联觉的发展与习得
2015-01-23李佳源
李佳源
(西华师范大学管理学院, 四川南充 637009)
1 引言
在日常生活中有一小部分正常人(约4%)声称当他们听音乐时可以看到颜色, 当听或说词汇时嘴里会感觉到味觉, 当看到字母或数字会感觉到某种特定颜色, ……这种奇异的现象, 就被称为联觉(synaesthesia)。联觉是由一种感觉引起另一种感觉的心理现象(黄希庭, 2007; Simner, 2012)。引起联觉的诱发刺激称为诱发物(the inducer), 诱发刺激引发的感觉称为伴随体验(the concurrent),如联觉可以是被嘴里的味觉、数字、词语、时间等不同的诱发物引起, 可能激发像声音、形状、颜色、味觉、气味、触摸感觉等等不同的伴随体验。自1812年Sachs第一次对联觉现象进行描述以来(Jewanski, Day, & Ward, 2009), 超过60多种联觉已经被报道(Novich, Cheng, & Eagleman, 2011)。
近年来, 随着认知心理学和神经生理学的不断发展, 对联觉的研究呈“井喷”之势, 从以往单一的行为实验转向利用神经成像技术探究联觉者的脑机制。不少研究者使用神经成像技术(fMRI和PET)发现颜色联觉者大脑的V4区有增加的白质(Rouw, Scholte, & Colizoli, 2011)和灰质(Hänggi,Beeli, Oechslin, & Jäncke, 2008; Weiss & Fink,2009; Banissy et al., 2012), V1区和V2区也有增加的灰质(Rouw & Scholte, 2010; Jäncke, Beeli, Eulig,& Hänggi, 2009), 这些研究显示出联觉者与非联觉者存在大脑结构的差异, 这种差异不仅仅存在于某一特定大脑区域, 而且还存在于不同的大脑区域(Rouw et al., 2011)。那么联觉者与非联觉者这种大脑结构上的差异是由什么原因造成的呢?一种回答就是先天基因遗传造成了这种大脑结构的差异, 所有的人都是天生的联觉者, 但大多数人随后由于神经突触的修剪而丢失了这种跨通道连接(Spector & Maurer, 2009; Wagner & Dobkins,2011)。然而目前的基因遗传研究却无法解释儿童的联觉关联(Synaesthetic associations)为什么比成人更不稳定, 为什么这些联觉关联需要经过数年之后才逐渐地变得稳定(Simner, Harrold, Creed,Monro, & Foulkes, 2009), 也不能解释为什么会在诱发物与伴随物之间制造某种特定的关联, 比如为什么一个联觉者将字母Y与淡黄白色相关联(Ramachandran, Hubbard, & Butcher, 2004), 而另一个联觉者却将它与淡黄绿色相关联(Spector &Maurer, 2009)。