of axon growth is adjustable along the changes in neurotrophic factor concentrations at the distal end.
Dif erent phenotypes of Schwann cells and Schwann cells presenting dif erent nutritional factors and adhesion proteins af ect the occurrence of preferential motor reinnervation. However, the reasons for the changes in Schwann cell phenotypes and the method of af ecting preferential motor reinnervation deserve further investigation.
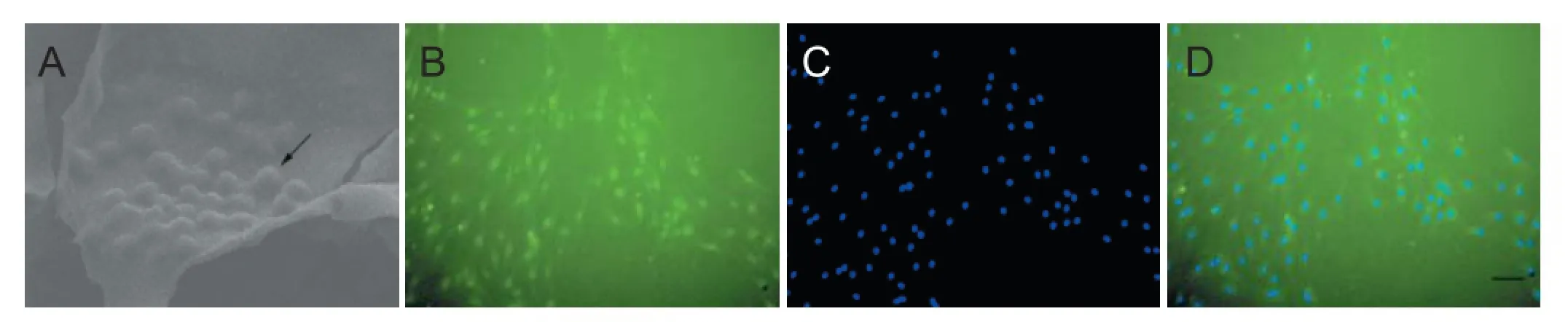
Figure 1 Morphology of Schwann cells after culture on the scaf olds (arrow; A), S-100 staining (B), DAPI nucleus counterstaining (C), and the merging of B and C (scale bar: 50 μm; D).
Another important factor is sufficient speed of nerve regeneration and reinnervation and functional reconstruction of the terminal ef ector. The ef ects of peripheral nerve repair, especially brachial plexus repair, are poor. The main reason for this is that the speed of axon growth is slow after peripheral nerve repair. When translocated or transplanted proximal axons grow at the distal end of the upper limb, the distal end innervated by the brachial plexus (especially the function of the hand) has shrunk. Therefore, methods to elevate the speed of axon regeneration and accelerate nerve repair are important in current research on nerve injury.
Schwann cells also exert a key ef ect in elevating the speed of nerve regeneration. Following peripheral nerve injury, abundant Schwann cells proliferate and greatly secrete nerve growth factor. High concentrations of nerve growth factor form at the injury site, promote nerve regeneration, and probably regulate axon growth by nerve growth factor-MEK-mitogen-activated protein kinase pathway. Simultaneously, Schwann cells secrete extracellular matrix and cellular adhesion molecules after peripheral nerve injury (May et al., 2004). These macromolecules play the sticking and guiding roles in nerve regeneration. The mechanism of axon growth promotion is that cellular adhesion molecules secreted by Schwann cells bind to corresponding molecules on the surface of axonal growth cones and contribute to the secretion of laminin and f bronectin from Schwann cells. The above factors bind to receptors on the surface of growth cones and induce changes in the structure of growth cones and prolong axons.
Schwann cells, as seed cells, have therefore become the key in tissue-engineered neural construction. Sun et al. (2009) and Jesuraj et al. (2011) added Schwann cells in chemically extracted acellular nerve allografts to enhance the ef ects of peripheral nerve regeneration. Moreover, a series of studies have been conducted on how to ensure the long-term survival of seed cells (Schwann cells) in scaf olds (Figure 1) and to fully exert their ef ects. (1) Elevating biological activity of Schwann cells: Keilhof et al. (2000) cut neuroma during nerve stump trimming and isolated Schwann cells; they found that this method obtained abundant Schwann cells with good viability and little rejection. (2) Improving cell function: with the development of molecular biology, it will be possible to maintain and enhance the function of Schwann cells by transgenic technology. Menei et al. (1998) believed that genetically modif ed Schwann cells that can secrete human brain-derived neurotrophic factor contributed to axon regeneration in rats with spinal cord transection. (3) Establishing an immortalized cell line: Wang et al. (2009) conf rmed that immortalized Schwann cells had a high aff nity to chitosan, and the ef ects on repairing sciatic nerve defects were evidently better than that of the control group. This may be due to the long-term survival of implanted Schwann cells and effective secretory function.
Hirata et al. (2001) established a one-to-one relationship between axons and Schwann cells and observed massive apoptosis of Schwann cells through downstream regulation, characterized by increased intracellular ceramide levels. This coincided with neuronal survival and apoptosis during normal sympathetic development of newborn rats. There is a dynamic equilibrium between P75NTR and TrkA during sympathetic differentiation. This equilibrium determines the correct establishment of a relationship between neurons and target organs. The regulation of nerve regeneration also has a kind of equilibrium. An exogenous high concentration of nerve growth factor contributes to axon regeneration. However, nerve growth factor also promotes Schwann cell apoptosis. Taken together, the addition of Schwann cells or exogenous nerve growth factor cannot improve the speed and quality of nerve regeneration. The reason for this may be that the number of Schwann cells is strictly controlled during myelination, and the relationship between axons and Schwann cells is one-to-one. Schwann cells that do not make contact with axons undergo apoptosis because they cannot obtain the necessary survival signals from axons. That is, once Schwann cells wrap around axons, proliferation is stopped, resulting in myelinogenesis (Meyer zu Hörste et al., 2006). Thus, the relationship between Schwann cells and axons requires further study.
In conclusion, the ef ects of Schwann cells vary following peripheral nerve injury. Schwann cells undergo phagocytosis, provide a stent and neurotrophic factors for nerve regeneration, maintain neuronal survival, guide ordered axon extension, promote axon myelination, and regulate their own survival and apoptosis. However, achieving real functional regeneration of peripheral nerves involves axon regeneration, Schwann cell myelination, various neurotrophic factor intervention, target organ protection, reinnervation and central nervous system re-adjustment. Some aspects deserve further research, including activation of neuronal growth, regulation of environmental factors on axonal growth, molecular mechanisms of Schwann cell myelination, and accurate and ef ective docking of dif erent f bers.
Hua-jun Jiang*
Department of Orthopedics, First Affiliated Hospital of Dalian Medical University, Dalian, Liaoning Province, China
*Correspondence to: Hua-jun Jiang, Ph.D., jianghuajun112@163.com.
Biomaterials with specif c topological structures for the repair and regeneration of injured peripheral nerves
Peripheral nerve injury can result in severe paralysis and dysfunction. For a long time, achieving the repair and regeneration of injured peripheral nerves has been an urgent goal in the f eld of medical science. Most of the recent studies on peripheral nerve repair and regeneration have concentrated on anastomosis, nerve transplantation, and artif cial nerve bridging. The key factors and precise mechanism driving peripheral nerve regeneration remain poorly understood, which severely restricts the development of clinical treatments. There is an urgent need to construct ef ective in vitro biological models for studying peripheral nerve repair and regeneration to solve the problem of how to treat peripheral nerve injury as soon as possible and to reduce the family and social burdens caused by such diseases.
The importance of material-cell contact and interactions for regenerative medicine is increasingly being recognized. Harrison (1911) found that the topological structure of a substrate af ected cell orientation, migration, and cytoskeletal structure by studying the biological behavior of cells grown on spider webs. This phenomenon increased attention on the interactions between biomaterials with a specif c topological structure and cells. With the rapid development of micro- and nano-material fabrication technologies for designing and controlling the surface of biological materials, the three-dimensional micro- and nano-topological structure of a material can be selected to simulate well the physiological extracellular matrix texture. In particular, micro- and nano-materials have been extensively applied in the repair and regeneration of cell populations within the highly oriented structures in nerves and tendon.
The preparation of peripheral nerve substitutes, i.e. biomaterials with a specif c topological structure, is of interest to the f eld of peripheral nerve injury to address several current problems, including the limited supply of biological grafts and immune rejection during autologous or allogeneic nerve grafting for the repair of peripheral nerve defects. For example, Ahmed et al. (2004) rolled the thin f lms formed by the evaporation of collagen solutions into tubes, creating nerve conduits after glutaraldehyde crosslinking. Fourier transform infrared spectroscopy showed that the entire molding process did not alter the collagen triple helix structure. Moreover, microscopic analysis indicated that the porous structure on the surface of the material contributes to axonal sprouting, adhesion, and growth from the proximal end of the injury site. Wang et al. (2014) developed a self-assembling peptide nanof ber scaf old-containing PLGA conduit and used this artif -cial nerve graft to bridge a 10-mm-long sciatic nerve defect in the rat, suggesting the potential of this material in repairing peripheral nerve injury. Yang et al. (2015) proposed a nerve guidance conduit made from silk f broin and PLGA through electrospinning and weaving. In vivo and in vitro experiments showed that the material was biocompatible and maybe reliable scaf olds for peripheral nerve repair.
Our research group simplifies scientific problems following clinically feasible strategies and investigates several key issues for peripheral nerve repair and regeneration, including improving the preparation and application of biomaterials with specif c topological structures, electrophysiology, and directional nerve cell growth. To this aim, we construct scaf olds from nano-silk f broin, porous silk f broin materials, conductive carbon nanocomposites, composite nanof brous scaf olds, and drug delivery carrier–cell scaf olds, in which the material degradation time corresponds to the pathological stage of peripheral nerve regeneration. These materials provide a three-dimensional structure for nerve repair, nutritional support for nerve regeneration, and are carriers of the seeded cells and related drugs. Several of our research results are summarized below.
Functional design of a nerve tissue-specif c glycerol-silk f broin material: To accurately mimic the low mechanical flexibility of neural extracellular matrix, we mix silk f broin and glycerol in a water soluble process to prepare silk f broin scaf olds with a variety of nanostructures to successfully simulate the natural nanostructures of neural extracellular matrix. The secondary structure of glycerol-silk f broin materials can be adjusted using the glycerol content. Controlling the mechanical properties and degradation rate of the scaf old allows the creation of scaf olds with satisfactory biocompatibility and adjustable mechanical properties. We expect this glycerol-silk f broin material to be an extremely valuable biological scaf old for the repair of peripheral nerve injury.
Improving the preparation of electrospun catalpol-loaded composite nanof brous scaf olds: Fabrication method of catalpol loading into the electrospun poly(lactic-co-glycolic acid)/multi-walled carbon nanotubes/silk f broin nanof brous scaf olds is successfully achieved. Immunocytochemistry, RT-PCR, and western blot assays are performed to evaluate hASCs neuronal dif erentiation, and it is shown that catalpol could signif cantly upregulate the expressions of βIII-tubulin and Nissl. The hASCs could be directly induced into neuronal-like cells, which might of er new avenues in nerve regeneration (Figure 1).
Oriented stem cell scaffold-dependent drug delivery carrier: To address the lack of induction of neural dif erentiation in most scaf olds, we design functional drug delivery carriers by altering dif erent material properties. Using silk f broin nanoparticles as a modif er, monodisperse vaterite microspheres of small particle size can be prepared using the co-precipitation method. The crystal structure and nanostructure of the microspheres can be adjusted with multi-parameter variability. The microspheres show high encapsulation ef ciency and pH response, but no burst release.
Regarding the conductive carbon nanocomposites, how to precisely match the biomaterial and inducer to control neural dif erentiation of adipose-derived stem cells remains dif cult. One solution is to transform polymers, carbon nano-materials, and catalpol into specif c “printing ink”. Using 3D bioprinting, drug delivery carrier–cell scaf olds are constructed using multi-nozzle extrusion. Such scaf olds can improve the neural dif erentiation of adipose-derived stem cells, the maturity of dif erentiated cells, and exhibit appropriate transport properties.
