大鼠基底外侧杏仁核组蛋白乙酰化对吗啡成瘾记忆的调控作用
2015-02-04乔晓孟殷芳圆李云肖魏曙光赖江华
乔晓孟,殷芳圆,李云肖,魏曙光,赖江华
大鼠基底外侧杏仁核组蛋白乙酰化对吗啡成瘾记忆的调控作用
乔晓孟,殷芳圆,李云肖,魏曙光,赖江华
西安交通大学法医学院 , 西安 710061
为了探索组蛋白乙酰化对吗啡成瘾记忆相关分子表达调控机制,文章选取健康成年雄性SD大鼠34只,随机分为正常对照组(n = 6)及基底外侧杏仁核(Basolateral amygdala, BLA)颅内定位手术组(n =28)。在条件性位置偏爱(Conditioned place preference, CPP)训练阶段,大鼠BLA内给予组蛋白去乙酰化酶抑制剂曲古抑菌素A(Trichostafin A, TSA)并且腹腔注射吗啡溶液(10.0 mg/kg),对照组给予相同体积的10%二甲基亚砜(Dimethyl sulfoxide,DMSO)或盐水。应用蛋白质印记方法,检测吗啡诱导大鼠CPP建立后BLA内组蛋白H3K14乙酰化和脑源性神经营养因子(Brain-derived neurotrophic factor, BDNF)蛋白表达水平。结果显示,腹腔注射10 mg/kg吗啡能成功建立CPP。吗啡、TSA联合给药组大鼠比单纯吗啡给药组大鼠表现出更强烈的CPP(<0.0001)。吗啡和TSA都能使BLA内的组蛋白H3乙酰化水平和BDNF的表达显著增高(< 0.0001),同时二者之间具有协同作用。结果表明,大鼠BLA内组蛋白乙酰化水平与吗啡成瘾记忆形成有关,抑制BLA内组蛋白去乙酰化酶(Histone deacetylases, HDACs)的活性可强化吗啡诱导的线索记忆的形成;大鼠BLA内BDNF参与了吗啡诱导的线索记忆的形成并可能受到组蛋白乙酰化的调控。
吗啡;条件性位置偏爱;基底外侧杏仁核;组蛋白乙酰化
药物成瘾是一种慢性、复发性脑疾病,其主要特征为对药物的耐受、强烈渴求、强迫性觅药行为,以及中断用药后出现戒断症状[1],不仅会给成瘾者自身造成严重的精神和躯体危害,还会引起严重的社会经济和公共卫生问题。《2013年中国禁毒报告》显示,截至2012年底,我国登记在册的吸毒人数有209.8万余,其中滥用阿片类毒品者有127.2万人,占60.6%。目前对以吗啡为代表的阿片类药物成瘾的发生、发展机制仍知之甚少,并且对阿片类药物成瘾的临床防治也缺乏理论指导。因此,对吗啡成瘾的神经生物学机制进行研究就显得十分紧要和迫切。