基于此, 一些研究者提出联觉具有后期发展习得的可能(Deroy & Spence, 2013;Rothen & Meier, 2014; Watson, Akins, Spiker,Crawford, & Enns, 2014), 并通过一系列的行为实验试图证实这种可能性。那么联觉是否具有发展的可能?如果联觉具有后期发展的可能, 那么非联觉者能否通过关联训练成为联觉者?这已经成为近期联觉研究中最具有争议性的问题, 得到了联觉研究者广泛的关注, 因此本文以常见的颜色联觉以及其他类型联觉为例, 以新的视角总结前人对联觉的发展、训练以及可习得性的相关研究成果, 分析与讨论非联觉者是否可以通过训练产生联觉体验以及能否成为真正的联觉者, 期望唤起更多研究者对联觉的新思考。
2 联觉的发展
联觉具有家族遗传性, 至少40%的联觉者有一个具有联觉的直系亲属(Barnett et al., 2008), 不同类型的联觉能在同一个家族中出现, 不同类型联觉可能与基因相关。Bargary和Mitchell (2008)显示了在直接控制皮层连接的基因中的突变是如何导致联觉的, 并且解释了在轴突指导、边界形成或修剪中的差异如何在获得联觉体验的临近区域之间创造直接、前馈的连接。Asher等人(2009)做了听觉—视觉联觉的第一个建立在43个多重家庭的全基因组连锁研究, 结果显示听觉—视觉联觉是与多重而非唯一的基因位点相关(2q24, 5q33,6p12, 12p12)。而Tomson等人(2011)将DNA关联分析和严格的分型方法相结合来研究颜色序列联觉(颜色周日、月份、字母和数字)的基因机制, 他们发现颜色序列联觉是与在16q12.2-23.1区域的一个基因相关, 并给出了6个备选基因:GABARAPL2、NDRG4、PLLP、KATNB1、CIAPIN1以及GNAO1。
联觉的基因遗传性表明基因倾向在决定联觉中起到重要的作用, 但并没有表明基因遗传就是决定联觉的唯一因素。某些联觉者可能对诱发刺激的低层次视觉形式敏感, 而某些联觉者则不一样, 比如一个看见数字“6”体验到粉红色颜色的联觉者, 他可能将他的这种粉红色体验描述成一种漂浮在外部空间中不同位置的一块斑点——在他的头部左边30厘米和前边1米处, 这种描述就会是一个直接的感觉体验描述, 数字“6”与粉红色相关联, 而不是概念“6”。然而同样的联觉者无论看见书面词“六”、骰子上的6个点、数词6、竖的6个指头还是作为一个思想的6, 他都体验到粉红色的颜色, 那么这种联觉就应是概念诱发, 因为它涉及对诱发刺激的具体特征进行一种抽象, 概念“6”与粉红色相关联, 而不是数字“6”。
但问题在于, 如何来评定联觉者的粉红色体验是数字“6”的视觉形式诱发还是概念词“6”诱发呢?一种评定方式是看不同视觉形式的数字(或字母)是否会诱发相同的伴随体验, 如果一个联觉者描述了两种或两种以上的数字(或字母)的不同视觉形式都是一种颜色, 那么就可以断定不是视觉形式诱发了联觉, 而是语言或概念范畴诱发了联觉。在大多数字母-颜色联觉中, 视觉上不同的形式能诱发相同的颜色, 只要它们是属于同一语言范畴, 即‘a,a, A, a’和‘a’可能都被体验为红色(Simner, 2012)。