We have successfully developed an electrospinning apparatus, bioprinting machine, and a multi-layer three-dimensional perfusion bioreactor to establish three-dimensional neural networks. These are valuable tools for studying in vitro directed differentiation. Following the structural features of nervous tissue and regulatory mechanisms of neural regeneration, self-assembly of the electrospinning apparatus is done by processing key components of the device outside of China and assembling the main components in China. Patents applications have been f led for these scaf olds and technologies. The advantages of the perfusion cell culture system and three-dimensional cell culture allow us to culture cells under continuous dynamic perfusion, which more accurately mimics the in vivo cell growth environment. In addition, this setup achievesf ow exchange in the microenvironment, improves the gas control and multi-layered scaf old structure, controls the multiple environments necessary for neural dif erentiation of stem cells, simplif es various technical parameters of three-dimensional culture, and successfully creates three-dimensional cell models in vitro.
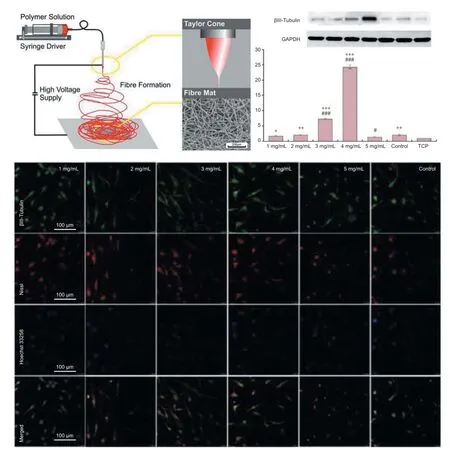
Figure 1 Potential neurogenesis of human adipose-derived stem cells on electrospun catalpol-loaded composite nanof brous scaf olds.
Following the requirements for clinical application, we independently designed and established a Research and Development Application Platform in Stem Cells and Regenerative Medicine, which was certif ed by the internationally recognized Good Manufacturing Practice Regulations. This was the f rst design criterion indexed by the international English reference book. This stem cell transformation platform, from the view of evidence-based medicine, optimizes clinical indications, standardizes operating procedures, provides a quality inspection system and indexes for monitoring the therapeutic ef ect, and promotes the clinical transformation of organ-specif c stem cell colonization scaf olds.
As shown by our numerous previous studies, we integrate novel ideas for peripheral nerve repair and regeneration. Validation of these ideas and the related clinical plan will increase the range of peripheral nerve injuries that can be repaired and improve the effects of peripheral nerve injury repair.
Jing Liu*, Jing-yun Ma
The First Af liated Hospital of Dalian Medical University, Dalian, Liaoning Province, China
*Correspondence to: Jing Liu, Ph.D., liujing.dlrmc@hotmail.com.
Drug control of Schwann cells during peripheral nerve repair
Peripheral nerve damage is a common injury in trauma patients. It can cause sensory loss, motor dysfunction and peripheral neuropathy. At present, more than 1 million people worldwide are af ected by peripheral nerve injury caused by traf c accidents, disease or iatrogenic injury (such as tumorectomy and orthopedic procedures). The surgical repair of most injuries is improving rapidly, owing to advances in microsurgical techniques, but other factors contributing to recovery from peripheral nerve damage, such as nerve protection and regeneration, cannot be improved by surgery. Therefore, ef ective drug therapy is important in the treatment of peripheral nerve injury. This article highlights the role of drug therapy in peripheral nerve regeneration.
The promotion of neuronal repair has always been an important research question in the f eld of nerve regeneration. A large number of drugs have been investigated as potential treatments for peripheral nerve injury, including neurotropic drugs (vitamins), neurotrophic factors, ganglioside, glucocorticoids and herbal extracts. However, although these drugs improve outcome to a certain degree, recovery is often unsatisfactory, and the wide range of underlying molecular mechanisms remain incompletely understood.
Drug ef ects on peripheral nerve regeneration after injury can be divided broadly into two types: (1) a direct ef ect on neurons; (2) indirect neuroprotection and promotion of nerve regeneration via ef ects on glial cells. The former is the focus of earlier studies, and the latter has begun to receive more attention in recent years. Glial cells, previously largely regarded as passive supporting cells of the nervous system, are now understood to play an important role in nerve regeneration and repair. Schwann cells–the only glial cells in the peripheral nervous system–can inf uence, and even determine the entire process of, peripheral nerve regeneration and repair (Frostick et al., 1998; Pearse et al., 2004). Schwann cells can be activated following peripheral nerve injury, and then proliferate, migrate, and form tubes of fusiform cells on the basal lamina. These synthesize and secrete various neurotrophic substances (such as brain-derived neurotrophic factor, nerve growth factor and glial cell line-derived neurotrophic factor) and cell adhesion molecules, and provide support for peripheral nerve regeneration. Schwann cells wrap regenerated axons, form the myelin sheath, and improve recovery of neurophysiological function (Johnson et al., 2005).
Previous studies have shown that the regulatory effect of Schwann cells is an important mechanism underlying the promoting ef ect of drugs on peripheral nerve regeneration (Ma et al., 2010, 2013). After injury, Schwann cells will be subjected to mechanical damage, hypoxia and oxidative stress, resulting in autophagy and apoptosis. The changes in biological behavior and function of Schwann cells will directly af ect nerve regeneration and repair (Figure 1).
Oxidation-reduction (redox) is one possible drug target in Schwann cells. Their high phospholipid content renders the nervous system vulnerable to attack by reactive oxygen species, causing oxidative stress-mediated cellular damage and leading to neurological impairment. Oxidative stress is a key mechanism underlying nerve injury. Antioxidant therapy attracts much attention as a research topic in nervous system diseases and injuries; however, excessive antioxidant levels are also harmful. Redox reactions constitute the most basic biochemistry of the human body, and have important regulatory ef ects. Numerous studies (Li et al., 2012; Sciancalepore et al., 2012; Lu et al., 2013; Kennedy et al., 2013; Yang et al., 2013) have shown that products of redox reactions, such as reactive oxygen species and superoxide, can regulate signaling pathways involving p38 MAPK, ERK1/2, Akt, IGF-1/IGF-1R and JNK, and promote cell proliferation, dif erentiation and migration. Neuronal stem cell proliferation requires a balance of reactive oxygen species, with excessive clearance resulting in impairments in cell proliferation and neuronal dif erentiation. In PC12 cells, the right amount of hydrogen peroxide promotes long-term activation of ERK1/2, ultimately contributing to dif erentiation. Moreover, reactive oxygen species may also enhance the ef ects of nerve growth factor.
Another potential drug target in Schwann cells is the process of autophagy. Schwann cell myelination depends, to a certain extent, on the regulation of autophagy for its morphology and structure, and promoting Schwann cell autophagy can inhibit the development of neuropathic pain. Therefore, autophagy-related regulatory mechanisms in Schwann cells may have an important ef ect on neuronal protection and regeneration after nerve injury; few studies have investigated this to date.
In conclusion, the body faces a variety of pathophysiological stresses after peripheral nerve injury. During the injury process, the biological behavior of Schwann cells and the promotion of peripheral nerve regeneration may be inf uenced by many factors, which can be controlled by drugs. This is a promising and broad-reaching topic of research for future studies concerning drug treatment for peripheral nerve repair.
Jun-xiong Ma, Liang-bi Xiang*
Department of Orthopedics, General Hospital of Shenyang Military Area Command of Chinese PLA, Rescue Center of Severe Wound and Trauma of Chinese PLA, Shenyang, Liaoning Province, China
*Correspondence to: Liang-bi Xiang, xiangliangbi1963@sina.com.
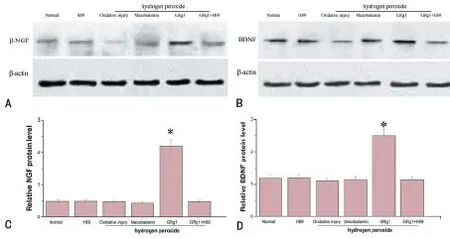
Figure 1 GRg1 could promote synthesis of NGF and BDNF of Schwann cells after hydrogen peroxide-induced oxidative injury.
In vivo transplantation of green f uorescent protein-labeled neural stem cells delays denervation-induced muscle atrophy
This study explored the survival, dif erentiation, and migration of green f uorescent protein (GFP)-labeled embryonic rat neural stem cells (NSCs) after transplantation into the distal injury site of the tibial nerve. We also observed the ef ects of this method on delaying denervation-induced muscle atrophy and investigated the possible involved mechanisms.
GFP-labeled embryonic rat NSCs were isolated, cultured, dif erentiated in vitro, and identif ed using immunof uorescence staining. F344 rat models of tibial nerve transection were successfully established. An in vitro-passaged GFP-NSC single-cell suspension was transplanted into the distal branches of the tibial nerve. At 12 weeks after transplantation, the tibial neve and the musculi triceps surae of the experimental side were collected. Frozen sections of the tibial nerve were analyzed using immunof uorescence staining, and NSC survival, dif erentiation, and migration in vivo were observed under a confocal laser scanning microscope and f uorescence microscope.
A total of 24 healthy, adult, F344, female rats, weighing 180 ± 20 g at 2 months of age, were equally and randomly divided into experimental and control groups. On the rat right thigh, the tibial nerve was horizontally transected at 1.5 cm above the knee. At the proximal end, the nerve was sutured using the end-folded method, thereby establishing animal models of denervated triceps surae. An in vitro-passaged GFP-NSC single-cell suspension was transplanted into distal branches of the transected tibial nerve of F344 rats in the experimental group. The stem cell culture medium was transplanted into the control group using the same method. At 4 and 12 weeks after transplantation, wet weights of the triceps surae was measured. Muscle f ber cross-sectional area, morphology, and area of the postsynaptic membrane were observed and measured using hematoxylin-eosin staining of the muscle, as well as trichrome staining and immunofluorescence staining of the postsynaptic membrane (anti-Synaptophysin for presynaptic membrane and α-BTX for postsynaptic membrane).
Stably passaged GFP-NSCs were obtained by isolation and culture in vitro. The cells were then dif erentiated into neurons, astrocytes, and microglia in vitro. Results showed that GFP-NSCs transplanted into the distal tibial nerve of F344 rats survived, differentiated. Some of the cells dif erentiated into neurons (Figure 1) and extended axonal structures towards the distal end. At 4 and 12 weeks after transplantation, the wet weight of the triceps surae and muscle f ber cross-sectional area were signif cantly greater in the experimental group than in the control group (P < 0.05). At 12 weeks, the postsynaptic membrane was visibly shrunken and the structure did not appear normal in the control group. Morphology of the postsynaptic membrane in the experimental group was near to normal. The area of postsynaptic membrane was signif cantly greater in the experimental group than in the control group (P <0.05).
Experimental results verif ed that GFP-NSCs exhibit good biological activity, and can dif erentiate into neurons after transplantation in vivo. Additionally, in vivo transplantation of GFP-NSCs can delay denervation-induced muscle atrophy, which provides a novel clinical method for prevention and treatment of muscle atrophy following peripheral nerve injury.
Yun-dong Shen, Jian-guang Xu*
Department of Hand Surgery, Huashan Hospital, Fudan University, Shanghai, China (Shen YD, Xu JG)
Department of Hand and Upper Extremity Surgery, Jing’an District Centre Hospital, Shanghai, China (Shen YD)
*Correspondence to: Jian-guang Xu, M.D., Ph.D., xujianguang@hotmail.com.
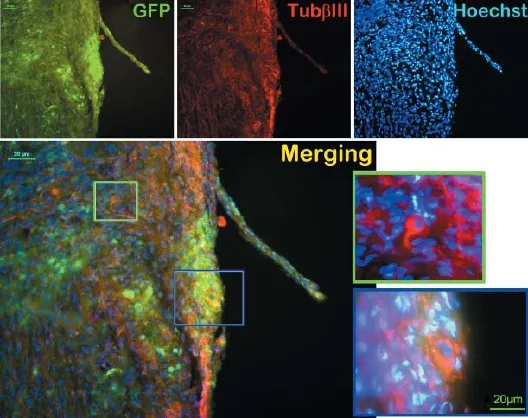
Figure 1 Green f uorescent protein (GFP) labeled neural stem cells transplanted into the transected distal tibial nerve of F344 rats dif erentiate into neurons.
Key issues of stem cell therapy for peripheral nerve injury
Peripheral nerve injury is mainly induced by trauma and causes sensory, motor, and autonomic dysfunction in control regions, and greatly impacts upon quality of life. Traditional treatment methods rely on direct suture and autologous and allogeneic nerve grafts. However, postoperative functional recovery is not satisfactory. Therefore, developing new methods for repair of peripheral nerve injury is an important issue. Stem cell technology has boomed in recent years and provides a novel pathway for treating refractory disease. Mesenchymal stem cells (MSCs) can be easily obtained and proliferated, are not associated with ethical controversy, present low immunogenicity, can be used in allogeneic transplantation, and show good prospect for the treatment of peripheral nerve injury (Jarocha et al., 2015). Nevertheless, many critical issues need to be solved before successful treatment of peripheral nerve injury using stem cells.
