北极新奥尔松地区土壤中生物硅含量及形态特征
2015-01-27刘森冉祥滨臧家业王能飞
刘森 冉祥滨 臧家业 王能飞
(国家海洋局第一海洋研究所,海洋生态研究中心,山东青岛266061)
0 引言
硅是地壳中含量第二丰富的元素,平均含量为28.8 wt%[1],且其循环与碳循环密切相关,是地表地球化学过程中发挥关键作用的元素之一。在陆地生态系统中,植被通过多种方式对硅酸盐矿物风化产生影响[2-4],并吸收大量硅,形成具有一定形态的植硅体[5]。在陆地上,植物每年固定的硅达60-200 Tmol Si[6],与海洋生物硅(BSi) 的产量(240 Tmol Si·a-1)处于同一量级[7],两者构成了地表系统生物硅库的绝大部分。相比于成岩硅而言,以植硅体为主的生物硅比大多数土壤中硅酸盐矿物更易溶解,且数量庞大,因此已成为河流等地表水中溶解硅的主要来源[8-10],并在陆源硅向海洋的输送中发挥着关键作用[11]。由此可见,植硅体是全球硅循环中十分重要的环节,值得深入研究。
近些年来,人们针对地表硅循环的研究颇多,主要包括陆地、陆-海硅循环及人类活动对硅循环的影响等几方面,相关研究已逐渐成为全球环境变化研究的重要领域[12]。目前有关硅的研究区域涉及到湿地[5,13]、雨林[2]、湖泊[14]、水库[15]和河口[16-19]等多种陆地生态系统,但对极地地区硅的研究却极为少见。北极海区是全球碳循环的重要汇区[20],属于全球环境变化的重要研究领域。从现有研究来看,极区硅相关的研究主要集中在白令海、楚科奇海等海域[20-22]和极地湖泊[23],且研究内容以硅藻参与硅循环,重建古地理及古海洋环境演变过程等为主,而对极地陆区土壤中生物硅的研究尚属空白。更为重要的一点,北极斯瓦尔巴特群岛具有独特的气候特征,属于一类典型的生态系统,对其研究有利于揭示高纬度地区硅循环的基本规律。本文利用中国第五次北极考察获得的土壤、苔原植物样品,对北极土壤中生物硅的含量及土壤、植物中植硅体的形态进行初步探究,并与其他地区生物硅的相关研究进行对比,旨在为区域硅的生物地球化学循环提供科学依据。
1 材料与方法
1.1 区域概况
研究地区(11°43'56″-12°08'57″E, 78°53'46″-78°56'37″N)位于北冰洋上的斯瓦尔巴特群岛新奥尔松地区,距北极点很近,在巴伦支海和格陵兰海之间,由9个主岛和众多小岛组成。群岛多山地,沿岸多峡湾,近60%的区域被冰川覆盖,永冻土层厚达500 m[24],夏季地表以下2-3 m的土层解冻。新奥尔松地区为极地气候,冬夏各有100多天的极夜和极昼,但由于北大西洋暖流影响,群岛西部地区气候相对温和,年平均气温在-22-7℃之间,年均降水量约200 mm,又具有极地-海洋性气候特征。斯瓦尔巴特群岛植被主要是地衣和苔藓类,另有少数地柳、矮桦木等植物,65%的地区作为自然公园保护,以维护其独特的动植物资源,因此受到人为活动干扰较少。
1.2 样品采集与保存
2012年科研人员执行北极黄河站夏季科学考察任务期间,在斯瓦尔巴特群岛西北部新奥尔松地区采集了数种土壤样品及地表植被,土壤性质见表1,具体采样站位见图1。
共在此地区采集15个表层土壤样品,另采集发草(发草属,DeschampsiaBeauv.)、苔藓(黑藓属,Andreaea)等苔原植物;将采集到的土壤样品和植物样品分袋装好,冷冻,带回实验室分析。

表1 北极新奥尔松地区土壤样品采样点坐标、地貌和土壤性质Table 1.Topography and soil properties of sampling stations in the Ny-Ålesund, Arctic
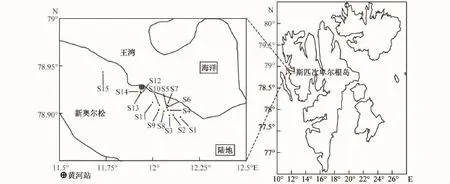
图1 北极新奥尔松地区采样区域站位示意图Fig.1.Location of sampling stations in the Ny-Ålesund, Arctic
1.3 实验方法