研究提示,表观遗传变异的长时稳定性可能是成瘾记忆长期存在的一个重要机制[2]。组蛋白乙酰化是表观修饰的重要方式之一,越来越多的证据表明,组蛋白的乙酰化-去乙酰化调控机制在许多重大脑功能中发挥着至关重要的作用,如神经元分化[3]、神经退行性变[4,5]、昼夜比率[6]、癫痈发作[7,8]、记忆形成及药物成瘾[9~11]。组蛋白去乙酰化酶(Histone deacetylases, HDACs)可通过催化组蛋白残基末端的去乙酰化反应来调节下游基因的表达,从而产生持久的神经生物学改变并最终影响个体行为调控。然而,HDACs在吗啡成瘾记忆形成中的作用及其对记忆相关重要基因的表达调控尚不清楚。脑源性神经营养因子(Brain-derived neurotrophic factor, BDNF)已经被证明对学习记忆、突触可塑性以及维持多巴胺能神经元的存活和功能起到十分重要的作用[12],然而在慢性吗啡作用下这些分子的具体变化尚未见报道。本研究通过观察大鼠吗啡成瘾条件性位置偏爱(Conditioned place preference, CPP)建立后大鼠BLA内组蛋白H3K14的乙酰化水平和BDNF的蛋白表达水平,探索组蛋白乙酰化对记忆相关分子的表达调控。
1 材料和方法
1.1 实验动物
健康成年雄性SD大鼠34只,8周龄左右,体重230~250 g,由西安交通大学动物实验中心提供。随机分为正常对照组(即homecage组,不参与任何手术及训练;= 6)、基底外侧杏仁核(Basolateral amygdala, BLA)颅内定位手术组(= 28)。BLA颅内定位手术根据造模后的不同给药方案分为4个亚组,分别为:盐水+二甲基亚砜(Dimethyl sulfoxide, DMSO)组(saline + DMSO,= 7),盐水+曲古抑菌素A(Trichostafin A,TSA)组(saline + TSA,= 7),吗啡+DMSO组(morphine + DMSO,= 7),吗啡+TSA组(morphine + TSA,= 7)。各组分笼饲养,自由饮水、进食。
1.2 BLA颅内定位术
大鼠用20%戊巴比妥钠,按50 mg/kg腹腔注射麻醉。俯卧位固定于大鼠脑立体定位仪(深圳瑞沃德生命科技公司)上,沿颅顶正中线切开皮肤,暴露头骨,按照G. Paxinos大鼠脑立体定位图谱第五版[13]定位:前囟−2.9 mm;中线两侧旁±5.0 mm;距颅骨表面高度−7.5 mm,用标记笔标记,用牙科钻在标记处钻孔,在颅骨表面固定2颗微型螺钉,去除进样器,先左后右,分别插入套管至BLA注射区,用玻璃离子体水门汀(粉:水=1 : 1)覆盖整个脑切面待干燥后抹一层牙托粉(义齿基托树脂Ⅱ型)进一步固定。每次BLA内微量给药时,用尖端突出于引导套管1.0 mm、容量为1.0 μL的微量注射器,抽吸1.0 μL的TSA(美国Sigma-Aldrich公司)溶液,将注射器经引导套管插入BLA上方,用同一速度缓慢推注药物注射入BLA,注射用时大于60 s。药物全部推进完毕后针头原位停留60 s以促进药物在脑组织中的扩散,并防止外溢。对照组大鼠双侧给予10%DMSO(美国Sigma-Aldrich公司)作为对照。