First, stem cells are easily af ected by the surrounding environment. Stem cell quality control relates to safety and ef cacy of its clinical application. There are various types, characteristics, preparation methods, and treatment plans for stem cells. As future bio-therapeutic products, stem cell preparation at every stage of research and development must have strict standard operating procedures (including preparation, in vitro testing, animal experiments, clinical research, and treatment) to ensure quality control of stem cell preparations and increased safety and ef cacy of treatment (National Health and Family Planning Commission of China, 2015). Of the numerous stem cell sources, Wharton’s jelly-derived MSCs (WJMSCs) derived from medical waste have high purity and express the main embryonic stem cell markers. Moreover, their differentiation ability is higher than MSCs derived from bone marrow and fat. Increasing numbers of researchers preserve WJMSCs in stem cell pools, so WJMSCs have the potential to provide the basis of large-scale clinical application. Indeed, we obtained abundant WJMSCs with in vitro serial subculture, although WJMSC senescence emerged and oncogene c-Myc expression was upregulated. Furthermore, dif erentiation of WJMSCs into myocardial and nerve cells had undergone signif cant changes. These f ndings suggest that in vitro passaging may cause a signif cant change in stem cell properties, and indicates that early-passage stem cells are needed for future clinical application (Lian et al., 2015). Accordingly, stem cell senescence, and oncogene and dif erentiation ability should be monitored. In addition, the major factors af ecting stem cell properties should be investigated in subsequent in vivo and in vitro tests and clinical studies, with corresponding quality control indices and standards set to improve the safety and ef cacy of stem cell therapy.
Second, increasing the dif erentiation rate of stem cells in vivo contributes to greater safety and ef cacy of stem cell therapy. Most studies have found that only a small number of MSCs dif erentiate into neurons or Schwann cells in vivo. It is presently believed that the MSC repair mechanism is paracrine. The neuroprotective ef ect of stem cells avoids further injury to nerve cells at early-stage injury, and cell replacement ef ects support long-term functional recovery in patients (Shear et al., 2004). Increasing the directed dif erentiation rate of MSCs into nerve cells in vivo may further improve the treatment ef ect, achieve therapeutic ef ciency, and reduce tumorigenic risk after stem cell transplantation. Nevertheless, how to ensure a high survival rate in vivo, and simultaneously further increase the directed dif erentiation rate of MSCs is another key issue that remains to be resolved. If directed pre-dif erentiation of MSCs into nerve cells in vitro can be performed to a certain extent before transplantation (thereby reducing mesodermal dif erentiation ability and appropriately increasing the proportion of nerve cells), it will be useful in the treatment of nervous system disease. Creating an appropriate microenvironment for stem cell dif erentiation in vivo is conducive to maintaining the repair function of stem cell-derived dif erentiated cells (Bhatia, 2010). Thus, we developed a method of directed pre-dif erentiation of stem cells induced by cell co-culture. That is, nerve cells grown in a three-dimensional environment were used to simulate in vivo nerves to ef ciently induce MSC dif erentiation into nerve cells. This system promotes directed induction of stem cells, achieves complete separation of target and induced cells, ensures application security (Liu et al., 2013), and provides a new means for increasing stem cell function and reducing clinical risk.
Third, future large-scale clinical research and application requires many dif erentiated cells derived from stem cells. Culture f asks and culture plates are dif cult to meet these clinical needs, so it is necessary to develop a large-scale dif erentiation induction system based on a bioreactor. On the one hand, f uid f ow in bioreactors increases the material exchange rate of the system and raises dissolved oxygen levels. While by coupling to a three-dimensional culture carrier, the specif c surface area of the bioreactor system is far larger than an ordinary two-dimensional culture system, and is suitable for large-scale cell culture. According to the characteristics of stem cell dif erentiation, we constructed a three-dimensional microenvironment to induce directed dif erentiation of stem cells using a three-dimensional biomimetic scaf old, microcapsules, magnetic particles, and rotary bioreactors (Figure 1). Specif cally, a modif ed three-dimensional biomimetic scaf old provides an extracellular matrix microenvironment for stem cells that is similar to in vivo nervous tissue. Magnetic particles remove induced cells in microcapsules. The rotary bioreactors increase mass transfer in the system and provide a three-dimensional microgravity environment for the cells. Development of this system is a key approach for reinforcing pre-dif erentiation of stem cells in vitro and producing functional cells.
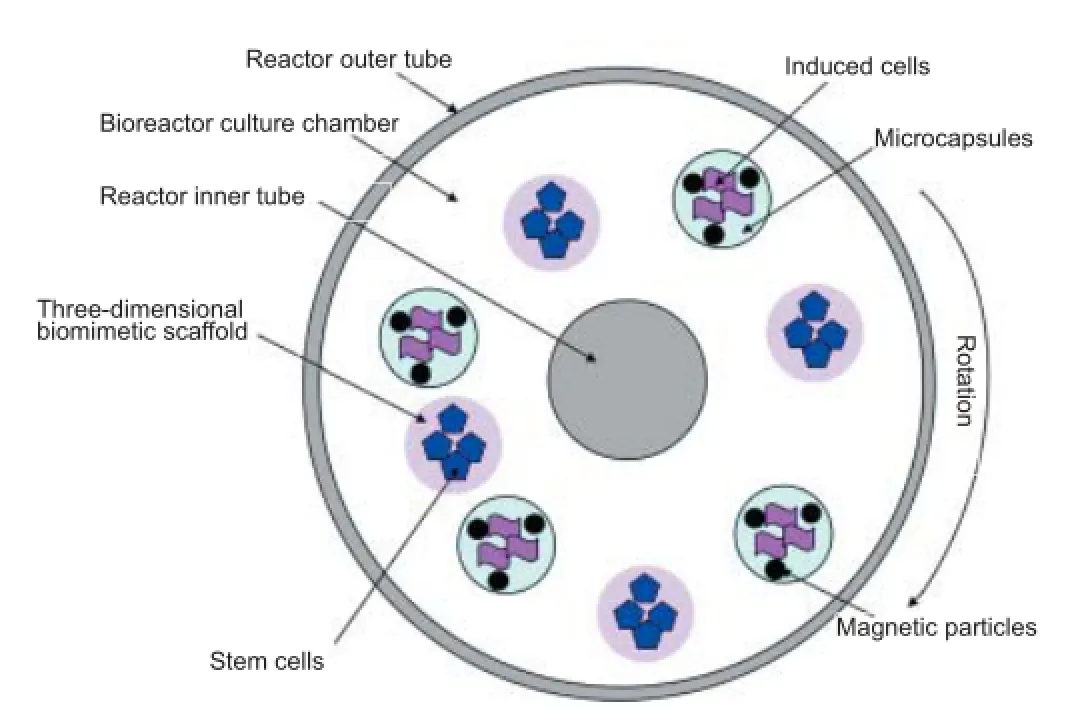
Figure 1 Three-dimensional microenvironment for inducing directed dif erentiation of stem cells.
In summary, along with the development of science and technology, the problems that restrict the clinical application of stem cells will gradually be resolved. It is believed that stem cells will play an increasing role in the repair of peripheral nerve injury, and bring more hope to people.
This study was supported by the National Natural Science Foundation of China, No. 31271055, 31470944, 20906088.
Yang Liu, Guang-wei Sun*
Laboratory of Biotechnology, Dalian Institute of Chemical Physics, Chinese Academy of Sciences, Dalian, Liaoning Province, China
*Correspondence to: Guang-wei Sun, M.D., sungw@dicp.ac.cn.
Repair, regeneration and remodeling of injured peripheral nerves
Data from the World Health Organization (WHO) revealed that approximately a quarter of the world’s population suffers from varying degrees of accidental trauma. The most common and severe painful injury is nerve damage-induced limb disability and dysfunction. Approximately 20 million patients are af ected by peripheral nerve injury, with approximately two million new cases per year. The ef ective repair of peripheral nerve injury and methods of promoting this are major public health concerns in China. In the following sections we will discuss recent studies and the areas of focus on regeneration following peripheral nerve injury.
Neuroprotective mechanisms: Neuroprotection is always a key area of research for preserving function following peripheral nerve injury. Neurons from spinal cord and dorsal root ganglion undergo a series of changes in morphology, metabolism, biochemistry and gene expression, as well as the induction of apoptosis after peripheral nerve injury. Oxidative stress also occurs in neuronal cells after peripheral nerve injury. Intracellular calcium overload and reactive oxygen species interfere with mitochondrial function, and activate p53, Bax and caspase 3 pathways, resulting in the induction of apoptosis. Following peripheral nerve injury, neurons undergo a protective response, increasing the gene expression of neurotrophic factors, which in turn promote neuronal protection and survival. Excitotoxicity of action potentials and the disruption of axonal transport at the injury site are key factors that activate neurons. The administration of exogenous neurotrophic factors might be an important strategy for inhibiting neuronal apoptosis. Various researchers have verif ed the protective ef ects of the biological conduit small gap sleeve bridging method for nerve injury on the local microenvironment. In addition, complementary ef ects of combination strategies incorporating multichannel channels conduits with nerve growth factors, hormones or Chinese medicinal compounds have been observed. However, the mechanism by which neurons induce self-protective responses following peripheral nerve injury remains unclear and deserves further investigation. Understanding neuroprotective mechanisms may help identify more ef ective methods to inhibit neuronal apoptosis and elevate repair ef ects after peripheral nerve injury.
Repair methods following peripheral nerve injury: Conventional repair methods for peripheral nerve injury involve the use of epineurial or perineurial sutures to reconnect the damaged ends; however, these methods cannot achieve ef ective reinnervation of dif erent nerve f bers in injured neurons. Numerous researchers have proposed many methods to achieve ef ective reinnervation of sensory and motor f bers. For example, the reinnervation of nerve f bers can be controlled by connecting to the capillary network of the injured epineurium, while sensory f bers and motor f bers can be distinguished by immunohistochemical staining of intraoperative frozen sections. However, these methods have their own limitations. The capillary network can be damaged after nerve injury, and intraoperative pathological staining is time-consuming. Zhang et al. (2013) proposed biodegradable conduit small gap tubulization for the repair of peripheral nerve injury, which has been verif ed in Sprague-Dawley rats and rhesus macaques. Recently, a multi-center human clinical trial was completed, in which the biological conduit small gap sleeve bridging method for peripheral nerve injury ef ectively promoted the ef ective reinnervation of dif erent f bers in the proximal and distal ends of the conduit, perfectly restoring the function of distal target organs, and reducing the occurrence of painful neuromas (Zhang et al., 2013). The mechanisms proposed for achieving these ef ects were proposed as follows: (1) the biological conduit-constructed small gap fully permits selective regeneration; (2) it reduces the spillover of regenerated nerve f -bers; (3) the constructed regeneration chamber protects the local microenvironment and is conducive to the role of neurotrophic factors; (4) or the regeneration chamber provides the possibility and feasibility of repairing thick nerves using thin nerves (Jiang et al., 2010). Compared with the epineurial suture, the biological conduit small gap sleeve bridging method is a technological innovation for the repair of peripheral nerve injury. The clinical trial also found a number of limitations that cannot be explained by the traditional theory of peripheral nerve regeneration: (1) An enlarged compensation of the number of nerve f bers occurred during peripheral nerve regeneration: when donor never f bers in the proximal end were used at ratios of 1/1, 2/1, 4/1, and 8/1 to repair distal nerve f bers, the nerve regeneration magnif cation ratio (the ratio of the number of distal nerve f bers to proximal nerve fibers) were 0.98:1, 1.51:1, 2.39:1 and 2.89:1, respectively. The maximum magnif cation ratio was 3.3:1 after Hill function curve f tting. (2) During peripheral nerve repair, a donor nerve is used to bridge the distal and proximal ends of a damaged nerve to help regain control of the distal nerve-innervated antagonistic muscle. The functions of above two nerves restore. The above phenomena have attracted great attention from experts and caused them to rethink their theoretical beliefs on nerve regeneration and repair on the whole part.