在序语言人格化联觉中, 语言序列, 比如字母、数字、周日和月份会导致一种人格化类型和性别, 比如字母a可能是一个忙碌的母亲, 而数字9可能是一个忠实的丈夫(Simner &Holenstein, 2007)。而在词汇-味觉联觉中, 词汇诱发事物味觉体验, 这些体验要么可能是感觉的,即在嘴里对气味的感知, 要么可能是概念的, 即心理关联到一种食物类型, 比如, 联觉者A面对词汇“John”时, 在嘴里体验到食物的气味, 然而联觉者B面对着词汇“Shoulder”时, 却体验到食物的概念(Simner, 2012)。这些事实说明这些联觉的伴随体验可能依赖于诱发物(比如字母、数字、词汇等)的概念范畴, 而不依赖于它们的视觉特征。当联觉第一次被建立时, 对视觉特征敏感可能出现在儿童的早期阶段, 然而一旦这些关联被形成(即诱发物和它们相应的伴随体验关联), 那么联觉体验就可能对视觉特征不敏感(Simner &Haywood, 2009)。因而联觉可能涉及儿童后期形成的高层次认知, 比如字母-颜色联觉, 由字母体验到颜色的成人联觉者可能被这个字母的概念范畴(语言或概念范畴)所激发, 即使最初的字母与颜色的关联在儿童时期被建立在视觉特征基础上。
为了实时地探究在儿童时期联觉的发展,Simner等(2009)对英国21所小学年龄在6~7岁的600名儿童进行了字母-颜色联觉的行为测试, 经过字母-颜色一致性的评定程序, 确立了47个具有潜在联觉的儿童。一年之后, 再用相同的程序对这47个儿童进行测试, 有8个儿童依然具有字母-颜色一致性, 这8个儿童被视作是真正的联觉者, 而其他39个则被看作具有较高记忆力的非联觉者。他们同时还发现这8个字母-颜色联觉者习得了平均6.4个新的字母-颜色关联, 即6~7岁的儿童联觉者有平均10.5个稳定的字母-颜色关联,而在同样测试中的8~9岁的儿童联觉者却有平均16.9个稳定的字母-颜色关联。Simner和Bain(2013)在4年之后又用相同的程序对10~11岁的这些儿童进行了测试, 看这些联觉者在4年后是否还会有一致性关联表现, 是否还会有一致性的字体-颜色关联的进一步增加, 是否一些儿童的联觉会消失。测试结果显示之前的8个联觉者中还有5个符合联觉的标准, 而且这5个联觉者的字母-颜色关联进一步地增加到了平均25.7个。西姆纳等人的研究一方面表明了一些儿童在早期具有联觉体验特征, 而随着年龄的增加会消失,许多儿童早期可能都是潜在的联觉者, 然而随着之后的神经突触的修剪使得大部分儿童的跨通道连接分离, 而一小部分的儿童由于修剪不充分而继续保有跨通道连接(Maurer & Mondlach, 2005;Hochel & Milán, 2008), 即使是同卵双胞胎也会出现一个具有联觉, 而另一个没有发展出联觉的情况(Smilek et al., 2002)。另一方面, 他们的研究发现儿童联觉的一致性关联会随着年龄的增加而增加, 这个发现与儿童习得字母表与数字的发展阶段相吻合, 由于字母-颜色联觉是与被习得的语言符号紧密联系在一起的, 它不可能是在婴儿时期就已经如此存在, 虽然神经影像数据表明了在婴儿时期感知语言的外侧裂语言区域已经形成(Dehaene-Lambertz et al., 2006), 但仅仅是在具体的语言词汇被习得之后, 用于驱动字母-颜色联觉的表征类型才能被建立。