1.3.1 可溶出态硅的提取与测定
称取1.00 g烘干土样于洁净、干燥的离心管中,加10 mLMilli-Q水,室温下连续震荡4 h,离心,取上清液,适当比例稀释后测定可溶出态硅的含量,以 μg Si·g-1计[25-26]。
1.3.2 生物硅的提取与测定
称取100 mg烘干、研磨、过筛后的土样于洁净、干燥的离心管中,加10 mL 0.5%HCl,反应30 min后加 Milli-Q水,离心、清洗残留 HCl。烘干,加40 mL 2%Na2CO3溶液,在85℃恒温水浴下连续提取8 h,期间每隔1 h离心、取上清液125μL,适当比例稀释后测定硅浓度,并与时间作图,曲线线性部分与纵轴的截距即为样品生物硅的含量,以mg Si·g-1计[27]。提取出的可溶出态硅和生物硅用QuAAtro营养盐自动分析仪(硅钼蓝法)进行测定,溶液浓度在1-10μmol/L时准确度为5%-10%,浓度在10-100μmol/L时准确度为1%-5%。
1.3.3 颗粒有机碳、颗粒有机氮的测定
称取适量烘干土样于洁净、干燥的离心管中,加6 N HCl去除无机碳成分,反应完全后加Milli-Q水离心、清洗3-5遍。烘干,取一定量土样称重,用元素分析仪(EURO EA3000)测定颗粒有机碳、氮的含量[28-29],土壤有机碳和有机氮样品分析的相对标准偏差分别为3.30%和1.20%。
1.3.4 生物硅的形态提取
土壤:称取5 g烘干土样放入烧杯中,加30%H2O2去除有机质,再加入1∶1 HCl去除碳酸盐,重复此过程至样品反应完全;加 Milli-Q水,离心(2 000 r·min-1,2 min),倾去上清液,去除粘土,重复此过程至上层溶液澄清;用ZnBr2重液(密度为2.35 g·cm-3)浮选;浮选后的植硅体样品离心清洗后制片[30],用Nikon E100显微镜观察,每个样品统计200个以上的生物硅的形态。
植物:分别将发草茎叶、发草根部、苔原植物清洗,烘干,剪成小段,称重后放入烧杯,加入少量浓HNO3,加热,至有机质绝大部分被氧化分解,样品不再呈粘稠状,溶液澄清。样品烧干,稍冷却后加数毫升HClO4,加热,除去少量未能被HNO3氧化的有机物,最后,烧杯中的样品呈白色粉末状。加少量HCl,加热,使其中的 Ca、K等灰分分解,用 Milli-Q水将植硅体从烧杯中洗出,离心、清洗3-4次,制片[30],用 Nikon E100 显微镜观察,每个样品统计200个以上的植硅体的形态。
2 结果与讨论
2.1 可溶出态硅
土壤样品中可溶出态硅的含量如图2所示。调查区域内,土样的可溶出态硅的平均值为10.7μg·g-1,其中,上覆植被为发草的土样(S14)中可溶出态硅的含量最高,为26.3μg·g-1,河边沉积土(S7)数值最低,为3.90μg·g-1。通过对比可以看出,有植被覆盖的土壤样品可溶出态硅的平均含量为13.2μg·g-1,高于裸露土壤可溶出态的平均含量(8.24μg·g-1),如,有地衣覆盖的红土(S8)和裸露红土(S9)可溶出态硅的值分别为19.5和6.85μg·g-1,上覆植被为发草的溪边土(S14)中可溶出态硅的含量为26.3μg·g-1,这些站位都比邻近地区无植被的 S12(5.94μg·g-1)和 S13(4.43μg·g-1)土样含量高;由此可见,植被对可溶出态硅的含量有显著影响。一般而言,植物出现后会提高裸地的风化速率[31-32],即所谓的生物加速风化作用。在植物的作用下,岩石的风化速率得到提高,这便使得土壤中可溶出态硅含量增加。另外,极地地区降雨较少,土壤颗粒较小的流动性也使得土壤中可溶出态硅容易在植被根系附近积累,提高其含量。

图2 各土壤样品可溶出态硅的含量Fig.2.The content of soluble silica in soil samples in the Ny-Ålesund, Arctic
2.2 生物硅