1.3 吗啡CPP模型的建立与测试
自然偏爱测试(1~2 d):建立CPP前,第1 d上午8: 00将实验箱(上海吉量软件科技公司)隔板取走,让大鼠自由在实验箱两侧适应观察15 min,第2 d让大鼠在箱子内自由走动15 min,并记录其在两侧各停留的时间,确定大鼠偏好倾向,以非天然偏爱箱(白箱)为吗啡伴药箱。吗啡CPP建立:第3 d,吗啡组大鼠BLA核团内微量注射165mmol/L的TSA溶液(溶于DMSO中)或等体积的10%DMSO,30 min后腹腔注射10 mg/kg吗啡(沈阳第一制药厂)溶液,然后将其放置于白箱,用隔板封闭CPP箱,使大鼠在白箱中训练30 min;同时,盐水组BLA内注射TSA或DMSO,随后腹腔注射生理盐水,然后将其放置于白箱,用隔板封闭CPP箱,使大鼠在白箱中训练30 min。第4 d,所有大鼠BLA内给药后均腹腔注射生理盐水,然后将其放置于黑箱,用隔板封闭CPP箱,使大鼠在黑箱中训练30 min。条件性测试:第5 d,所有大鼠均不给予任何药物,将CPP箱中间的隔板打开,使大鼠可在两箱中自由穿梭,测试时间为15 min (900 s),记录大鼠在各箱中的停留时间、活动路程及穿梭次数。
为更好地观察TSA对吗啡CPP建立过程的影响,本研究共经历4个训练循环(3~4 d、6~7 d、9~10 d、12~13 d)和4次条件性测试(5 d、8 d、11 d、14 d),如图1所示。
1.4 蛋白检测
CPP测试结束后即刻以颈椎脱臼法处死大鼠,剥离大脑,分离BLA组织,每20 mg加入1.0 mL预冷的RIPA裂解液(预先加入蛋白酶抑制剂和磷酸酶抑制剂),提取蛋白质。加入样品缓冲液,沸水浴5 min,离心后取上清,进行12%SDS-PAGE电泳,采用半干法将蛋白转移至PVDF膜。用5%的牛血清蛋白封闭液室温封闭PVDF膜2 h。将抗乙酰化组蛋白H3K14兔多克隆抗体(美国Cell Signaling Technology公司)、抗BDNF兔多克隆抗体(美国Abcam Technology公司)、抗GAPDH小鼠多克隆抗体(美国Cell Signaling Technology公司)用封闭液稀释至1:1000浓度,4℃冰箱孵育过夜。TBS-T洗膜3次,每次10 min,将相应的二抗用TBS-T稀释至1:10000,室温下孵育1.5 h。TBS-T洗膜4次,每次10 min,进行化学发光检测。采用美国Bio-Rad公司的Quantity One软件分析条带结果,计算条带的光密度,进行相对定量分析。计算检测的目的蛋白与内参蛋白(GAPDH)条带的光密度比值来表示检测蛋白的相对表达水平。