随后, Meier, Rothen和Walter (2014)又探究了439名年龄跨度为18~91岁成年人的字形-颜色联觉一致性的发展情况。他们使用相关性和准实验的方法检测了一致的字形-颜色关联以及评定了其与年龄的关系, 结果显示少量一致性的字形-颜色关联会有与年龄相关的下降。由于之前的研究发现在儿童和少年时期一致性的字形-颜色关联会增加(Simner et al., 2009; Simner & Bain,2013), 与这个结果一起表明字形-颜色联觉体验的变化呈现一种类似于许多其它认知功能的倒U形曲线(Shing & Lindenberger, 2011; Weiermann &Meier, 2012)。同时他们也评定了色谱的宽度, 结果显示最常见颜色词的出现, 比如红、蓝、绿, 不会随着年龄增长而发生变化, 黄色、橙色、品红色等二次色随着年龄增加很少会出现, 然而褐色、灰色和白色等非情感颜色则会随着年龄增加会经常出现。这一结果表明字形-颜色关联的一致性具有与年龄相关的下降并不是一个简单的、偶然的结果, 它们可能是与成年人发展过程中颜色感知与识别的系统变化有关。随着年龄的增加,一些字形不再诱发联觉颜色, 特定的关联也可能会丢失, 这说明在联觉的关联一致性变化中会涉及到低层次的感觉加工与高层次的认知加工。随着年龄的增加, 二次色的加工(黄色、橙色、品红色)会减少, 使得它们的感知会变得越来越弱, 甚至于会消失, 在诱发物与伴随物之间的关联很可能向柔和色(褐色、灰色和白色)变化, 最后这个关联会随着时间的推移而消失, 比如, 一个黄色伴随体验可能向白色衰退, 而再以后这种白色伴随体验则可能完全消失。这种变化与重组与记忆类似(Nader & Hardt, 2009), 一些字形-颜色关联由于衰退感觉加工会被重建, 而这些重建的关联也可能会丢失。
所有这些研究表明在婴儿时期的联觉不是一个最终的结果, 相反地, 它仅仅是一个发展的起点。儿童时期联觉关联的一致性和宽度会随着年龄增大而增加, 在青年时期联觉的关联一致性会变得相对稳定, 而联觉关联的宽度则会达到最高峰值, 然而在中年、老年时期, 联觉关联的一致性和宽度会逐渐下降, 甚至有些联觉关联会完全消失(图1)。因此, 联觉关联不是完全被基因所决定,不少联觉关联后期可能会发展变化, 而联觉关联的这种发展变化可能会涉及多种因素的相互作用,比如基因、大脑、环境与文化等, 它们可能共同影响联觉的发展变化(Rouw & Scholte, 2010; Ronga,Bazzanella, Rossi, & Iannetti, 2012)。
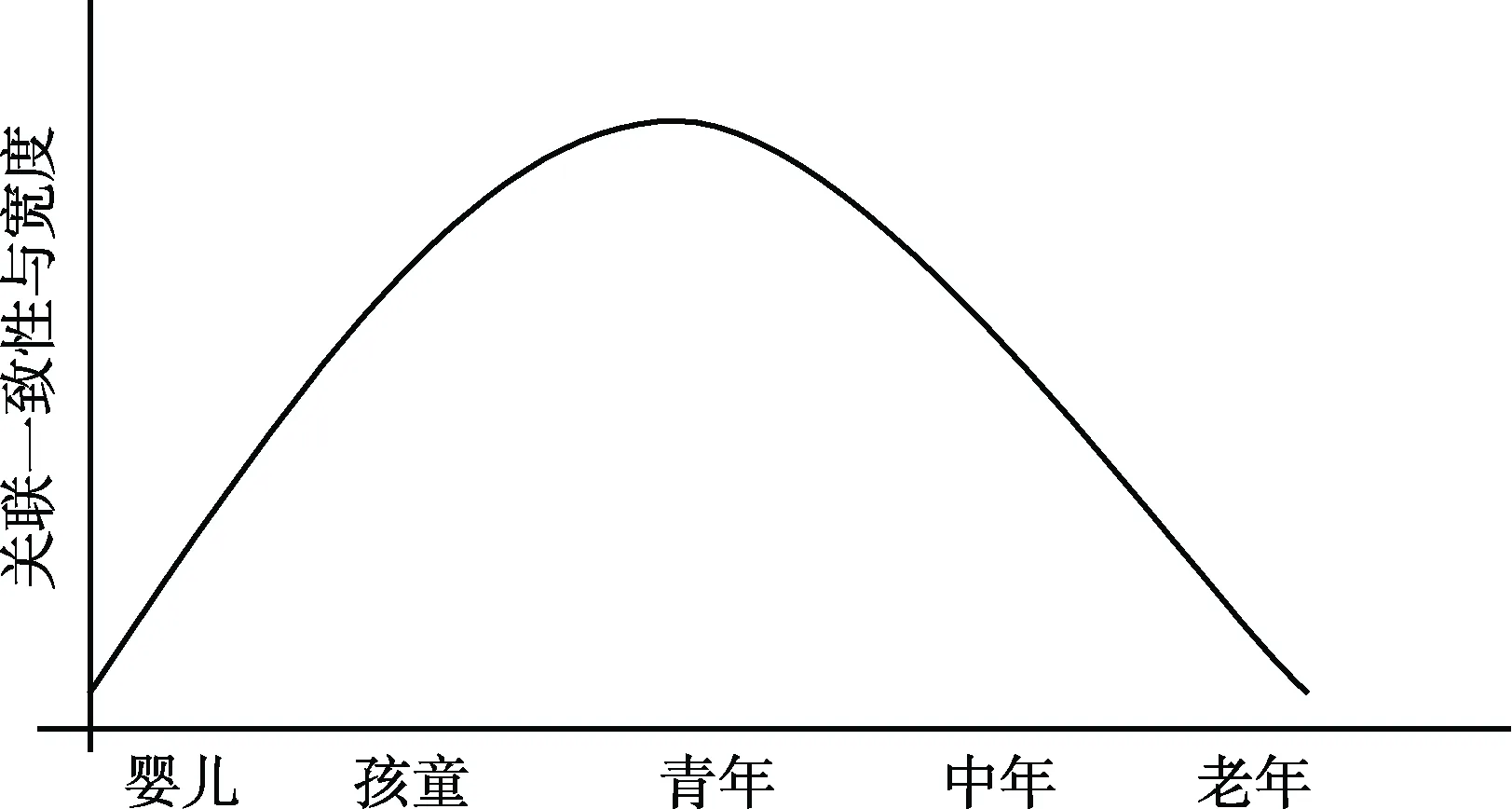
图1 联觉关联一致性发展变化情况
3 联觉可训练习得吗?