土壤样品中生物硅的含量如图3所示。调查区域内,土样中BSi含量(以下均以mg Si·g-1计)平均值为7.56 mg·g-1,最大值出现在S4站位(山坡砾土)中,为9.97 mg·g-1,最小值出现在S2站位(山坡粘土),为5.60 mg·g-1。邻近站点土壤样品中BSi含量差别不大,如 S5(7.65 mg·g-1)、S6(7.80 mg·g-1)、S7(7.52 mg·g-1)、S12(7.82 mg·g-1)、S13(8.06 mg·g-1)和 S14(7.12 mg·g-1)。不过,有地衣覆盖的红土(S8)和裸露的红土(S9)的BSi含量相近,但是有植被覆盖的柳下土(S10)和发草土(S14)BSi含量比这些地区高,这可能表明高等植物(如北极柳和发草)体内硅含量较低等植物地衣高;在北极柳和发草植株死亡分解后,体内残留的植硅体存留于土壤中,导致BSi含量比地衣土高。另外,溪边土壤S12、S13、S14和河流土S5、S6、S7中BSi含量相当,这些样品均为河流冲刷携带而来的土壤,这说明河流影响的同一地区BSi含量相差不大,这很大程度上是由于河流对物质输送过程中的归一化作用,使得不同来源的BSi经河水混合表现出相类似的性质。
表2为不同纬度地区BSi含量的比较,可以看出,不同纬度地区的BSi含量不同。在热带地区,马永贝(刚果)(Mayombe,Congo)的一个热带雨林中,表层土壤 BSi含量均值为 2.5 mg·g-1[2],亚马逊三角洲表层土壤 BSi含量为 2.0 mg·g-1[16],两者数值相近,均低于本研究地区BSi的含量,这很大程度上是由于热带地区生物硅循环速率快,侵蚀强度高的缘故[2]。在温带干旱地区,恒续林地BSi平均值为 7.0 mg·g-1,放牧林为 4.6 mg·g-1,放牧草场为 5.6 mg·g-1,耕地为 3.5 mg·g-1[33],同样比北极土壤中BSi含量要低,另外,这几种土壤中生物硅含量的差异从一定程度上体现了土壤利用方式(即人类活动)对生物硅的影响,显然,极地地区较轻的人类活动干扰使得BSi容易在土壤中累积。在温带湿润的湿地地区,如卡罗莱纳州北部开普菲尔河(Cape Fear)河口湿地表层土壤中BSi含量均值为4.0 mg· g-1[34],中国黄河口湿地BSi均值为9.9 mg·g-1[35],比利时斯海尔德(Schelde)河口湿地 BSi含量为 4.4 mg·g-1[36],除黄河口湿地外,其他两个湿地中BSi含量也比北极土壤低。Strufy等[5]对瑞典北部的姆多斯国家公园(Muddus National Park)的寒带湿地进行研究,发现其BSi含量高达51.3 mg·g-1,这一数值远远高于其他已有数据报道地区的BSi含量。这也从一定程度上说明极地地区的BSi的含量要高于温、热带地区,表明寒带地区较低的温度与较少的地表径流量使得BSi容易在土壤中被保存下来。表2中还列出了各研究地区土壤的上覆植被;植被为酸角树、芦苇、莎草的土壤生物硅含量较其他土壤高,也从一定程度上说明了植被种类的不同对生物硅含量高低具有显著的影响。另外,与海洋沉积物相比,本研究区域陆地表层土壤中BSi含量并不高,如安哥拉海盆、几内亚海脊等海区沉积物中 BSi含量介于 22-54 mg·g-1[37]。 在寒带海区沉积物中,南大西洋毛德海隆(Maud Rise)地区 BSi含量为 504 mg·g-1[37]。 显然,土壤或者沉积物中生物硅的含量受到其上植物和浮游植物生物量、物质周转速率、迁移和转化等因素的综合影响,并表现出一定的规律性;而这个规律性在陆地上与植物对硅酸盐风化速率的影响[31-32]研究是一致的,即植物的出现往往会提高区域土壤中硅酸盐风化速率,致使土壤中活性硅和具有潜在活性的硅的含量升高。

表2 不同纬度地区生物硅含量比较Table 2.Comparison of biogenic silica in different latitudes
2.3 颗粒有机碳、颗粒有机氮
