Who is who after spinal cord injury and repair? Can the brain stem descending motor pathways take control of skilled hand motor function?
2015-02-07GuillermoGarcía-Alías,V.ReggieEdgerton
Who is who after spinal cord injury and repair? Can the brain stem descending motor pathways take control of skilled hand motor function?
Over the last years, anatomical, electrophysiological and genetic studies have carefully dissected the pathways connecting the brain and the spinal cord. Lawrence and Kuypers (1968) described the organization of the descending motor pathways in the non-human primate spinal cord. Although there are some diff erences between species regarding the precise anatomical location of each spinal pathway and the selective connectivity onto spinal interneurons and motoneurons, the pattern of organization described is conserved among the mammalian spinal cord (Courtine et al., 2007). Based on their description, the major descending motor pathways are grouped depending on their anatomical origin and their terminal distribution pattern in the spinal grey matter. The motor cortex projects corticospinal axons to the spinal cord, which mostly run in the contralateral cord and innervate the mid and dorsal grey matter neurons. On the other hand, the spinal pathways originating in the brain stem are subdivided in the lateral and ventromedial systems. The ventromedial system fi bers originate in the reticular formation and the vestibular complex, and terminate in the ventral and medial parts of the ventral horn grey matter. In contrast, the lateral system fi bers originate in the red nucleus and preferentially terminate in the dorsal and lateral parts of the dorsal and medial grey matter (Figure 1). Together with the corticospinal fi bers, the lateral brain-stem system contacts interneurons related to motorneurons of distal muscles, whereas on the other hand, the ventromedial system pathways contact interneurons related to motoneurons of proximal muscles. Functionally, the corticospinal and lateral brain stem pathways are involved in the control of distal arm and hand muscles involved in skilled forelimb movements. The ventral brainstem pathways are mostly involved in the control of the proximal musculature of the trunk and limbs involved in posture and locomotion. Behavioral and electrophysiological studies mostly performed in cats and non-human primates corroborate these anatomical fi ndings (Lemon, 2012). Furthermore, rodents with injuries in discrete areas of the spinal cord selectively impair specific motor functions, evidencing the functional specifi city of each spinal pathway and the spinal networks that they innervate (Schucht et al., 2002; Anderson et al., 2007)
However, the functional and anatomical dichotomy between the spinal systems controlling skilled and less skilled motor movements cannot be that clearly distinct. First, both systems cannot work independently. Take for example the task of reaching and grasping an object; a motor action which requires from the synchronized and precise set of skilled body movements. While extending the arm and shaping the opening of the hand to the contour of the object, the body posture is also adjusted by correcting the activity pattern of the trunk musculature. In addition, although the object manipulation requires of precise movement of the digits, the whole reaching and grasping process needs the participation of both distal and proximal arm muscles (McCrea et al., 2002). In this line, elegant electrophysiological studies have shown the convergence of reticulospinal and corticospinal inputs onto cervical spinal interneurons controlling not only proximal but also distal arm muscles, evidencing the role of the brain stem pathways in the control of hand movement (Riddle et al., 2009).
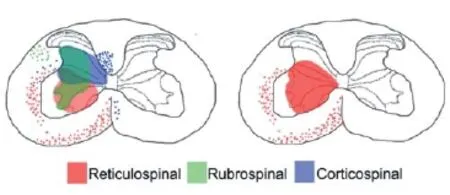
Figure 1 Diagrams depicting caudal cervical spinal cord transverse sections.
Despite the lack of a complete and detailed description of the circuitry involved in reaching and grasping, robust sets of experiments are starting to reveal the neuronal architecture connecting the cortex, the brainstem and the spinal cord (Alstermark and Isa, 2014). Strong structural diff erences appear between rodents and other species, which include: the absence of direct corticomotoneuron synapses and the unknown presence of a strong propiospinal system located at C4–5. Despite these diff erences, these studies evidenced the existence of an intricate connectivity between the motor cortex and motoneurons, including neuronal networks located at diff erent levels of the central nervous system (CNS), which are involved not only in the conduction but also in processing the reaching and grasping command. Importantly, in this motor engram, the reticulospinal tract together with the corticospinal and propisospinal pathways plays a preponderant role. A set of experiments by Alstermark et al. (2014) have revealed the importance of oligosynaptic excitatory corticofugal pathways to forelimb motorneurons in the control of skilled digit movements. This circuit may also be important in human walking recovery after stroke (Jang et al., 2014), although the functional weight in arm and hand function and recovery is unknown.