Treatment of nerve defects: The treatment of nerve defects, especially long-segment nerve defects, is a key area of clinical and basic research. Many researchers have investigated various neurological substitute materials, including artif cial peripheral nerves, nerve allografts, and artif cial nerves incorporated with a wide variety of bioactive substances. Studies examining tissue-engineered artif cial nerve replacement of nerve defects in China have always been at the forefront of the world. Many institutes in China perform studies using bionic microtubule-like ultrastructural nerve scaf olds. Substitutes for nerve grafts, such as nerve canal bridges and artif cial nerves have been used in clinical applications. Similar progress in the fabrication of artif cial peripheral nerves has been achieved in the USA, Germany, Japan and China. The main dif erence in artif cial peripheral nerves fabricated to date lies in their biocompatibility. Thus, it is necessary to examine their biocompatibility, to prepare artif cial peripheral nerve scaf olds that are compatible with human physiological requirements and bodily f uids.
Promoting peripheral nerve regeneration: Even if sophisticated microsurgical techniques are applied for peripheral nerve injury, functional recovery remains unsatisfactory. A key problem in the f eld of nerve injury is improving the rate of axon regeneration and accelerating nerve regeneration after injury. Researchers outside of China have used direct current electric f elds, electric pulse stimulation, ultrashort wave and hyperbaric oxygen to promote peripheral nerve regeneration, with the ef ects of these being variable. Chinese researchers have investigated mechanisms of activating neuronal growth, improving the microenvironment for axonal regeneration and myelination following nerve injury, as well as regulating genes that promote neurite extension and myelination. To date, a number of studies have investigated genesresponsible for myelination, including Krox20 (EGR2), Sox2, Sox10, Oct6 (SCIP/Tst1), NFКB (nuclear factor-κB), SREBP (sterol regulatory element-binding protein) and JUN (c-Jun); however, the underlying molecular mechanism of myelination remains unclear.
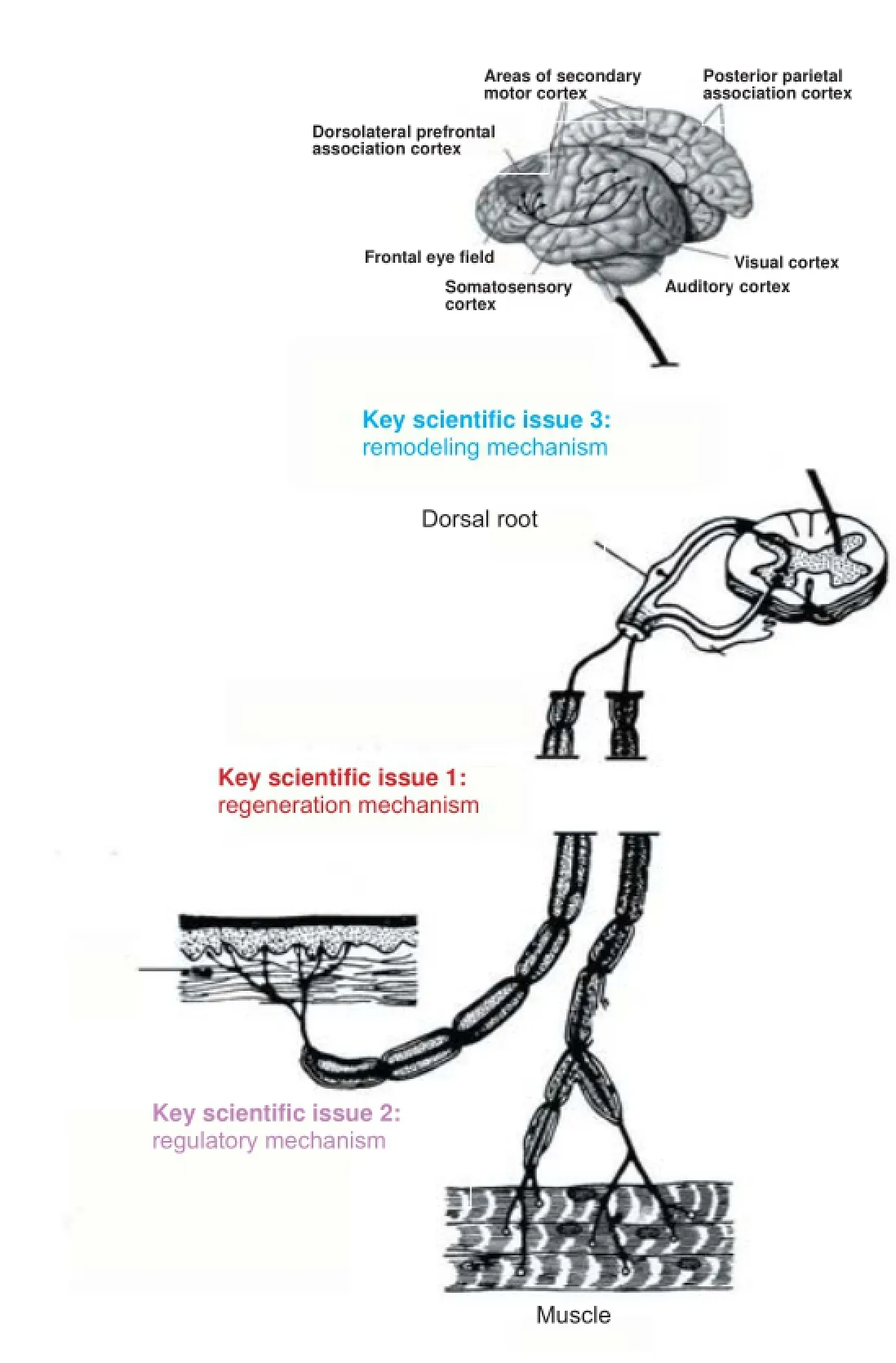
Figure 1 Process of nerve injury, repair and regeneration in the hypothesis of “Neural regeneration after peripheral nerve injury repair is a system remodeling process of interaction between nerves and terminal ef ector”.
Inhibiting denervation atrophy of muscle: Peripheral nerve injury repair, especially following brachial plexus injury, has been a major focus of scientif c research and clinical studies. Researchers have explored many methods, including nerve transposition or grafting, but their reparative ef ects have been limited to date. The speed of axonal growth is slow after repair, and following brachial plexus injury, the distal end, which is responsible for hand function, contracts when proximal axons grow into the distal end of the upper limb. Inf ammation, motor endplate degeneration, muscle atrophy, apoptosis and proliferation of muscle satellite cells all occur after denervation. In aseptic inf ammation induced by cytotoxicity, inf ammatory factors secreted from various inf ltrating inf ammatory cells, such as tumor necrosis factor-α (TNF-α), interleukin (IL)-1 and IL-6, trigger the process of skeletal muscle mass loss. Further study is required to determine the most ef ective mechanism for promoting faster proximal nerve regeneration and to protect against delayed motor endplate degeneration of distal target organs.
The 973 Program of the State Ministry of Science and Technology of China, entitled “Mechanisms of neural regeneration and central nerve remodeling after peripheral nerve injury repair”, were f rst to propose the hypothesis that neural regeneration after peripheral nerve injury repair is a system remodeling process that involves interaction between nerves and terminal ef ectors (Zhang et al., 2015). Their hypothesis addresses the entire process of nerve injury, repair and regeneration, and focuses on three important aspects af ecting nerve repair, as follows: the damaged nerve and postoperative regenerative mechanisms; end-ef ector and pain-related regulatory mechanisms; and ef ector-induced remodeling of the nerve (Figure 1). We look forward to new insights into the regenerative mechanism of peripheral nerve following injury, and hope that a new scientif c method for clinical treatment of this disorder will be developed.
This study was funded by the Chinese National Ministry of Science and Technology 973 Project, No. 2014CB542201 and 863 Project, No. SS2015AA020501; the Ministry of Education Innovation Team (IRT1201), the National Natural Science Fund, No. 31571235, 31371210, 31271284, 31171150; the Educational Ministry New Century Excellent Talents Support Project, No. BMU20110270.
Pei-xun Zhang*, Na Han*, Bao-guo Jiang*Peking University People’s Hospital, Beijing, China
*Correspondence to: Pei-xun Zhang, Na Han, Ph.D. or Bao-guo Jiang, Ph.D., M.D., hannaqa@hotmail.com or jiangbaoguo@vip.sina.com.
Molecular mechanism of the regulation of neuron-intrinsic regeneration
Adult neurons in the peripheral nervous system have a certain capacity for regeneration. Fully understanding the mechanism of peripheral nerve regeneration will hopefully help address limitations encountered in central nervous system regeneration. This paper discussed key issues in nerve repair and regeneration focusing on transcriptional regulatory factors, key regulatory proteins, non-coding RNAs, and epigenetic regulation of acetylation.
In models of peripheral sciatic nerve injury, transcription factors related to regeneration-associated gene transcription in dorsal root ganglia include ATF3 (activating transcription factor 3), CREB (cAMP response element-binding protein), STAT3 (Signal transducer and activator of transcription 3), JUN (c-Jun), Sox11, SMAD1, p53, NFIL3 (nuclear factor, interleukin 3 regulated), C/EBPbeta (CCAAT/enhancer binding protein), C/EBPdelta, NFКB (nuclear factor-κB), NFAT (Nuclear factor of activated T-cells), and KLF4 (Kruppel-like factor 4), KLF7 (Ma and Willis, 2015). Seven classical regulatory pathways are involved, as follows: the cAMP/PKA/CREB; JNK/c-JUN; ATF3; JAK/STAT3; CBP/p300/PCAF-p53; KLF4; and BMP4/Smad1 pathways. There are links between these pathways, including: c-Jun may act together with ATF3 and STAT3 to induce peripheral nerve regeneration; KLF4 may cooperate with p53 to transactivate the enhancer of p21Cip1/Waf1 and to in turn af ect nerve growth; and KLF4 can bind to the tyrosine 705 phosphorylation site of STAT3 to inhibit the regeneration of the axons of the optic nerve.
Park et al. (2008) found that PTEN (phosphatase and tensin homolog) knockout not only reduced apoptosis, but also promoted axonal regeneration. Furthermore, rapamycin could suppress axonal regeneration in the PTEN knockout model (Lu et al., 2014), suggesting the mammalian target of rapamycin (mTOR) signaling pathway was essential for axonal regeneration. However, the mTOR pathway was not solely responsible for axonal regeneration in the PTEN knockout model, but glycogen synthase kinase 3 also participated in the process (Park et al., 2008). Further research is required to identify the exact mechanism responsible for axonal regeneration in the PTEN knockout. If mTOR activity is maintained by PTEN knockout in cortical neurons, axonal regeneration of the corticospinal tract could be greatly promoted (Lu et al., 2014). In dorsal root ganglia, PTEN or TSC2 (tuberous sclerosis complex 2) deletion can promote axon growth and regeneration. These results indicate that PTEN/ mTOR signaling plays an important role in axonal regeneration, with or without activating ribosomal protein S6 kinase.
Suppressor of cytokine signaling (SOCS) proteins inhibit signal transduction and transcriptional activation by binding to Janus kinase (JAK) or cell receptor tyrosine residues. Of the SOCS proteins, SOCS3 is extensively expressed in the nervous system. SOCS3 knockout in retinal ganglion cells has been shown to promote neuronal survival and axonal regeneration after optic nerve injury. Furthermore, gp130 knockout obviously suppressed axonal regeneration in SOCS3 knockout mice, suggesting that axonal regeneration in the SOCS3 knockout is regulated by gp130 signaling. In retinal ganglion cells, SOCS3 overexpression by adenoviral transfection almost completely suppressed axonal regeneration. The ef ects of injecting recombinant human ciliary neurotrophic factor (CNTF) into the vitreous body can also be inhibited by SOCS3 overexpression, indicating that SOCS3 overexpression can suppress cell survival.
In retinal ganglion cells, SOCS3 knockout upregulated mTOR activity post-nerve injury, and contributed to the response of retinal ganglion cells to the damage factor. Lu et al. (2014) proposed that PTEN and SOCS3 synergistically promote axonal regeneration through two independent pathways.