土壤样品中颗粒有机碳(POC)与颗粒有机氮(PON)的含量如图4所示。研究区域内,PON平均值为1.43 mg·g-1,最大值为 9.84 mg·g-1,出现在S4(山坡位置)土样中,最小值为0.11 mg·g-1,出现 S1(山顶腐殖土)土样中。POC平均值为13.7 mg·g-1,最大值为 69.3 mg·g-1,同样出现在S4土样中,最小值为1.00 mg·g-1,出现在S1土样中。土样中POC和PON显著正相关(R2=0.970,n=14,p<0.01)。另外,这14个土壤样品 C/N值均在7-33之间,其中,S13站位(溪流河道,冲蚀土)C/N值尤为高,为32.2,表明土壤有机质主要是来源于陆地高等植物,生物硅形态分析的结果也有力地证明了这一点。对土样中的BSi与POC、PON做相关性分析,还可以得出,BSi与 POC(R2=0.372,n=14,p<0.01)、PON(R2=0.383,n=14,p<0.01)呈显著正相关(图5),这也进一步证实土壤中BSi的来源及含量与有机质含量有一定关系,三者具有相似的来源。另外,BSi与POC和PON拟合的曲线的截距均为负值,这表明BSi较后者更容易被保存下来。值得一提的是,S4站位在采样期间并无植被覆盖,但其BSi含量和POC与PON的含量却是最高的。在邻近S4站位的其他采样点中,硅与碳含量也普遍较高,S4的高值很大程度上是由于早期植被留存的结果,极地地区较轻的人类活动干扰以及较弱的物质流动性使得生物成因的碳与硅容易在土壤中累积,并造成局部高值。

图3 各土壤样品生物硅的含量Fig.3.Content of soluble silica in soil samples in the Ny-Ålesund, Arctic
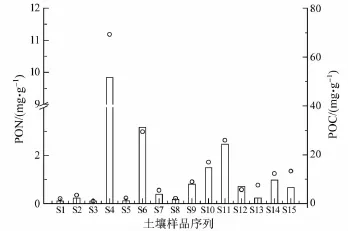
图4 土壤样品中POC和PON含量Fig.4.The contentof particle organic carbon and particle organic nitrogen in soil samples

图5 土壤样品中BSi与POC(a)和PON(b)的相关关系Fig.5.Correlations between BSiand POC/PON in soil samples
2.4 植物中植硅体的形态特征
图6为北极植物发草茎叶中植硅体的形态特征。由图可以看出,发草茎叶中的植硅体形态种类丰富,包括鞍形、骨形、帽形、扇形、齿形、尖形、圆形、平滑棒形、刺棒形、哑铃形等,大小在10-100μm之间。图7为发草根中的植硅体类型,其种类与茎叶中植硅体相似,大小在10-75μm之间。发草植物中的植硅体表现出了早熟禾亚科植物不同器官中发育的植硅体的外形轮廓特征,如占大多数的帽形、齿形、圆形的植硅体,及其内部硅质颗粒组成的泡囊等。王永吉等[30]曾指出,齿形和帽形硅酸体是反映寒冷气候的代表性类型。
苔藓中以平滑棒形植硅体为主,大小在20-100μm之间,尖形、帽形和哑铃形的植硅体只占少数(图8)。这是由于植硅体的形状依赖于原植物细胞的形状和细胞之间的空隙[30]的结果。表3为发草和苔原植物中各种形态的植硅体所占个数百分比,其中,发草中植硅体形态较为丰富,以帽形、齿形、圆形、平滑棒形、刺棒形、哑铃形等为主要形态,而苔藓中植硅体形态较为单一,以平滑棒形为主,只有少数帽形、刺棒形等形态特征,且数量较少,这体现了高等植物与低等植物的差异,其可能在地表硅循环中发挥着不同的作用,即高等植物对硅循环的影响可能明显高于低等植物。

图6 北极发草茎叶中植硅体的形态特征Fig.6.Morphology of the phytolith in the stems and leaves of Deschampsia Beauv

图7 北极发草根中植硅体的形态特征Fig.7.Morphology of the phytolith in the roots of Deschampsia Beauv