Recent experiments highlight the role played by brainstem motor descending pathways on the recovery of skilled hand function following injuries to the brain or spinal cord. These studies have shown the involvement of spinal tracts, other than the corticospinal and rubrospinal tracts, in the recovery of reaching and grasping in rodents subjected to spinal cord injuries. Several studies have reported that preserved descending motor axons projecting through the spared lateral and dorsolateral spinal cord must have been responsible for guiding the achieved recovery (Girgis et al., 2009; Hurd et al., 2013; Weishpurt et al., 2013). Anatomical studies are starting to fi nely corroborate these fi ndings. Rodents with unilateral stroke spontaneously recovered the ability to use their forelimbs to walk on along a rope. This recovery was accompanied by a strong reorganization of the cortico-reticulo and reticulo-spinal connectivity (Bachmann et al., 2014). Similarly, reticulospinal axons from rodents with a spinal cord lateral hemisection spontaneously branch above the injury and form contacts with propriospinal neurons, which bypass the injury and render motor recovery. These rearrangements were accompanied by substantial locomotor recovery (Filli et al., 2014). We recently performed a set of experiments in which we damaged both the corticospinal and rubrospinal tracts by infl icting a bilateral pyramidotomy at the level of the medulla and a bilateral crush of the dorsolateral funiculi at the C4spinal segment. A group of animals received intraspinal injections of chondroitinase ABC at C7(Garcia-Alias et al., 2015). It has been widely demonstrated that the digestion of chondroitin sulfate proteoglycans in the extracellular matrix promotes plasticity and recovery after injury to the CNS (Soleman et al., 2014). Our fi ndings showed that the animals which received chondroitinase and intense forelimb rehabilitation showed signifi cant redistribution of reticulospinal processes in the caudal cervical spinal cord compared to the injured and untreated animals (Figure 1). In addition, chondroitinase treated animals recovered their ability to reach and grasp.These results suggest that these therapies enhanced the sprouting of reticulospinal axons which favored the reconnectivity between the cortex and the motor cortex and led to the recovery of an animal’s manual dexterity. Taken together, these experiments indicate that some degree of hand functional recovery can occur after injury by promoting plasticity of pathways other than the corticospinal and rubrospinal pathways.
Due to the functional relevance of corticospinal tract integrity for the appropriate processing of sensorimotor input in humans, many experiments are focused on promoting spinal cord repair by increasing the regenerative or reparative capacities of corticospinal axons. Additional eff ort has been made to alternatively promote plasticity of the functionally complementary rubrospinal axons (Raineteau et al., 2001; Siegel et al., 2015). The recent identifi cation of brain stem pathways involved in the control (Esposito et al., 2014) and the recovery of arm and hand function, as described above, opens a new venue for the development of restorative strategies aimed at substituting the plasticity of the corticospinal axons with reticulospinal or other brain stem descending motor axons.
However, independently of the plasticity displayed by the other descending tracts, there still remains the question of whether or not, corticospinal reconnectivity must be an indispensable requirement for the effi cient repair of voluntary and skilled motor control. Although the human CNS is much more cortical than any other mammalian CNS (Swanson, 1995), and the important role played by the corticospinal tract in motor control, it must be noted that the specifi c sensory motor function displayed by the corticospinal tract is still unclear. Direct corticomotoneuron connection allows fine and fractionated digit control, and offers speed to movements. However, the absence of corticospinal axons “per se” does not abolish movement in non-human primates, suggesting that other spinal tracts must act as motor executors. If the candidate pathway to restore fi ne motor control has its origins in the brain stem, identifying its nature, connectivity and physiology will be imperative in order to fi nd strategies to overcome their limitations for spontaneous recovery and to enhance their structural and functional plasticity. An alternative approach, based in a much wider integrative interpretation of the CNS function, could off er a view in which the most important factor in promoting motor recovery is to maintain the continual fl ow of information along the neural axes independently of the nature of descending motor pathways which conduct the motor command. The descending volleys must cross the injury, which would act as an “electrical funnel” and once passed, they will need to interact with a newly shaped spinal networks. Furthermore, this descending volley will need to readapt to the restrictions created post-injury, which will require the integration and processing of limited information coming from above the injury with the altered information coming from the periphery.
Whether or not enhancing motor recovery can be driven by one or more specifi c pathways, activity dependent therapies will be a key element to teach the injured spinal cord to create, retain or strengthen any useful connections, and eliminate those which are maladaptive. This training could be further strength by the concomitant use of pharmacological and/or electrical interventions.
This work was supported by a grant from Wings for Life, Spinal Cord Foundation (WFL-US-004/11) to GGA, the National Institute of Biomedical Imaging and Bioengineering of the National Institutes of Health under Award Number U01EB015521 and the Christopher & Dana Reeve Foundation.
The authors thank Mr. Anthony Yeung for his reviewing the paper.
VRE holds a shareholder interest in NeuroRecovery Technologies. VRE is also President and Chair of the company’s Board of Directors. VRE, hold certain inventorship rights on intellectual property licensed by The Regents of the University of California to NeuroRecovery Technologies and it’s subsidiaries.
Guillermo García-Alías*, V. Reggie Edgerton
Department of Integrative Biology and Physiology, University of California, Los Angeles, CA, USA
*Correspondence to: Guillermo Garcia-Alias, Ph.D., guillermo284@gmail.com.
Accepted: 2015-07-18
orcid: 0000-0003-0197-0503 (Guillermo Garcia-Alias)
Alstermark B, Isa T (2012) Circuits for skilled reaching and grasping. Annu Rev Neurosci 35:559-578.