Non-coding RNA, including microRNA (miRNA) and long non-coding RNA (lncRNA), constitutes a complex regulatory network. miRNA participates in cell apoptosis, proliferation, migration, differentiation, development and metabolism. lncRNA are RNA molecules > 200 bp long that were initially believed to be “noise” of genome transcription because they did not encode proteins. Nevertheless, recent studies showed that lncRNA is involved in chromatin modification, transcriptional activation, interference and nuclear transport. We analyzed changes in miRNA expression prof les in L4–6dorsal root ganglia after sciatic nerve injury, and found that miR-21 and miR-222 suppressed the apoptosis of dorsal root ganglion neurons by targeting tissue inhibitor of metalloproteinases-3 (TIMP3). Many current studies are focused on miRNA ef ects on the growth of neuronal processes. For example, miR-21 was rapidly upregulated after dorsal root ganglion neuronal injury, where it promoted the regeneration of neuronal processes by targeting Sprouty2. miR-222, miR-431, miR-145, miR-138, miR-214 and miR-132 regulated axonal regeneration by targeting PTEN, kringle containing transmembrane protein 1 (Kremen1), roundabout guidance receptor 2 (Robo2), Sirtuin type 1 (SIRT1), Slit-Robo GTPase-activating protein 3 (srGAP3) and Ras GTPase activator 1 (Rasa1). Recent studies conf rmed that 24 lncRNAs were downregulated in dorsal root ganglion after sciatic nerve injury. Bioinformatics analysis verified that possible target genes of lncRNA were associated with the MAPK pathway. Gu et al. (2014) hypothesized that interference of lncRNA BC089918 and uc.217 could contribute to the growth of neuronal processes in dorsal root ganglion. These studies described the molecular regulation of nerve regeneration from a new perspective.
Genetic studies on acetylation have mainly focused on the ef ect of histone acetylation, the most common form, on chromosomal structure and the activation of nuclear transcription factors. Recent studies showed that histone deacetylase 5 (HDAC5) nuclear export is a key step in axonal regeneration of dorsal root ganglia. HDAC5 nuclear export induces a series of gene expression changes that promote axonal regeneration. Simultaneously, HDAC5 also can move to the injury site to assist in the formation of microtubules, which contribute to axonal regeneration. However, nerve cells in the cerebrum and spinal cord lack HDAC5 nuclear export, which can partially explain why axonal regeneration in the central nervous system is dif cult (Cho et al., 2013).
The activation of neuronal intrinsic regeneration is controlled by a series of extracellular signals associated with the regenerative capacity of neurons, mainly involving transcriptional regulatory factors, key regulatory proteins, non-coding RNAs and the epigenetic regulation of acetylation. Historically, the major limiting factor in the study of neuronal intrinsic regeneration was the measurement of neurite length in vitro compared to those that are damaged in vivo. However, the establishment and optimization of microf uidic cell culture and the “spot” culture method have largely addressed this limitation. Another limitation is the ef ective delivery of drugs to target neurons. Recently developed methods of intrathecal injection of siRNA and combined injection of siRNA and miRNA in a peripheral nerve regeneration chamber have achieved good results. The establishment of these methods widely broadens the f eld of neuronal intrinsic regeneration, and deepens the understanding of the underlying regulatory mechanisms of nerve regeneration. A greater understanding has helped develop new strategies for treating nervous system diseases, and provides useful scientif c guidance for clinical application.
This work was supported by the National Natural Science Foundation of China, No. 31200799 and 81571198; the New Century Excellent Talents in University, No. NCET-12-0742; and the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
Song-lin Zhou, Bin Yu*
Jiangsu Key Laboratory of Neuroregeneration, Co-innovation Center of Neuroregeneration, Nantong University, Nantong, Jiangsu Province, China
*Correspondence to: Bin Yu, yubin@ntu.edu.cn.
Neurotropism in peripheral nerve regeneration
The peripheral nervous system is essential for the functioning of the limbs and organs. Most peripheral nerves comprise a mixture of sensory, motor and autonomic neurons. The main function of peripheral nerves is to carry information obtained from the peripheral ef ector to the central nervous system, and to transmit the instruction issued by the central nervous system to the end ef ector to produce physiological activity. In cases of peripheral nerve rupture, the pathway between the central nervous system and target organ is interrupted, and Wallerian degeneration, a series of changes in cell and molecular biology, occurs in the distal segment of the nerve (Li et al., 2013). The proximal nerve has good regenerative capacity, but disruption to the original pathway may cause regenerated f bers to be redirected to incorrect target organs, leading to the wrong muscles being controlled by the motor nerve f bers and resulting in motor dysfunction. Similarly, incorrectly regenerated sensory nerve f bers will cause sensory abnormalities. Miswiring in the autonomic nervous system can lead to autonomic dysfunction due to mislocalization, and neurotropism is the key to preventing mislocalization.
The concept of neurotropism was f rst proposed by Forssman in 1898, and improved by Cajal in 1928 (Ramon et al., 1959; Weiss et al., 1944). During the growth and development of a nerve, new axons are guided in the direction of chemicals released from peripheral tissues. Neurotropism is classif ed into tissue specif city, topographic specif city and end-organ specif city. In tissue specif city, the regenerated nerve grows only towards the distal nerve, not other tissues. Topographic specif city means that several regenerated axons grow towards the same nerve in the distal site. In end-organ specif city, motor and sensory axons grow into the corresponding endoneurial tube at the distal site. At present, various methods have been used to repair peripheral nerve injury, such as autologous nerve transplantation, nerve allografts and nerve conduits. Neurotropism provides a new target for improving the docking precision of regenerating nerves. However, two problems deserve our consideration: the physical basis of neurotropism, and whether neurotropism can be achieved under injury conditions.
Neurotropism in peripheral nerve regeneration depends on the distance between the nerve stumps, the distal nerve volume, nutritional factors, and neural structures. The nerve stump distance must fall within a limited range, at which the distal nerve and the target organ exert chemotactic ef ects on the regeneration of the proximal nerve. Too big or too small a distance is not conducive to chemotaxis; if the distance between the stumps is too small, proximal axons can grow to the distal stump without the aid of chemotactic factors, but if it is too large, the nutritional factors and chemokines released from the distal stump and target organ cannot achieve an ef ective concentration at the nerve stump. Lundborg et al. (1982) reported that the distance between nerve stumps should be 6–10 mm for optimal chemotaxis. Following peripheral nerve rupture, the route between the central nervous system and target organ is disrupted, and a series of changes in cell and molecular biology occur in the distal segment, including the disintegration and removal of axons and myelin sheath. Schwann cells proliferate extensively and secrete various factors for nerve regeneration. Chemotaxis is inf uenced by the size of the distal nerve, with larger volumes at the distal site being more conducive to regeneration of the proximal nerve (Iwabuchi et al., 1999; Takahashi et al., 1999). Neurotrophic factors and extracellular matrix proteins play an important role in neurotropism (Hari et al., 2004; Naidu, 2009). Höke et al. (2006) showed that neurotropism has phenotypic specif city, meaning that Schwann cells in motor and sensory nerves have dif erent phenotypes, secrete dif erent inducers, and exert dif erent ef ects during nerve growth. Maness and Schachner (2007) suggested that recognition molecules on the surface of axons form the basis of neurotropism. At present, neurotropism is recognized by experts in China and worldwide, but its precise mechanism remains unclear.
The repair of peripheral nerve damage remains an unresolved medical problem, especially with long-segment defects. A number of methods exist for the treatment of peripheral nerve injury, with autologous nerve transplantation being the “gold standard”. However, its clinical application is limited because of the need for more than one surgical procedure, risking new trauma, and donors are limited. Improvements in acellular techniques and immune elimination have brought new hope for the treatment of peripheral nerve injury, but nerve allografts still have some disadvantages, such as a limited source and risk of infection. Progress has been made in the design of various peripheral nerve repair conduits. These provide a good three-dimensional bionic space for nerve regeneration and contribute ef ectively to neurotropism It is found that the ef ects of alpha-cyanoacrylate adhesives combined with silicone tube are identical to that of epineurial suture in restoring damaged nerve morphology (Figure 1) and function in rats with sciatic nerve injury. Moreover, the operation time is short, showing the potential of its clinical application. It may require additional time to investigate how to ef ectively promote peripheral nerve regeneration during repair so as to successfully apply in the clinic. Our focus for the future is to identify the most ef ective and reliable method of repairing peripheral nerve damage. With continued improvements in medical technology, and the understanding of peripheral nerve anatomy and the role of neurotropism in nerve regeneration, we believe that we can f nd a better way to treat peripheral nerve injury.

Figure 1 Toluidine blue staining results of alpha-cyanoacrylate adhesive and silicone tube repair (A) and epineurial suture (B) on the anastomotic stoma of sciatic nerve after distal transection (× 100)
Li-cheng Zhang, Jian-heng Liu, Li-hai Zhang, Pei-fu Tang*
Department of Orthopedics, General Hospital of People’s Liberation Army, Beijing, China
*Correspondence to: Pei-fu Tang, pftang301@126.com.
Intervention, repair, and protection of nerve injury in carpal tunnel syndrome
Carpal tunnel syndrome is induced by median nerve compression at the wrist. Neuropsychological tests are often helpful for diagnosing the disease. During electrophysiological diagnosis of carpal tunnel syndrome, distal latencies of compound muscle action potentials of the median nerve and abductor pollicis brevis are important diagnostic indicators of the disease stage and choice of treatment. Paget (1853) f rst described median nerve compression at the wrist induced dysfunction in the corresponding innervated area. Phalen (1966) established principles for diagnosing and treating carpal tunnel syndrome. Several recent research advances have been made describing the pathophysiology and electrophysiology of carpal tunnel syndrome.
Although the disorder is spontaneously generated in most patients with carpal tunnel syndrome, the specif c cause remains unclear. Fernández-de-las-Peñas et al. (2013) reported that the Val158Met polymorphism may af ect the degree of pain and function in patients with carpal tunnel syndrome, but is not associated with the occurrence of carpal tunnel syndrome. Local factors, such as gout, cancer, an abnormal lumbricalis (Wang et al., 2014), wrist fracture, trauma, and the long-term use of vibrating tools can lead to carpal tunnel syndrome. Systemic factors, such as diabetic neuropathy, pregnancy, and thyroid dysfunction, can also result in carpal tunnel syndrome. Because the space in the carpal tunnel is very limited, any swelling induced by tendon sheath synovitis can compress the median nerve and lead to corresponding symptoms.
Median nerve compressed by increases in the content of the carpal tunnel or decrease in the size of the carpal tunnel volume gradually leads to nerve conduction abnormalities or functional impairment. Nerve microcirculation disturbance occurs after median nerve compression and stretch, which can result in changes to the myelin sheath and axon. Changes in the median nerve are clinically divided into early, middle, and late stages based on the anatomy. During the early stage, clinical symptoms occur intermittently, commonly at night. Disease induction is mainly associated with a redistribution of blood f ow and muscle pump failure in the supine position. If the pressure exceeds 40–50 mmHg, the oxygen supply to the nerve microcirculation is reduced and venous stasis and penetration appear, f nally resulting in endoneurial edema that af ects axonal transport. During the middle stage, numbness can be observed in the area innervated by the median nerve during the day or night. The microcirculation is abnormal, f uid pressure increases, the myelin sheath and the nodes of Ranvier are destroyed, and electrophysiological abnormalities become measurable. During the late stage, the clinical symptoms persist, sensorimotor disorders appear, axonal injury worsens, and Wallerian degeneration occurs.
Electrophysiological examination is an important tool for diagnosing carpal tunnel syndrome. During the early stage of carpal tunnel syndrome, the sensory component of the median nerve is involved and the sensory conduction velocity decreases. For patients with normal sensory conduction velocity, measuring the motor nerve conduction velocity can improve the diagnostic positive detection rate. Gu et al. (2004) reported standards for electrophysiological staging of carpal tunnel syndrome. Middle and early stages: a negative electromyogram (EMG) in the abductor pollicis brevis; distal motor latency of the median nerve < 4.5 ms; and a sensory potential latency dif erence in the ring f nger ≥ 4 ms or, among f ngers 1–3, a decreased SNAP amplitude of at least one f nger exceeding 1/2 of the uninjured side. Middle stage: a positive or negative EMG (±); distal motor latency ≥ 4.5 ms; elicited sensory potentials of f ngers 1–3, but with velocity < 40.0 ms. Late stage: a positive EMG (+); noticeably prolonged or absent distal motor latency; and among f ngers 1–4, a loss of sensory potential of at least one f nger.
High-frequency ultrasound has been a powerful ancillary facility in the carpal tunnel syndrome diagnosing owing to its convenience and low cost. Dif use or localized swelling and echo reduction usually interpret the median nerve pathology under high-frequency ultrasound. Apparent broadening of the median nerve proximal to the transverse carpal ligament is one specif c change that occurs in carpal tunnel syndrome. However, ultrasound is inaccurate in distinguishing delicate connective tissue pathology, and its echo interpretation is expert-dependent. Magnetic resonance imaging (MRI) can reveal the structural changes in the carpal tunnel and identify pathological changes. Determination of the pressure in the carpal tunnel can diagnose or determine whether there is thorough decompression. Pressures within the carpal tunnel > 2 kPa in the rest position and > 18 kPa during active wrist f exion can be used as a basis for the diagnosis of carpal tunnel syndrome. Pressures within the carpal tunnel after surgery of < 0.7 kPa in the rest position and< 2 kPa during active f exion indicate complete surgical release.