图8 苔藓中植硅体的形态特征Fig.8.Morphology of the phytolith in Andreaea
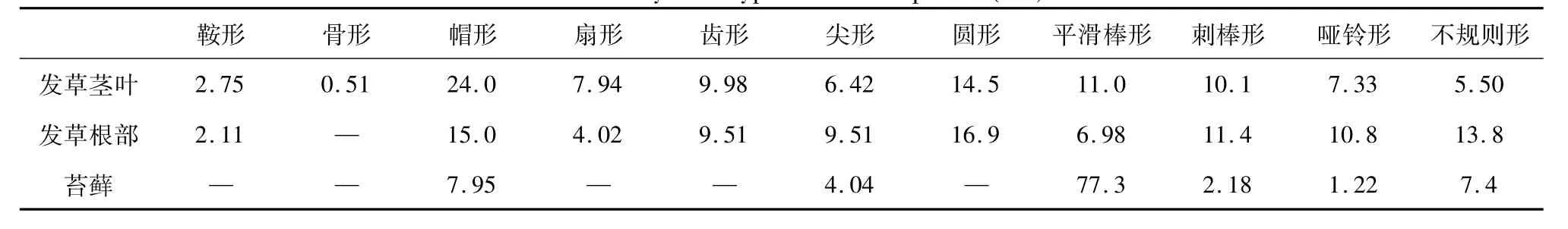
表3 苔原植物中植硅体类型统计表(%)Table 3.Phytolith types in tundra plants(%)
2.5 土壤中的生物硅形态特征
选择上覆植被为发草的土样S14(记为S-Ⅰ),以及无植被的混合土样(记为S-Ⅱ)提取植硅体。土壤中生物硅主要由植硅体和硅藻组成;其中,有植被覆盖的土壤中植硅体形态主要有平滑棒形、尖形、齿形、三角形、帽形、圆形、扇形、刺棒形及藻类(表4),大小在10-100μm之间,这与发草植株中的植硅体形态特征是相似的。无植被覆盖的土壤提取到的植硅体数量较少,形态较为单一,但藻类较多,且多为羽纹硅藻纲。这说明土壤中的植硅体主要是来源于其上植物,而不同区域间流动性较低,几乎不存在跨区域输送现象。另外,北极区夏季河流及表层土解冻,形成泥泽,可为藻类生长提供有利条件,使得藻类也成为生物硅的来源之一。
通常来说,硅与陆地植物的生长发育息息相关[5],而生物硅便是由生物生理作用所形成的无定形硅。据统计,陆地生态系统所产生的生物硅库都非常巨大,在 200 ×1012mol·a-1左右[5],是地表和水体硅循环中最主要的参与者,也是地表、海洋生物地球化学中重要的一环。近年来的研究还证实,尽管物理、化学风化过程起到基础性的作用,但生物过程才是控制陆地硅循环及产出的主要过程[9]。这主要是因为植物能加快岩石的风化,其对风化速率提高的程度应在一个量级以内[38]。正是由于地表生物过程的巨大影响,高等植物产生的植硅体是土壤中最能代表生物硅的硅质颗粒,其往往是控制河流溶解硅和生物硅输出的主要因素。遗憾的是,本次调查中没有采集到河流水样,这使得我们无法得到生物硅输送的基础数据。不过,从目前来看,无论是生物硅还是可溶出态硅的含量,植物生长都是控制其含量和组成的重要因素,其很有可能随地表径流汇入近海,对近海硅循环产生一定的影响,值得进一步关注。

表4 土壤中生物硅的类型统计表(%)Table 4.Biogenic silica composition in soil samples(%)
3 结论
(1)北极中国黄河站站基表层土壤中颗粒有机碳的含量为 1.00-69.3 mg·g-1,平均值为13.7 mg·g-1;颗粒有机氮含量为0.11-9.84mg·g-1,平均值为1.43 mg·g-1;可溶态硅含量为3.90-26.3 μg·g-1,平均值为10.7 μg·g-1;生物硅含量为5.60-9.97 mg·g-1,平均值为7.56 mg·g-1,且其含量与土壤颗粒态有机碳和颗粒有机氮呈正相关关系,表明三者有相似来源。
(2)北极表层土壤生物硅比温带地区生物硅含量要高。植物样品中植硅体形态以帽形、圆形、平滑棒形、齿形、刺棒形、哑铃形等形态为主,大小在10-100μm之间。土壤样品中的植硅体主要来源于植物,形态主要有平滑棒形、尖形、齿形、三角形、帽形、圆形、扇形及刺棒形,另外硅藻也是北极土壤中生物硅的组成部分,其中裸露土壤中藻类(羽纹纲硅藻)占生物硅的多数(93.3%)。
(3)对比发现,有植被覆盖的土壤中,土壤有机碳、可溶出态硅、植硅体形态丰富度和生物硅含量明显比其他无植被覆盖或者寡植被区土壤高,这表明陆地植物生长对该地区土壤硅循环存在显著影响,值得进一步关注。
1 Wedepohl K H.The composition of the continental crust.Geochimica et Cosmochimica Acta, 1995, 59(7):1217-1232.
2 Alexandre A,Meunier JD,Colin F,etal.Plant impacton the biogeochemical cycle of silicon and related weathering processes.GeochimicaetCosmochimica Acta,1997, 61(3):677-682.