图1 吗啡CPP流程图
M: 吗啡; S: 盐水。
1.5 数据处理与统计分析
采用GraphPad Prism 6.0软件对数据进行统计分析,实验结果均使用均值±标准误(mean±SEM)来表示。大鼠总活动距离、总穿梭次数以及CPP评分均采用重复测量的双因素方差分析并采用Tukey’s post hoc 检验进行两两比较。BLA内BDNF表达量以相对于各组中Sal+DMSO组的相对倍数表示,采用双因素方差分析研究吗啡和TSA对蛋白表达的影响。以< 0.05为显著性检验水准。
2 结果与分析
2.1 大鼠体重变化
2.1.1 手术恢复期
手术结束后,大鼠经历连续5 d的恢复期,以消除因手术应激、伤痛、感染等因素影响总体代谢,从而对行为学实验造成干扰。在恢复期,本文测量了大鼠的体重变化,如图2所示。双因素方差分析表明,在整个恢复期手术组(experimental,n = 28)大鼠体重与Hc组(homecage,n = 6)相比没有明显区别(Ftime (4, 150)= 0.9523,= 0.4357;Fexperimental (1, 150)= 0.03969,= 0.8424)。在恢复期结束时,手术组大鼠平均体重为252.2±18.3 g,Hc组大鼠平均体重为256.5±16.7 g,说明手术组大鼠经历5 d的恢复期后,其基础代谢水平与对照组无显著差异。这一结果提示,手术或可能造成的疼痛、应激等因素并未对大鼠进食和能量代谢造成严重影响。
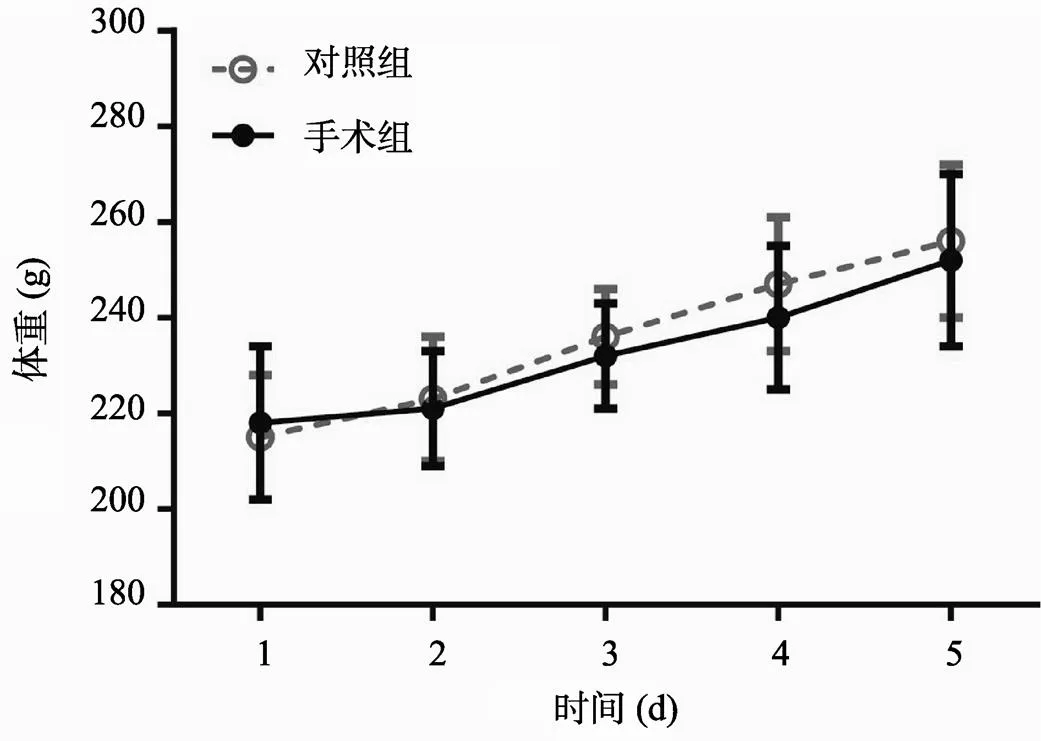
图2 大鼠恢复期体重变化
2.1.2 CPP实验期
本文分别测量了前测阶段(1 d、2 d)和4次CPP测试(5、8、11、14 d)时的大鼠体重,见图3。双因素方差分析表明,在为期14 d的CPP训练和测试阶段,所有大鼠的体重皆有显著增长(Ftime(5,162)=16.18,<0.0001),各个药物处理组大鼠的体重变化与Hc组相比没有明显区别(Ftreatment (4, 162)= 1.615,= 0.3066)。在CPP实验结束时,Hc组大鼠平均体重为213.3± 12.2 g,Sal(Saline)+DMSO组为215.2±15.0 g,Sal+ TSA组为329.4±13.1 g,Mor(Morphine)+DMSO组为293.6±12.6 g,Mor+TSA组为304.2±16.0 g。这说明大鼠BLA内给予TSA(165mmol/L)并且腹腔注射10.0 mg/kg吗啡溶液不会显著影响大鼠基础代谢,也可以排除总体代谢差异对脑内分子变化的干扰。
2.2 CPP行为测试