The methods for the intervention, repair, and protection of nerve injury in carpal tunnel syndrome mainly include both conservative and surgical treatments. Conservative treatments include rest, bracing, replacement of occupational work, physiotherapy, and penetration of glucocorticoid and osteopathic manipulation. Surgical treatments include traditional surgical incision, small incision, and endoscopic therapy. Ginanneschi et al. (2014) found that the local injection of steroids increased the electric threshold of the Aβ f ber in the median nerve and improved clinical symptoms, leading to their present extensive use. At the proximal end of the carpal tunnel, a thickened and hardened transverse carpal ligament or antebrachial fascia can also cause entrapment. Therefore, forearm fascia release should be considered during surgery, especially through a small incision at the wrist or endoscopy.
Preoperative nerve conduction is associated with postoperative functional recovery. Understanding the preoperative electrophysiological conditions of the median nerve can help the clinician assess the prognosis to achieve targeted therapies. Yang et al. (2006) verif ed that postoperative grip strength was stronger in the latency< 4.0 ms group and latency 4.0–6.0 ms group than in the latency >6.0 ms group. The latency period ref ects the conduction velocity of the action potentials. Nerve demyelination or changes in axon diameter and external compression af ect nerve conduction velocity. An intraoperative latency time of the median nerve > 6.0 ms ref ects severe median nerve injury, which requires a long recovery time and attaining normalcy is dif cult.
If compression is released during the early stage, carpal tunnel syndrome can be quickly improved. If released during the middle stage, carpal tunnel syndrome can still be relieved, but the nerve microcirculation may need to be reconstructed, and the repair of myelin sheath will require several weeks to several months. If compression is released during the late stage, the recovery of nerve function will rely on nerve regeneration overall several months or the neurological function may not be completely recovered. These outcomes indicate the importance of early treatment of carpal tunnel syndrome.
This study was supported by the Key Talent Fund Project of “Science Education for Health” Engineering of Health Department of Jiangsu Province of China, No. RC2011101.
Gu-heng Wang, Ren-guo Xie*
Department of Hand Surgery, Af liated Hospital of Nantong University, Nantong, Jiangsu Province, China (Wang GH)
Trauma Center, Department of Orthopedic Surgery, Shanghai First People’s Hospital, Shanghai Jiao Tong University School of Medicine, Shanghai, China (Xie RG)
*Correspondence to: Ren-guo Xie, M.D., Ph.D., xrg1969@yahoo.com.
Signif cance and strategy of promoting seed cell survival in tissue-engineered artif cial nerve construction
Peripheral nerve injury (PNI) is one of the most common clinical diseases, af ecting 2.8% of all trauma patients. Almost half PNI result in nerve defect with an extensive gap which must be bridged with permissive natural or artif cial grafts. In recent decades, tissue-engineered artif cial nerve graft was widely studied and accepted a permissive strategy for repairing the peripheral nerve defect. Typical tissue engineering graft includes biomaterial scaf old, growth and dif erentiation factors, and seed cells. For nerve tissue engineering, Schwann cells and kinds of stem cells can be used as seed cells. The seed cells are prospected to secrete various bioactive molecules to enhance the axonal regeneration, as well as dif erentiate to remyelinate the regenerated axons. However, the poor survival of seed cells remains a challenge during artif cial nerve construction. To reveal the current progress of seed cells application in artif cial nerve, we reviewed 841 articles concerning tissue-engineered artif cial grafting for nerve defects using PubMed, Google Scholar and Endnote. All articles were in the Science Citation Index and published between 1984 and 2014. Only 76 (9.0%) of the articles reported that seed cells were added to artif cial nerve grafts. Of these, only eight (10.5%) reported survival of the seed cells. That is to say, more than 90% of the studies on artif cial nerves did not use seed cells, and approximately 90% of the studies using seed cells did not mention cell survival. We propose that the most likely reason for this is the low survival rate of seed cells in artif cial nerve. In our previous experimental studies, we also found the same problem that the survival rates of tissue-engineered Schwann cells or other stem cells in the brain, spinal cord and other tissues were higher than in artif cial nerves. Therefore, further studies should be carried out to determine why the survival rate of seed cells is low in artif cial nerves, to seek new strategies to elevate the survival rate, and to strengthen the ef ects of tissue-engineered artif cial nerves in the repair of peripheral nerve defects.
In accordance with our literature review and prior study results, we believe that three factors deserve consideration: (1) Histocompatibility of scaf olds. The most commonly used scaf olds at present are polymer compounds, whose histocompatibility is mainly detected by in vitro cell culture experiments. However, the culture medium used in vitro is abundant and frequently replaced, diluting the substances that maybe harmful for cell growth. We recommend that in vivo experiments have to be used to draw relevant conclusions. Furthermore, we consider that the physical and chemical properties of ideal scaf olds should be close to the natural extracellular matrix, or that at least a layer of this matrix should be coated on the surface of polymer materials. For example, the self-assembling peptide nanof ber scaf old (SAPNS), a gel-like nanomaterial, shows good tissue and cell compatibility in vivo and in vitro. After SAPNS was f lled within a conduit of poly(lactic-co-glycolic acid) or artery, it could signif cantly promote cell survival and migration, and subsequent nerve regeneration (Zhan et al., 2013; Wang et al., 2014) (Figure 1). (2) Immunogenicity of seed cells. Whenever possible, autologous cells should be used to minimize the ef ect of immune rejection on seed cell survival. With the development of induced pluripotent stem cell technique, autologous cells can retrodif erentiate into pluripotent stem cells, or be induced to dif erentiate into Schwann cells. Recently, we isolated a kind of stem cells from peripheral blood, which have similar immunophenotypings and multipotential characters with that of bone marrow or adipose tissue derived mesenchymal stem cells. These cells were named as peripheral blood mesenchymal stem cells (PBMSCs) (Dai et al., 2010; Wu et al., 2015). Under certain conditions, PBMSCs can differentiate into Schwann cells, providing possibilities for autologous cell transplantation (Dai et al., 2010; Wu et al., 2015). (3) Vascularization of artif cial nerve. Peripheral nerve is frequently distributed among intermuscular septum where connective tissue and capillaries are underdeveloped. As the angiogenesis is slow after the artif cial nerve was transplanted, seed cells in artif cial nerve are vulnerable to ischemic and hypoxic damage, and metabolites are dif cult to remove. So transplanted seed cells might die before the vascularization of artif cial nerve. This may be the most important factor in the poor survival of seed cells in artif cial nerves. We are currently exploring methods to accelerate vascularization after grafting, to promote the survival of seed cells, and to improve the contribution of artif cial nerves to the repair of peripheral nerve defects.
This study was funded by Chinese National Ministry of Science and Technology 973 Project, No. 2014CB542202; Natural Science Foundation of China, No. 81371354, 81571182; Natural Science Foundation of Guangdong Province, No. S2013010014697; and Science and Technology Foundation of Guangdong Province, No. 2015A020212024.
Xiang-hai Wang, Jia-song Guo*
Department of Histology and Embryology, Southern Medical University, Guangzhou, Guangdong Province, China (Wang XH, Guo JS)
Key Laboratory of Tissue Construction and Detection of Guangdong Province, Guangzhou, Guangdong Province, China (Guo JS)
Institute of Bone Biology, Academy of Orthopedics, Guangzhou, Guangdong Province, China (Guo JS)
*Correspondence to: Jia-song Guo, Ph.D., jiasongguo@smu.edu.cn.
Figure 1 A sample of artif cial nerve conduit used to bridge the sciatic nerve defect.
Challenges for repairing peripheral nerve defects using 3D nerve grafts with nerve tissue-derived extracellular matrix and basement membrane tube-like conduits
The repair and reconstruction of nerve defects is a major challenge in the f eld of peripheral nerve injury. To address short-segment nerve defect repair, end-to-end anastomosis is adopted. For long-segment nerve defect repair, however, autograft remains the gold standard because it contains the structural and biological components ideally matched to the requirements of the peripheral nerve. However, the use of autografts has some disadvantages, including donor site morbidity and the limited availability of expendable autogenous nerve graft tissue. As an alternative, an allograft comprising a nerve guide/conduit, can be used to treat nerve defects up to a few centimeters long (Meek and Coert, 2008). We previously ref ned the extraction method for this (Sondell et al., 1998) and were able to obtain chemically extracted acellular nerve allografts from canine and human sciatic nerves, which have more interfascicular epineurium, and a thicker perineurium and epineurium than rat sciatic nerves. After verifying low immunogenicity of these constructs, we successfully repaired 5-cm sciatic nerve defects in a canine model (Zhong et al., 2007). Furthermore, 21 patients with nerve defects were successfully treated with chemically extracted acellular nerve allografts, with all patients regaining sensation and motion (Guo et al., 2008). As a result, we believe that the best alternative for peripheral nerve gap repair is a nerve conduit, which recreates in vivo structures and contains essential components needed for peripheral nerve regeneration (Zhou et al., 2014).
It is well-known that peripheral nerves have a def ned structure comprising Schwann cells from the myelin sheath wrapped around axons–similar to a wire wrapped by insulators–to facilitate the maintenance of normal nerve function. Furthermore, Schwann cells are wrapped by a layer of basement membrane comprising a specialized extracellular matrix (ECM) with unique components. Similar to other tissues, ECM in the peripheral nerve basement membrane is a natural substrate, which not only provides a support structure and attachment site for cells, but supplies biological signals that play important roles in and signif cantly impact cell migration, adhesion, dif erentiation, proliferation, and gene expression. Peripheral nerve tissue ECM contains an abundance of macromolecules, such as collagens (type-I, -III, and -IV), laminin, fibronectin, chondroitin sulfate, proteoglycans, and nerve neurotrophic factors.
Collagens are structural proteins that are required for normal ECM assembly and organization, thus acting as repair materials for damaged peripheral nerves. Collagen type-I accounts for 90% of the ECM derived from nerves, so is commonly used for repairing peripheral nerve defects alone or in combination with other materials, such as poly(ε-caprolactone), chitosan, hyaluronic acid and Matrigel™. Many ECM proteins produced by Schwann cells, such as f bronectin and laminin, also referred to as neurite-promoting factors, play an important role in adhesion of the axon with neural cell adhesion molecule, N-cadherin, for neural cell regeneration. As a prominent constituent in the peripheral nerve ECM, laminin exists predominantly in the endoneurium and perineurium of the peripheral nerve and is mainly produced by Schwann cells. Within the peripheral nervous system, laminin is composed of α, β, and γ chains. The f ve α chains, three β chains, and three γ chains, are not only widely distributed along the cell surface in a continuous region, but are also a ubiquitous component of the tight network of collagen type-IV, glycoproteins, and proteoglycans within the basement membrane. Laminin is usually combined with other materials, such as collagen or chitosan, to enhance axonal growth. Laminin could therefore be a suitable target for delivery of exogenous neurotrophic factors to repair nerve defects. Fibronectin interacts with collagen, heparin and integrin family cell-surface receptors, to inf uence cell adhesion, morphology, migration, thrombosis, and embryonic dif erentiation. Fibronectin is predominantly located in the perineurium of the peripheral nervous tissue but is only sparsely distributed along the cell surface. For peripheral nerve defect repair, f bronectin is usually added alone, or in combination with other components, such as alginate hydrogel matrix, collagen and laminin, to enhance nerve repair. As a result, these ECM components not only provide structural support to guide newborn neurof lament extensions, but play important roles as promoters or inhibitors. Therefore, ECM components should be considered as suitable biomimetic materials for conduit generation.
During peripheral nerve regeneration, nerve axons extending to distal target organs not only need chemical inducement, but also need physical structure support and guidance. Therefore, microstructure and three-dimensional (3D)-architecture play important roles in supporting defected nerve regeneration. In order to mimic the structures and components of native nerves, great ef orts have been made in the development of nerve conduits. Aside from the consideration of suitable materials, fabrication techniques are also important for this approach. Previous studies have designed open-porous, biodegradable and micro-structured nerve guides, with continuous, longitudinally orientated channels for orientated nerve growth. In both rat and rabbit static nerve injury models, these biomaterials have shown promising results (Bozkurt et al., 2009; Hu et al., 2009; Zhang et al., 2013). Yao et al. (2010) designed and fabricated robust multichannel collagen conduits for nerve regeneration, and found that the number of channels in the scaffold greatly af ects the physicochemical properties. Material surface topography has also been shown to af ect the biological behavior of cells in vitro and ef ect peripheral nerve regeneration in vivo. Wang et al. (2015) have shown that conduits developed using micro-tube array sheets have the potential to facilitate peripheral nerve regeneration by modifying critical biological behaviors and guiding orientated nerve growth. Electrospinning is a simple and versatile technique that has recently been used to fabricate f brous tissue-engineered scaf olds with a similar ECM to f brous structures, which have been used to enhance nerve regeneration. Using these methods, however, the most dif cult challenge is how to generate a conduit comprising many micro-tubes representative of the structure of the tube-like nerve endoneurium (Peng et al., 2012).