3 Kelly E F, Chadwick O A, Hilinski T E.The effect of plants onmineral weathering.Biogeochemistry,1998,42(1-2):21-53.
4 Hinsinger P,BarrosO N F,BenedettiM F,etal.Plant-induced weathering ofa basaltic rock:Experimentalevidence.Geochimica etCosmochimica Acta,2001, 65(1):137-152.
5 Struyf E,Mörth CM,Humborg C,etal.An enormousamorphous silica stock in borealwetlands.Journal of Geophysical Research:Biogeosciences(2005-2012),2010,115(G4),doi:10.1029/2010JG001324.
6 Conley D J.Terrestrial ecosystems and the global biogeochemical silica cycle.Global Biogeochemical Cycles, 2002, 16(4):68-1-68-8.
7 Tréguer P, Nelson D M, van Bennekom A J, et al.The silica balance in the world ocean:a reestimate.Science, 1995, 268(5209):375-379.
8 Cary L,Alexandre A,Meunier JD,etal.Contribution of phytoliths to the suspended load of biogenic silica in the Nyong basin rivers(Cameroon).Biogeochemistry, 2005, 74(1):101-114.
9 Derry L A,Kurtz A C,Ziegler K,etal.Biological control of terrestrial silica cycling and export fluxes to watersheds.Nature,2005,433(7027):728-731.
10 Farmer V C,Delbos E,Miller JD.The role of phytolith formation and dissolution in controlling concentrationsof silica in soil solutionsand streams.Geoderma, 2005, 127(1-2):71-79.
11 Meunier JD, Guntzer F, Kirman S, et al.Terrestrial plant-Si and environmental changes.Mineralogical Magazine,2008,72(1):263-267.
12 冉祥滨,于志刚,臧家业,等.地表过程与人类活动对硅产出影响的研究进展.地球科学进展,2013,28(5):577-587.
13 车宏.黄河口三角洲湿地及邻近海域硅的分布特征与影响因素.青岛:国家海洋局第一海洋研究所,2013.
14 Schelske C L.Biogeochemical silicamass balances in Lake Michigan and Lake Superior.Biogeochemistry, 1985, 1(3):197-218.