CPP行为测试结果如图4所示。Pre-test的CPP评分表明4个实验组大鼠之间基础偏爱值没有差异,所有大鼠皆自然偏爱黑箱且在黑箱中的停留时间基本相同,CPP评分在−46.0~−149.6 s之间。Sal+ TSA组大鼠在整个实验过程中均未产生显著的CPP,表明BLA内给165mmol/L TSA本身不具有奖赏效应,不能诱导位置偏爱的形成。Mor+DMSO组大鼠在Test 3(< 0.01)和Test 4(< 0.0001)中表现出显著的伴药箱(白箱)偏爱,表明腹腔注射10 mg/kg吗啡可成功诱导大鼠建立CPP。Mor+TSA组大鼠在接受第一个循环的条件性训练后(Test 1,<0.05)即表现出显著的位置偏爱,而这种强烈的位置偏爱一直持续至实验结束(Test 4,<0.0001)依然存在。
2.3 BLA内组蛋白乙酰化及BDNF的表达变化
2.3.1 组蛋白H3乙酰化改变
双因素方差分析表明,吗啡和TSA对BLA内组蛋白H3K14乙酰化水平具有显著影响(Fmorphine(1,28)= 68.1,< 0.0001;FTSA (1, 28)= 134,<0.0001),如图5所示。Mor+TSA联合给药后,BLA内的组蛋白H3乙酰化水平比Sal+DMSO组增高4.82倍,与Mor+ DMSO组相比也明显增高(<0.05)。以上结果提示,腹腔注射吗啡或BLA内给予TSA(165mmol/L)皆能显著提高BLA内组蛋白H3(K14)乙酰化水平,且二者具有协同作用。
2.3.2 BDNF表达改变
油茶病虫害较少,主要是油茶炭疽病、油茶软腐病、油茶煤污病和油茶尺蠖(又叫量步虫)。均造成一定程度的落果、落叶和枝枯,造成减产,但不会死树毁林。这几种病都与油茶林经营管理水平密切相关。林内通风透光,树体健壮,肥、水、土条件好,病枝、病叶、病果、过密枝得到及时剪除和深埋,发病就轻,反之就较重。品种的抗病能力也有差异,造林前可以选择抗性强的品种。发病期,可自配波尔多液控制病情。实在有必要可喷施退菌特800倍液防治。量步虫幼虫身体光滑无毛,行动迟缓,可以直接人工捕杀。
对BLA内BDNF蛋白进行检测发现,吗啡和TSA皆对BDNF表达具有显著作用(Fmorphine(1,28)= 76.44,<0.0001;FTSA (1, 28)= 28.82,<0.0001),同时二者之间具有显著的交互作用(Fmorphine×TSA (1, 28)= 5.71,<0.05),如图6所示。Mor+TSA联合给药后,BLA内BDNF表达比Sal+DMSO组增高3.03倍(<0.0001)。
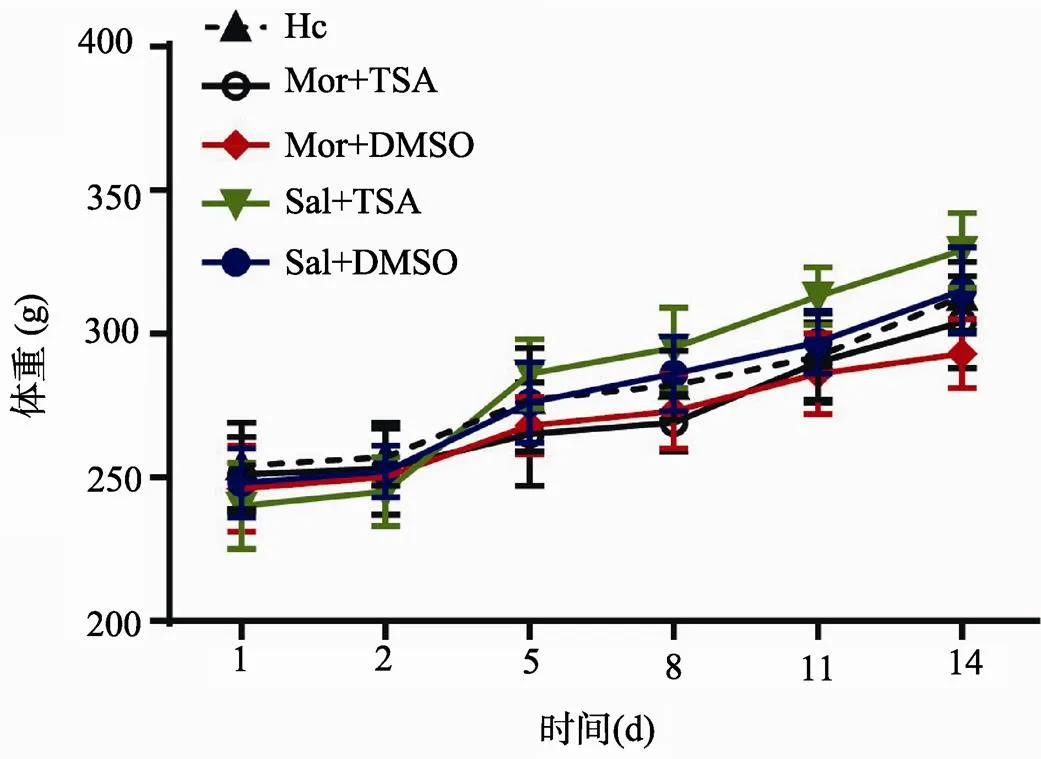
图3 大鼠CPP实验期间体重变化

图4 大鼠CPP评分