Alstermark B, Ogawa J, Isa T (2004) Lack of monosynaptic corticomotoneuronal EPSPs in rats: disynaptic EPSPs mediated via reticulospinal neurons and polysynaptic EPSPs via segmental interneurons. J Neurophysiol 91:1832-1839.
Anderson KD, Gunawan A, Steward O (2007) Spinal pathways involved in the control of forelimb motor function in rats. Exp Neurol 206:318-331.
Bachmann LC, Lindau NT, Felder P, Schwab ME (2014) Sprouting of brainstem-spinal tracts in response to unilateral motor cortex stroke in mice. J Neurosci 34:3378-3389.
Courtine G, Bunge MB, Fawcett JW, Grossman RG, Kaas JH, Lemon R, Maier I, Martin J, Nudo RJ, Ramon-Cueto A, Rouiller EM, Schnell L, Wannier T, Schwab ME, Edgerton VR (2007) Can experiments in nonhuman primates expedite the translation of treatments for spinal cord injury in humans? Nat Med 13:561-566.
Esposito MS, Capelli P, Arber S (2014) Brainstem nucleus MdV mediates skilled forelimb motor tasks. Nature 508:351-356.
Filli L, Engmann AK, Zörner B, Weinmann O, Moraitis T, Gullo M, Kasper H, Schneider R, Schwab ME (2014) Bridging the gap: a reticulo-propriospinal detour bypassing an incomplete spinal cord injury. J Neurosci 34:13399-13410.
García-Alías G, Truong K, Shah PK, Roy RR, Edgerton VR (2015) Plasticity of subcortical pathways promote recovery of skilled hand function in rats after corticospinal and rubrospinal tract injuries. Exp Neurol 266:112-119. Girgis J, Merrett D, Kirkland S, Metz GA, Verge V, Fouad K (2007) Reaching training in rats with spinal cord injury promotes plasticity and task specific recovery. Brain 130:2993-3003.
Hurd C, Weishaupt N, Fouad K (2013) Anatomical correlates of recovery in single pellet reaching in spinal cord injured rats. Exp Neurol 247:605-614. Jang SH, Chang CH, Lee J, Kim CS, Seo JP, Yeo SS (2013) Functional role of the corticoreticular pathway in chronic stroke patients. Stroke 44:1099-1104.
Lawrence DG, Kuypers HG (1968) The functional organization of the motor system in the monkey. I. The eff ects of bilateral pyramidal lesions. Brain 91:1-14.
Lawrence DG, Kuypers HG (1968) The functional organization of the motor system in the monkey. II. The eff ects of lesions of the descending brainstem pathways. Brain 91:15-36.
Lemon RN (2008) Descending pathways in motor control. Annu Rev Neurosci 31:195-218.
McCrea PH, Eng JJ, Hodgson AJ (2002) Biomechanics of reaching: clinical implications for individuals with acquired brain injury. Disabil Rehabil 24:534-541.
Raineteau O, Fouad K, Noth P, Thallmair M, Schwab ME (2001) Functional switch between motor tracts in the presence of the mAb IN-1 in the adult rat. Proc Natl Acad Sci U S A 98:6929-6934.
Riddle CN, Edgley SA, Baker SN (2009) Direct and indirect connections with upper limb motoneurons from the primate reticulospinal tract. J Neurosci 29:4993-4999.
Schucht P, Raineteau O, Schwab ME, Fouad K (2002) Anatomical correlates of locomotor recovery following dorsal and ventral lesions of the rat spinal cord. Exp Neurol 176:143-153.
Siegel CS, Fink KL, Strittmatter SM, Caff erty WB (2015) Plasticity of intact rubral projections mediates spontaneous recovery of function after corticospinal tract injury. J Neurosci 35:1443-1457.
Soleman S, Filippov MA, Dityatev A, Fawcett JW (2013) Targeting the neural extracellular matrix in neurological disorders. Neuroscience 253:194-213.
Swanson LW (1995) Mapping the human brain: past, present, and future. TINS 18:471-474.
Weishaupt N, Li S, Di Pardo A, Sipione S, Fouad K (2013) Synergistic eff ects of BDNF and rehabilitative training on recovery after cervical spinal cord injury. Behav Brain Res 239:31-42.
10.4103/1673-5374.165318 http://www.nrronline.org/
García-Alías G, Edgerton VR (2015) Who is who after spinal cord injury and repair? Can the brain stem descending motor pathways take control of skilled hand motor function? Neural Regen Res 10(11):1735-1736.
杂志排行

中国神经再生研究(英文版)的其它文章
- Intracellular sorting pathways of the amyloid precursor protein provide novel neuroprotective strategies
- The role of the Rho/ROCK signaling pathway in inhibiting axonal regeneration in the central nervous system
- VEGF in the nervous system: an important target for research in neurodevelopmental and regenerative medicine
- Studying neurological disorders using induced pluripotent stem cells and optogenetics
- Ef cacy of glucagon-like peptide-1 mimetics for neural regeneration
- Compliant semiconductor scaf olds: building blocks for advanced neural interfaces