15 Ran X B,Yu ZG,Yao Q Z,et al.Silica retention in the Three Gorges Reservoir.Biogeochemistry,2013,112(1-3):209-228.
16 DeMaster D J,Knapp G B,Nittrouer CA.Biological uptake and accumulation of silica on the Amazon continental shelf.Geochimica et Cosmochimica Acta, 1983, 47(10):1713-1723.
17 Correll D L, Jordan T E, Weller D E.Dissolved silicate dynamics of the Rhode riverwatershed and estuary.Estuaries,2000,23(2):188-198.
18 Hou L J,Liu M,Xu SY,etal.Distribution and accumulation of biogenic silica in the intertidal sediments of the Yangtze Estuary.Journal of Environmental Sciences, 2008, 20(5):543-550.
19 Carbonnel V,Lionard M,Muylaert K,et al.Dynamics of dissolved and biogenic silica in the freshwater reaches of a macrotidal estuary(The Scheldt, Belgium).Biogeochemistry, 2009, 96(1-3):49-72.
20 李宏亮,陈建芳,刘子琳,等.北极楚科奇海和加拿大海盆南部颗粒生物硅的粒级结构.自然科学进展,2007,17(1):72-78.
21 冉莉华,陈建芳,金海燕,等.白令海和楚科奇海表层沉积硅藻分布特征.极地研究,2012,24(1):15-23.
22 沙龙滨.格陵兰西部海域1200年以来硅藻记录及古气候、古海冰重建.上海:华东师范大学,2012.
23 姜珊.过去3000年南北极典型地区生态环境变化的沉积记录及对比.合肥:中国科学技术大学,2012.
24 Humlum O,Instanes A,Sollid JL.Permafrost in Svalbard:a review of research history,climatic background and engineering challenges.Polar Research,2003,22(2):191-215.
25 Fox R L, Silva JA, Younge O R, et al.Soil and plant silicon and silicate response by sugarcane.Soil Science Society of America Journal,1967,31(6):775-779.
26 Khalid R A,Silva JA.Residual effects of calcium silicate in tropical soils:II.Biological extraction of residual soil silicon.Soil Science Society of America Journal, 1978, 42(1):94-97.
27 DeMaster D J.The supply and accumulation of silica in themarine environment.Geochimica et Cosmochimica Acta,1981,45(10):1715-1732.
28 Fischer G,Gersonde R,Wefer G.Organic carbon,biogenic silica and diatom fluxes in themarginalwinter sea-ice zone and in the Polar Front Region:interannual variationsand differences in composition.Deep Sea Research Part II:Topical Studies in Oceanography,2002,49(9-10):1721-1745.
29 Reddy K R, DeLaune R D.Biogeochemistry ofWetlands:Science and Applications.Fresno:Witerwoof Inc,2008.
30 王永吉,吕厚远.植物硅酸体研究及应用.北京:海洋出版社,1993.
31 Moulton K L,Berner R A.Quantification of the effect of plants on weathering:Studies in Iceland.Geology,1998,26(10):895-898.
32 Moulton K L, West J, Berner R A.Solute flux and mineralmass balance approaches to the quantification of plant effects on silicate weathering.A-merican Journal of Science, 2000, 300(7):539-570.
33 Clymans W, Struyf E, Goovers G, et al.Anthropogenic impact on biogenic Si pools in temperate soils.Biogeosciences Discussions,2011,8:4391-4419.
34 Norris A R, Hackney C T.Silica content of a Mesohaline Tidal Marsh in North Carolina.Estuarine, Coastal and Shelf Science, 1999, 49(4):597-605.
35 刘森,冉祥滨,车宏,等.黄河口湿地土壤中生物硅的分布与植硅体的形态特征.土壤,2014,46(5):886-893.
36 Struyf E,Van Damme S,Gribsholt B,et al.Biogenic silica in freshwatermarsh sediments and vegetation(Schelde estuary,Belgium).Marine E-cology Progress Series, 2005, 303:51-60.
37 Müller P J, Schneider R.An automated leachingmethod for the determination of opal in sediments and particulatematter.Deep Sea Research Part I:Oceanographic Research Papers, 1993, 40(3):425-444.
38 Drever J I.The effect of land plants on weathering rates of silicateminerals.Geochimica et Cosmochimica Acta, 1994, 58(10):2325-2332.