与各次测试中Sal + DMSO组比较,*< 0.05,**< 0.01,***< 0.0001;线段所指示的两组相互比较,#< 0.05,###< 0.0001。

图5 乙酰化组蛋白H3(aceH3K14)蛋白表达
与Sal + DMSO组比较,**< 0.01,***< 0.0001;线段所指示的两组相互比较,#< 0.05。
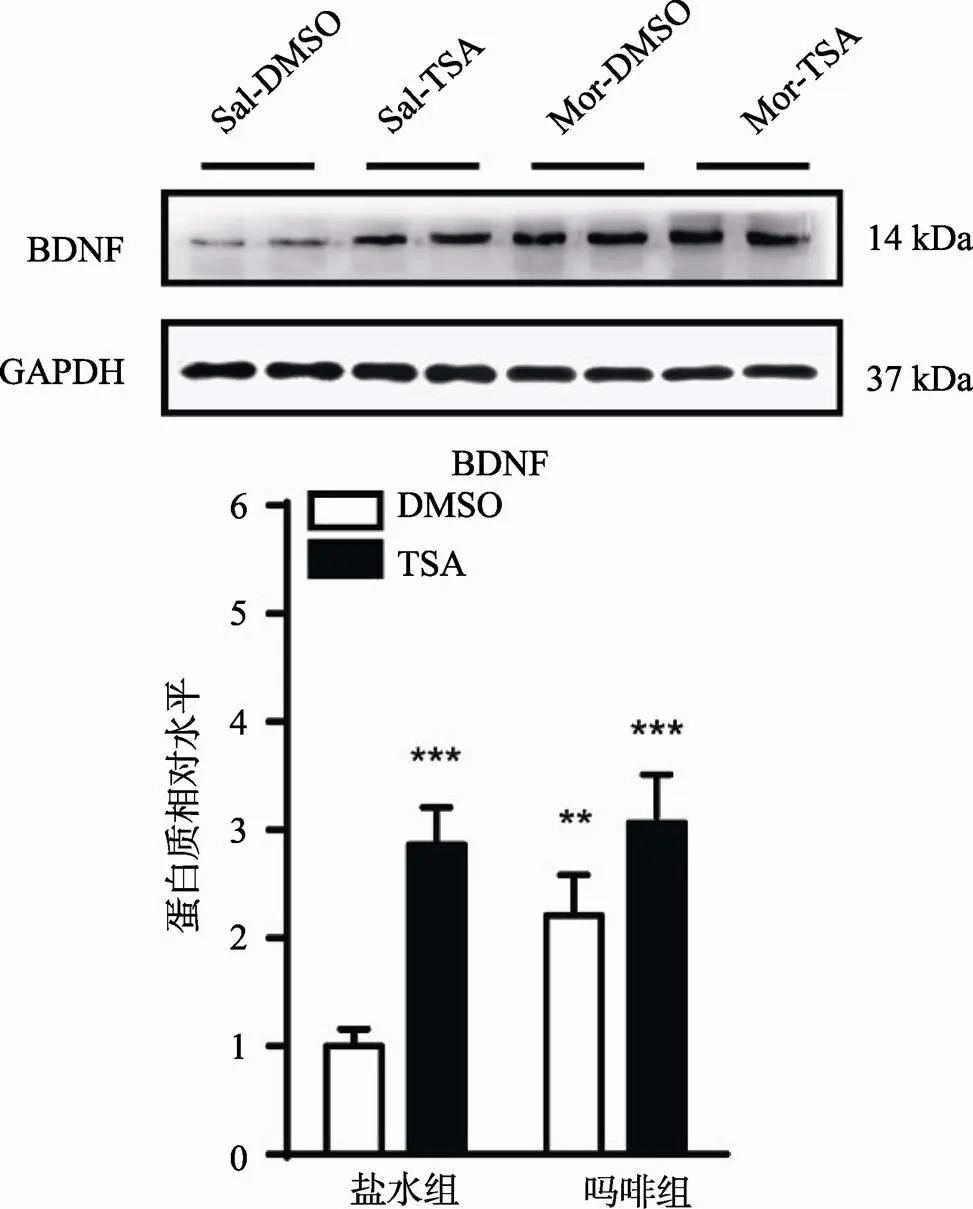
图6 BDNF蛋白表达改变
与Sal + DMSO组比较,**< 0.01,***< 0.0001。
3 讨 论
随着表观遗传学在后基因时代迅速发展,近年来,对表观遗传学的大量研究发现,基因表观事件的发生与精神活性物质的滥用和成瘾有着密切的关系[14]。介导表观遗传现象的分子机制主要集中在DNA 甲基化和组蛋白乙酰化修饰两种方式,其中组蛋白乙酰化在近几年提出的表观遗传学理论中占有重要地位,在精神活性物质所致的成瘾记忆和行为敏化中也发挥着重要作用。
本研究对BLA内组蛋白H3K14位点的总体乙酰化水平进行了检测,首次证明连续的吗啡全身给药可使BLA内H3K14位点的乙酰化水平增高,TSA也可显著增高H3K14位点的乙酰化水平,且二者对组蛋白H3K14乙酰化具有协同作用。然而在BLA内TSA和吗啡对组蛋白H3K14的协同作用是否是导致TSA能强化吗啡相关线索记忆的直接原因?H3K14乙酰化与药物相关记忆的一致性变化仅仅是一种巧合还是有必然的因果联系呢?尽管从我们目前的实验结果上尚不能直接得出确切结论,然而一种可能的解释是:BLA这一脑区负责了奖赏相关(正性奖励和无/负性奖励)线索记忆的形成,而组蛋白乙酰化可通过某种机制促进并加强了新记忆的形成过程。未来尚需运用更具特异性的干预手段对这一问题进行更深入的研究。
记忆的形成和存储涉及神经元的许多适应性改变,包括神经突触的功能和结构变化,而这些变化都需要相关基因的表达发生相应变化[15]。BDNF已经被证明对学习记忆、突触可塑性以及维持多巴胺能神经元的存活和功能起到十分重要的作用[12]。在本实验中,吗啡所诱导的组蛋白乙酰化增加能显著提高BLA内BDNF蛋白表达,提示BLA内BDNF可能参与了吗啡诱导的线索记忆的形成。这一结果显示,成瘾性药物可以通加强组蛋白乙酰化来增加转录因子活性,从而提高转录效率并促进突触可塑性改变。
本研究成功建立了吗啡诱导的CPP模型,并在BLA内给予TSA对组蛋白乙酰化水平进行干预,实验结果证实在TSA本身不影响CPP的状态下,能加强吗啡诱导的CPP形成。此外,分子结果提示吗啡所引起的组蛋白乙酰化增加对下游基因BDNF的表达可能有一定的调控作用,同时说明BDNF在线索相关奖赏记忆的形成中具有重要作用。根据我们目前的研究结果,要阐明吗啡诱导记忆改变所涉及的表观遗传学机制,将来尚需从以下几个方面进行深入研究:
(2)本研究仅观察了BLA内总体水平的组蛋白H3K14乙酰化改变,由于BLA内BDNF和DFosB与HDACs存在复杂的交互作用,未来应采用染色质免疫沉淀(Chromatin Immunoprecipitation,ChIP)技术观察BDNF和DFosB基因启动子区H3乙酰化以阐明这一问题。
(3)由于大脑复杂的神经信号传递通路和基因表达的调节,虽然组蛋白乙酰化已经开始用于研究药物滥用引起的行为反应,但是还不能用来准确的解释药物依赖的发病机理和成瘾行为的形成,在将来的研究中,从组织、细胞等多个水平结合药物、基因等多种干预方法进行成瘾行为、线索记忆及其分子机制的研究,将是进一步明确药物成瘾发生发展机制的重要内容。
[1] Cami J, Farré M. Mechanisms of disease: drug addiction., 2003, 349(10): 975–986.
[2] Hyman SE, Malenka R C, Nestler EJ. Neural mechanisms of addiction: the role of reward-related learning and memory., 2006, 29: 565–598.
[3] Hu XL, Wang YP, Shen Q. Epigenetic control on cell fate choice in neural stem cells., 2012, 3(4): 278–290.
[4] Hoshino M, Tagawa K, Okuda T, Murata M, Oyanagi K, Arai N, Mizutani T, Kanazawa I, Wanker EE, Okazawa H. Histone deacetylase activity is retained in primary neurons expressing mutant huntingtin protein.2003, 87(1): 257–267.
[5] Steffan JS, Bodai L, Pallos J, Poelman M, McCampbell A, Apostol BL, Kazantsev A, Schmidt E, Zhu YZ, Greenwald M, Kurokawa R, Housman DE, Jackson GR, Marsh JL, Thompson LM. Histone deacetylase inhibitors arrest polyglutamine-dependent neurodegeneration in., 2001, 413(6857): 739–743.