Three points must be considered when developing conduits for nerve regeneration. First, the structure of the conduits should contain many microtubes in parallel to the axis of the peripheral nerve. Second, the wall of the microtubes needs be made of f bers, which should also be parallel to the axis of the peripheral nerve. Third, the size of the microtubes and diameter of the f bers should direct the extension of neurites from both types of neurons and guide the oriented migration of Schwann cells, without af ecting other critical biological properties, such as proliferation and neurotrophin expression. Based on these considerations, the generation of suitable nerve conduits with a biomimetic ECM component and endoneurium-like microtube structure is certainly a challenge for future developments in the f eld of nerve repair and regeneration.
This work was funded by the National Natural Science Foundation of China, No. 31170946, 31300805; the People’s Liberation Army 12thFive-Year Plan Period, No. BWS11J025; and the National Basic Research Program of China, No. 2012CB518106, 2014CB542201.
Jiang Peng*, Yu Wang
Institute of Orthopedics, Chinese PLA General Hospital, Beijing, China (Peng J, Wang Y)
The Neural Regeneration Co-innovation Center of Jiangsu Province, Nantong, Jiangsu Province, China (Peng J)
*Correspondence to: Jiang Peng, M.D., Ph.D., pengjiang301@126.com.
Ahmed MR, Venkateshwarlu U, Jayakumar R (2004) Multilayered peptide incorporated collagen tubules for peripheral nerve repair. Biomaterials 25:2585-2594.
Aszmann OC, Roche AD, Salminger S, Paternostro-Sluga T, Herceg M, Sturma A, Hofer C, Farina D (2015) Bionic reconstruction to restore hand function after brachial plexus injury: a case series of three patients. Lancet 385:2183-2189.
Barrie KA, Steinmann SP, Shin AY, Spinner RJ, Bishop AT(2004) Gracilis free muscle transfer for restoration of function after complete brachial plexus avulsion. Neurosurg Focus 16:E8.
Behrend C, Reizner W, Marchessault JA, Hammert WC (2011) Update on advances in upper extremity prosthetics. J Hand Surg Am 36:1711-1717. Bhatia M (2010) Developmental biology. Microenvironment mimicry. Science 329: 1024-1025.
Bozkurt A, Deumens R, Beckmann C, Olde Damink L, Schügner F, Heschel I, Sellhaus B, Weis J, Jahnen-Dechent W, Brook GA, Pallua N (2009) In vitro cell alignment obtained with a Schwann cell enriched microstructured nerve guide with longitudinal guidance channels. Biomaterials 30:169-179.
Brooks DN, Weber RV, Chao JD, Rinker BD, Zoldos J, Robichaux MR, Ruggeri SB, Anderson KA, Bonatz EE, Wisotsky SM, Cho MS, Wilson C, Cooper EO, Ingari JV, Safa B, Parrett BM, Buncke GM (2012) Processed nerve allografts for peripheral nerve reconstruction: a multicenter study of utilization and outcomes in sensory, mixed, and motor nerve reconstructions. Microsurgery 32:1-14.
Chen Z, Zhang J, Chen T, Chen Z, Li H, Zhang EW, Lineaweaver WC, Zhang F (2011) Study of sensory and motor fascicles in brachial plexus and establishment of a digital three-dimensional graphic model.Ann Plast Surg 67:615-619.
Cho Y, Sloutsky R, Naegle KM, Cavalli V (2013) Injury-induced HDAC5 nuclear export is essential for axon regeneration. Cell 155:894-908.
Dai X, Zhang XD, Jiang YW, Tang YJ, Zhang LH, Liu ZY, Dong WR, Guo JS (2010) Isolation and identif cation of the peripheral blood derived mesenchymal stem cells and induction of its into Schwann cells. Zhongguo Linchuang Jiepouxue Zazhi 28:203-207.
Doi K, Hattori Y, Sakamoto S, Dodakundi C, Satbhai NG, Montales T (2013) Current procedure of double free muscle transfer for traumatic total brachial plexus palsy. JBJS Essent Surg Tech 3:e16.
Dy CJ, Garg R, Lee SK, Tow P, Mancuso CA, Wolfe SW (2015) A systematic review of outcomes reporting for brachial plexus reconstruction. J Hand Surg Am 40:308-313.
Fernández-de-las-Peñas C, Ambite-Quesada S, Ortega-Santiago R, Martínez-Perez A, Díaz HF, Martínez-Martín J, Parejam JA (2013) Catechol-O-methyltransferase Val158Met polymorphism is associated with pain and disability, but not widespread pressure pain sensitivity, in women with carpal Tunnel syndrome. Pain Physician 16:E591-600.
Forciniti L, Ybarra III J, Zaman MH, Schmidt CE (2014) Schwann cell response on polypyrrole substrates upon electrical stimulation. Acta Biomater 10:2423-2433.
Frostick SP, Yin Q, Kemp GJ (1998) Schwann cells, neurotrophic factors, and peripheral nerve regeneration. Microsurgery 18:397-405.
Ginanneschi F, Filippou G, Bonifazi M, Frediani B, Rossi A (2014) Ef ects of local corticosteroid injection on electrical properties of aβ-f bers in carpal tunnel syndrome. J Mol Neurosci 52:525-530.
Gu X, Ding F, Williams DF (2014) Neural tissue engineering options for peripheral nerve regeneration. Biomaterials 35:6143-6156.
Gu YD, Zhang GM, Chen DS (1989) Brachial plexus avulsion injury treatment with contralateral C7 transfer. Chin Med J (Chin) 69:563-565.
Guimard NK, Gomez N, Schmidt CE (2007) Conducting polymers in biomedical engineering. Prog Polym Sci 32:876-921.
Guo JH, Liu Y, Lv ZJ, Wei WJ, Guan X, Guan QL, Leng ZQ, Zhao JY, Miao H, Liu J (2015) Potential neurogenesis of human adipose-derived stem cells on electrospun catalpol-loaded composite nanof brous scaf olds. Ann Biomed Eng 43:2597-2608.
Guo YZ, Wang Y, Sun MX, Tao S, Tang PF, Liang YT, Lu SB (2008) Clinical study on chemically extracted nerve allograft. Zhonghua Shenjing Waike Zazhi 24:916-918.
Hari A, Djohar B, Skutella T, Montazeri S (2004) Neurotrophins and extracellular matrix molecules modulate sensory axon outgrowth. Int J Dev Neurosci 22:113-117.
Harrison RG (1911) On the stereotropism of embryonic cells. Science 34:279-281.
Hirata H, Hibasami H, Yoshida T, Ogawa M, Matsumoto M, Morita A, Uchida A (2001) Nerve growth factor signaling of p75 induces dif erentiation and ceramide-mediated apoptosis in Schwann cells cultured from degenerating nerves. Glia 36:245-258.
Höke A, Redett R, Hameed H, Jari R, Zhou C, Li ZB, Griffin JW, Brushart TM (2006) Schwann cells express motor and sensory phenotypes that regulate axon regeneration. J Neurosci 26:9646-9655.
Hu X, Huang J, Ye Z, Xia L, Li M, Lv B, Shen X, Luo Z (2009) A novel scaffold with longitudinally oriented microchannels promotes peripheral nerve regeneration. Tissue Eng Part A 15:3297-3308.
Huang JF, Xiang J, Yan QJ, Li SP, Song LT, Cai XH (2013) Dog tibial nerve regeneration across a 30-mm defect bridged by a PRGD/PDLLA/β-TCP/ NGF sustained-release conduit. J Reconstr Microsurg 29:77-88.
Huang L, Zhuang X, Hu J, Lang L, Zhang P, Wang Y, Chen X, Wei Y, Jing X (2008) Synthesis of biodegradable and electroactive multiblock polylactide and aniline pentamer copolymer for tissue engineering applications. Biomacromolecules 9:850-858.
Iwabuchi Y, Maki Y, Yoshizu T, Narisawa H (1999) Lack of topographical specif city in peripheral nerve regeneration in rats. Scand J Plast Reconstr Surg Hand Surg 33:181-185.
Jarocha D, Milczarek O, Wedrychowicz A, Kwiatkowski S, Majka M (2015) Continuous improvement after multiple mesenchymal stem cell transplantations in a patient with complete spinal cord injury. Cell Transplant 24: 661-672.
Jesuraj NJ, Santosa KB, Newton P, Liu Z, Hunter DA, Mackinnon SE, Sakiyama-Elbert SE, Johnson PJ (2011) A systematic evaluation of Schwann cell injection into acellular cold-preserved nerve grafts. J Neurosci Methods 197:209-215.
Jesuraj NJ, Nguyen PK, Wood MD, Moore AM, Borschel GH, Mackinnon SE, Sakiyama-Elbert SE (2012) Dif erential gene expression in motor and sensory Schwann cells in the rat femoral nerve. J Neurosci Res 90:96-104.
Jiang B, Zhang P, Jiang B (2010) Advances in small gap sleeve bridging peripheral nerve injury. Artif Cells Blood Substit Immobil Biotechnol 38:1-4.
Johnson EO, Zoubos AB, Soucacos PN (2005) Regeneration and repair of peripheral nerves. Injury 36:S24-29.
Keilhof G, Fansa H, Smalla KH, Schneider W, Wolf G (2000) Neuroma: a donor-age independent source of human Schwann cells for tissue engineered nerve grafts. Neuroreport 11:3805-3809.
Kennedy LH, Sutter CH, Leon Carrion S, Tran QT, Bodreddigari S, Kensicki E, mohney rp, sutter tr (2013) 2,3,7,8-tetrachlorodibenzo-p-dioxin-mediated production of reactive oxygen species is an essential step in the mechanism of action to accelerate human keratinocyte dif erentiation. Toxicol Sci 132:235-249.
Li CB, Chen ZG, Chen TY, Zhang F (2013) Research progress of peripheral nerve defect repair. Guoji Gukexue Zazhi 34:420-423.
Li F, Wang H, Li L, Huang C, Lin J, Zhu G, Chen Z, Wu N, Feng H (2012) Superoxide plays critical roles in electrotaxis of f brosarcoma cells via activation of ERK and reorganization of the cytoskeleton. Free Radic Biol Med 52:1888-1896.
Li M, Guo W, Zhang P, Li H, Gu X, Yao D (2013) Signal f ow and pathways in response to early Wallerian degeneration after rat sciatic nerve injury. Neurosci Lett 536:56-63.
Lian J, Lv S, Liu C, Liu Y, Wang S, Guo X, Nan F, Yu H, He X, Sun G, Ma X (2015) Ef ects of serial passage on the characteristics, cardiac and neural dif erentiation of human umbilical cord Wharton’s jelly-derived mesenchymal stem cells. Stem Cell Int.
Liu X, Yue Z, Higgins MJ, Wallace GG (2011) Conducting polymers with immobilized f brillar collagen for enhanced neural interfacing. Biomaterials 32:7309-7317.
Liu Y, Liu C, Nan F, Xiao J, Lian J, He X, Guo X, Sun G, Ma X (2013) Microcapsule co-culture system enhances neural dif erentiation of mesenchymal stem cells. J Hard Tissue Biol 22:241-248.
Lu L, Wang YN, Sun WH, Liu ZH, Zhang Q, Pu LJ, Yang K, Wang LJ, Zhu ZB, Meng H, Yang P, Du R, Chen QJ, Wang LS, Yu H, Shen WF (2013) Two-dimensional f uorescence in-gel electrophoresis of coronary restenosis tissues in minipigs: increased adipocyte Fatty Acid binding protein induces reactive oxygen species-mediated growth and migration in smooth muscle cells. Arterioscler Thromb Vasc Biol 33:572-580.
Lu Y, Belin S, He Z (2014) Signaling regulations of neuronal regenerative ability. Curr Opin Neurobiol 27:135-142.
Lundborg G, Dahlin LB, Danielsen N, Gelberman RH, Longo FM, Powell HC, Varon S (1982) Nerve regeneration in silicone chambers: inf uence of gap length and of distal stump components. Exp Neurol 76:361-375.