[6] Etchegaray JP, Lee C, Wade PA, Reppert SM. Rhythmic histone acetylation underlies transcription in the mammalian circadian clock., 2003, 421(6919): 177–182.
[7] Huang Y, Doherty JJ, Dingledine R. Altered histone acetylation at glutamate receptor 2 and brain-derived neurotrophic factor genes is an early event triggered by status epilepticus., 2002, 22(19): 8422–8428.
[8] Tsankova NM, Kumar A, Nestler EJ. Histone modifications at gene promoter regions in rat hippocampus after acute and chronic electroconvulsive seizures., 2004, 24(24): 5603–5610.
[9] Guan JS, Haggarty SJ, Giacometti E, Dannenberg JH, Joseph N, Gao J, Nieland TJF, Zhou Y, Wang XY, Mazitschek R, Bradner JE, DePinho RA, Jaenisch R, Tsai LH. HDAC2 negatively regulates memory formation and synaptic plasticity., 2009, 459(7243): 55–60.
[10] Levenson JM, O'Riordan KJ, Brown KD, Trinh MA, Molfese DL, Sweatt JD. Regulation of histone acetylation during memory formation in the hippocampus., 2004, 279(39): 40545–40559.
[11] Kumar A, Choi KH, Renthal W, Tsankova NM, Theobald DEH, Truong HT, Russo SJ, Laplant Q, Sasaki TS, Whistler KN, Neve RL, Self DW, Nestler EJ. Chromatin remodeling is a key mechanism underlying cocaine-induced plasticity in striatum., 2005, 48(2): 303–314.
[12] Andressoo JO, Saarma M. Signalling mechanisms underlying development and maintenance of dopamine neurons., 2008, 18(3): 297–306.
[13] Paxinos G, Watson C. The Rat Brain, in Stereotaxic Coordinates, 2nd ed. Sydney: Elsevier Academic Press, 1986.
[14] Renthal W, Nestler EJ. Epigenetic mechanisms in drug addiction., 2008, 14(8): 341–350
[15] Guan Z, Giustetto M, Lomvardas S, Kim JH, Miniaci MC, Schwartz JH, Thanos D, Kandel ER. Integration of long-term-memory-related synaptic plasticity involves bidirectional regulation of gene expression and chromatin structure., 2002, 111(4): 483–493.
(责任编委: 方向东)
The role of histone acetylation in the basolateral amygdala in morphine-associated memory in rats
Xiaomeng Qiao, Fangyuan Yin, Yunxiao Li, Shuguang Wei, Jianghua Lai
To examine the regulatory effect of histone acetylation on memory related molecules, 34 healthy male SD rats were randomly divided into control and basolateral amygdala (BLA) intracranial positioning operation groups. In the process of conditioned place preference (CPP) training, Trichostafin A (TSA) was administrated by the route of BLA and morphine was injected into enterocoelia with dimethyl sulfoxide or saline as control. Expression levels of H3K14 acetylation and brain-derived neurotrophic factor (BDNF) in BLA were evaluated by Western blotting.The results showed that CPP could be established by intraperitoneal injection of morphine. Compared with control groups, a stronger place preference was established and expression of H3K14 acetylation and BDNF was significantly increased in the group treated with TSA and morphine. In addition, there was a synergistic effect between morphine and TSA. Our results suggested that the level of histone acetylation in BLA is associated with the formation of morphine memory in rats. Inhibition of the activity of histone deacetylases in BLA can promote the formation of cue-associated memory induced by morphine and the involvement of BDNF in BLA maybe was regulated by histone acetylation.
morphine; conditioned place preference; BLA; histone acetylation
2014-11-20;
2015-03-01
国家自然科学基金项目(编号:81172910)资助
乔晓孟,博士研究生,专业方向:法医物证学。E-mail: xiaomeng416520@126.com
赖江华,教授,研究方向:药物成瘾分子机制研究。E-mail: Laijh1011@mail.xjtu.edu.cn
10.16288/j.yczz.14-406
2015-3-11 9:19:22
http://www.cnki.net/kcms/detail/11.1913.R.20150311.0919.003.html