Ma J, Li W, Tian R, Lei W (2010) Ginsenoside Rg1 promotes peripheral nerve regeneration in rat model of nerve crush injury. Neurosci Lett 478:66-71.
Ma J, Liu J, Wang Q, Yu H, Chen Y, Xiang L (2013) The benef cial ef ect of ginsenoside Rg1 on Schwann cells subjected to hydrogen peroxide induced oxidative injury. Int J Biol Sci 9:624-636.
Ma J, Novikov LN, Kellerth JO, Wiberg M (2003) Early nerve repair after injury to the postganglionic plexus: an experimental study of sensory and motor neuronal survival in adult rats. Scand J Plast Reconstr Surg Hand Surg 37:1-9.
Ma TC, Willis DE (2015) What makes a RAG regeneration associated? Front Mol Neurosci 8:43.
Maness PF, Schachner M (2007) Neural recognition molecules of the immunoglobulin superfamily: signaling transducers of axon guidance and neuronal migration. Nat Neurosci 10:19-26.
May F, Weidner N, Matiasek K, Caspers C, Mrva T, Vroemen M, Henke J, Lehmer A, Schwaibold H, Erhardt W, Gänsbacher B, Hartung R (2004) Schwann cell seeded guidance tubes restore erectile function after ablation of cavernous nerves in rats. J Urol 172:374-377.
Meek MF, Coert JH (2008) US Food and Drug Administration/Conformit Europe-approved absorbable nerve conduits for clinical repair of peripheral and cranial nerves. Ann Plast Surg 60:466-472.
Menei P, Montero-Menei C, Whittemore SR, Bunge RP, Bunge MB (1998) Schwann cells genetically modif ed to secrete human BDNF promote enhanced axonal regrowth across transected adult rat spinal cord. Eur J Neurosci 10:607-621.
Meyer zu Hörste G, Prukop T, Nave KA, Sereda MW (2006) Myelin disorders: Causes and perspectives of Charcot-Marie-Tooth neuropathy. J Mol Neurosci 28:77-88.
Moore AM, Kasukurthi R, Magill CK, Farhadi HF, Borschel GH, Mackinnon SE (2009) Limitations of conduits in peripheral nerve repairs. Hand (N Y) 4:180-186.
Naidu M (2009) The role of cells, neurotrophins, extracellular matrix and cell surface molecules in peripheral nerve regeneration. Malays J Med Sci 16:10-14.
National Health and Family Planning Commission of China (2015) Quality Control of Stem Cell Preparations and Guidelines for Pre-Clinical Research (for Trial Implementation).
Pachuau J, Li DP, Chen SR, Lee HA, Pan HL (2014) Protein kinase CK2 contributes to diminished small conductance Ca2+-activated K+ channel activity of hypothalamic pre-sympathetic neurons in hypertension. J Neurochem 130:657-667.
Park KK, Liu K, Hu Y, Smith PD, Wang C, Cai B, Xu B, Connolly L, Kramvis I, Sahin M, He Z (2008) Promoting axon regeneration in the adult CNS by modulation of the PTEN/Mtor pathway. Science 322:963-966.
Pearse DD, Pereira FC, Marcillo AE, Bates ML, Berrocal YA, Filbin MT, Bunge MB (2004) cAMP and Schwann cells promote axonal growth and functional recovery after spinal cord injury. Nat Med 10:610-616.
Pei YZ, Liu X, Liu SS, Lv Q, Liu J, Kaplan DL, Zhu HS (2015) A mild process to design silk scaf olds with reduced b-sheet structure and various topographies at the nanometer scale. Acta Biomaterialia 13:168-176.
Peng J, Wang Y, Guo QY, Wang AY, Zhan Q, Wang X, Zhao Bin, Zhang L, Xu WJ, Lu SB, Zhao Z, Ren ZW (2012) Repairing Peripheral Nerve Gaps with Nerve Extracellular Matrix-Derived Scaffolds and Mesenchymal Stem Cells. Science (Regenerative Medicine in China):68-69.
Perier C, Tieu K, Guégan C, Caspersen C, Jackson-Lewis V, Carelli V, Martinuzzi A, Hirano M, Przedborski S, Vila M (2005) Complex I def ciency primes Bax-dependent neuronal apoptosis through mitochondrial oxidative damage. Proc Natl Acad Sci U S A 102:19126-19131.
Ramon Y, Cajal S (1959) Degeneration and regeneration of the nervous system. Hafner publishing Co: newyork.
Redett R, Jari R, Crawford T, Chen YG, Rohde C, Brushart TM (2005) Peripheral pathways regulate motoneuron collateral dynamics. J Neurosci 25:9406-9412.
Rivers TJ, Hudson TW, Schmidt CE (2002) Synthesis of a novel, biodegradable electrically conducting polymer for biomedical applications. Adv Funct Mater 12:33-37.
Sabongi RG, Fernandes M, Dos Santos JB (2015) Peripheral nerve regeneration with conduits: use of vein tubes. Neural Regen Res 10:529-533.
Schmidt CE, Shastri VR, Vacanti JP, Langer R (1997) Stimulation of neurite outgrowth using an electrically conducting polymer. Proc Natl Acad Sci U S A 94:8948-8953.
Sciancalepore M, Luin E, Parato G, Ren E, Giniatullin R, Fabbretti E, Lorenzon P (2012) Reactive oxygen species contribute to the promotion of the ATP-mediated proliferation of mouse skeletal myoblasts. Free Radic Biol Med 53:1392-1398.
Shear DA, Tate MC, Archer DR, Hof man SW, Hulce VD, Laplaca MC, Stein DG (2004) Neural progenitor cell transplants promote long-term functional recovery after traumatic brain injury. Brain Res 1026: 11-22.
Sondell M, Lundborg G, Kanje M (1998) Regeneration of the rat sciatic nerve into allografts made acellular through chemical extraction. Brain Res 795:44-54.
Sun XH, Che YQ, Tong XJ, Zhang LX, Feng Y, Xu AH, Tong L, Jia H, Zhang X (2009) Improving nerve regeneration of acellular nerve allografts seeded with SCs bridging the sciatic nerve defects of rat. Cell Mol Neurobiol 29:347-353.
Takahashi Y, Maki Y, Yoshizu T, Tajima T (1999) Both stump area and volume of distal sensory nerve segments inf uence the regeneration of sensory axons in rats. Scand J Plast Reconstr Surg Hand Surg 33:177-180.
Torigoe K, Tanaka HF, Takahashi A, Awaya A, Hashimoto K (1996) Basic behavior of migratory Schwann cells in peripheral nerve regeneration. Exp Neurol 137:301-308.
Wang GH, Xie RG, Tang JB, Mao T, Xing SG, Chen YL (2014) A case of carpal tunnel syndrome induced by lumbrical muscle belly. Zhonghua Shouwaike Zazhi 30:288.
Wang SF, Li PC, Xue YH, Yiu HW, Li YC, Wang HH (2013) Contralateral C7 nerve transfer with direct coaptation to restore lower trunk function after traumatic brachial plexus avulsion. J Bone Joint Surg Am 95:821-7, S1-2.
Wang W, Itoh S, Konno K, Kikkawa T, Ichinose S, Sakai K, Ohkuma T, Watabe K (2009) Ef ects of Schwann cell alignment along the oriented electrospun chitosan nanof bers on nerve regeneration. J Biomed Mater Res A 91:994-1005.
Wang XH, Pan MJ, Wen JK, Tang YJ, Hamilton AD, Li YY, Qian CH, Liu ZY, Wu WT, Guo JS (2014) A novel artificial nerve graft for repairing longdistance sciatic nerve defects: a self-assembling peptide nanof ber scaf old-containing poly(lactic-co-glycolic acid) conduit. Neural Regen Res 9:2132-2141.
Wang Y, Wang W, Wo Y, Gui T, Zhu H, Mo X, Chen CC, Li Q, Ding W (2015) Orientated guidance of peripheral nerve regeneration using conduits with a microtube array sheet (MTAS). ACS Appl Mater Interfaces 7:8437-8450.
Wang YL, Gu XM, Kong Y, Feng QL, Yang YM (2015) Electrospun and woven silk f broin/poly(lactic-coglycolic acid) nerve guidance conduits for repairing peripheral nerve injury. Neural Regen Res 10:1635-1642.
Weiss P, Taylor AC (1944) Further experimental evidence against “neurotropism” in nerve regeneration. J Exp Zool 95:233.
Witzel C, Rohde C, Brushart TM (2005) Pathway sampling by regenerating peripheral axons. J Comp Neurol 485:183-190.
Wu GF, Pan MJ, Wang XH, Wen JK, Cao ST, Li ZL, Li YY, Qian CH, Liu ZY, Wu WT, Zhu LX, Guo JS (2015) Osteogenesis of peripheral blood mesenchymal stem cells in self assembling peptide nanof ber for healing critical size calvarial bony defect. Sci Rep In Press.
Yang L, Qu M, Wang Y, Duan H, Chen P, Wang Y, Shi W, Danielson P, Zhou Q (2013) Trichostatin A inhibits transforming growth factor-β-induced reactive oxygen species accumulation and myof broblast dif erentiation via enhanced NF-E2-related factor 2-antioxidant response element signaling. Mol Pharmacol 83:671-680.
Yang MJ, Shi QL, Zhou L, Gu YD (2006) Intraoperative distal motor latency as a predictor of carpal tunnel release. Zhonghua Shouwaike Zazhi 22:92-94. Yao L, Billiar KL, Windebank AJ, Pandit A (2010) Multichanneled collagen conduits for peripheral nerve regeneration: design, fabrication, and characterization. Tissue Eng Part C Methods 16:1585-1596.
Zhan XD, Gao MY, Jiang YW, Zhang WW, Wong WM, Yuan Q, Su H, Kang X, Dai X, Zhang W, Guo JS, Wu WT (2013) Nanof ber nerve scaf olds facilitate functional regeneration of peripheral nerve injury. Nanomedicine 9:305-315.
Zhang P, Han N, Wang T, Xue F, Kou Y, Wang Y, Yin X, Lu L, Tian G, Gong X, Chen S, Dang Y, Peng J, Jiang B (2013) Biodegradable conduit small gap tubulization for peripheral nerve mutilation: a substitute for traditional epineurial neurorrhaphy. Int J Med Sci 10:171-175.
Zhang PX, Yin XF, Kou YH, Xue F, Han N, Jiang BG (2015) Neural regeneration after peripheral nerve injury repair is a system remodelling process of interaction between nerves and terminal ef ector. Neural Regen Res 10:52.
Zhang YG, Huang JH, Hu XY, Sheng QS, Zhao W, Luo ZJ (2011) Omentum-wrapped scaf old with longitudinally oriented micro-channels promotes axonal regeneration and motor functional recovery in rats. PLoS One 6:e29184.
Zhang YG, Sheng QS, Qi FY, Hu XY, Zhao W, Wang YQ, Lan LF, Huang JH, Luo ZJ (2013) Schwann cell-seeded scaf old with longitudinally oriented micro-channels for reconstruction of sciatic nerve in rats. J Mater Sci Mater Med 24:1767-1780.
Zhao Z, Wang Y, Peng J, Ren Z, Zhang L, Guo Q, Xu W, Lu S (2014) Improvement in nerve regeneration through a decellularized nerve graft by supplementation with bone marrow stromal cells in f brin. Cell Transplant 23:97-110.
Zhong H, Chen B, Lu S, Zhao M, Guo Y, Hou S (2007) Nerve regeneration and functional recovery after a sciatic nerve gap is repaired by an acellular nerve allograft made through chemical extraction in canines. J Reconstr Microsurg 23:479-487.
Zhou X, Han D, Xu R, Li S, Wu H, Qu C, Wang F, Wang X, Zhao Y (2014) A model of metabolic syndrome and related diseases with intestinal endotoxemia in rats fed a high fat and high sucrose diet. PLoS One 9:e115148.
10.4103/1673-5374.170301
s and corresponding authors:
猜你喜欢
杂志排行

中国神经再生研究(英文版)的其它文章
- Intracellular sorting pathways of the amyloid precursor protein provide novel neuroprotective strategies
- The role of the Rho/ROCK signaling pathway in inhibiting axonal regeneration in the central nervous system
- VEGF in the nervous system: an important target for research in neurodevelopmental and regenerative medicine
- Studying neurological disorders using induced pluripotent stem cells and optogenetics
- Ef cacy of glucagon-like peptide-1 mimetics for neural regeneration
- Compliant semiconductor scaf olds: building blocks for advanced neural interfaces