既然联觉可以发展变化, 那么就会出现与此相关的、令人感兴趣的问题:(1)非联觉者经过适当的配对关联训练能否具有联觉者的行为?(2)非联觉者经过适当的配对关联训练能否变成真正的联觉者?以往有两种测试方法用于证实真实的联觉者, 一种是联觉Stroop测试, 其常被用作证实真实联觉者的一种诊断标记(Dixon, Smilek, &Merikle, 2004; Ward, Li, Salih, & Sagiv, 2007)。在联觉Stroop测试中为联觉者提供具有颜色的字母,一些字母的颜色与联觉体验到的颜色是一致的,而一些字母颜色则与联觉体验到的颜色不一致。联觉者需要说出每个字母的颜色, 当一些字母颜色与联觉者体验到的颜色不一致时, 联觉者说出字母的颜色会比一致时慢, 甚至有时也会说错。另一种是联觉条件反射测试, 在联觉条件反射测试中为参与者(包括联觉者和非联觉者)在习惯化、条件反射和消退三个不同的阶段呈现不同颜色的幻灯片, 在适应阶段随意地将所有颜色(绿、蓝、白、红及黄)的幻灯片为参加者呈现多次, 而白色的幻灯片标有能诱发联觉颜色体验的一个字母D(或其他的字母); 在条件阶段几张作为条件刺激的幻灯片, 即蓝色幻灯片总是被惊叫之声伴随,按照一个固定的伪随机序呈现给参与者, 而其他颜色的幻灯片则不伴有惊叫之声。在消退阶段,蓝色幻灯片和白色幻灯片用交替的方式反复呈现多次(见图2)。通过皮肤电导反应检测发现, 在条件阶段的所有参加者对蓝色幻灯片都有惊叫之声诱发的皮肤电反应, 而对于标有字母D的白色幻灯片只有联觉者才会显示由惊叫之声诱发的反应, 非联觉者则没有(Bechara et al., 1995)。

图2 联觉条件反射测试(Bechara et al., 1995)
到目前为止, 为数不多的行为研究试图通过对非联觉者进行配对关联训练来诱发联觉(见表1)。这些研究针对颜色联觉类型, 较早期的两个(Kelly, 1934; Howells, 1944)是声音-颜色配对关联训练, 而近期的训练是与人后期习得的语言、概念、技能密切相关的字母(数字、字形)-颜色和游泳-颜色配对关联训练。这些研究使用了不同的训练模式, 比如反复试验, 要求当在呈现一个音调或字母时, 被试需从几个颜色斑点之间选择,并对被试的每一次选择进行反馈(Howells, 1944;Brang, Kanai, Ramachandran, & Coulson, 2011);被试以自己的速度阅读带有颜色字母的小说(Colizoli, Murre, & Rouw, 2013); 完成对带有颜色字母的快速视觉搜索任务(Kusnir &Thut, 2012);甚至于为了将特定数字与颜色相关联而进行催眠后指令(Cohen-Kadosh, Henik, & Walsh, 2009)。训练时间长度也从几分钟(Nunn et al. 2002)到20多天(Kelly, 1934)不等。大部分研究都进行了联觉Stroop测试, 其中3个研究不仅仅进行了联觉Stroop测试, 而且还进行了联觉条件反射测试(Meier &Rothen, 2009; Rothen, Wantz, & Meier, 2011; Rothen et al., 2013)。只有3个研究在实验中没有使用联觉Stroop测试, 在Kelly (1934)的研究中使用的是内省报告, 将“如果音调能够唤起被试者自动的感觉或颜色影像, 那么一个真实的条件反射就已经被建立”作为内省的标准。Cohen-Kadosh等人(2009)采用了数字觉察任务(Digit Detection Task)测试, 在数字觉察任务中, 被试需要觉察一个色彩背景上的消色差的数字是否与数字指定的颜色一致或不一致。而Brang等人(2011) 采用了建立在认知加工而非感觉加工基础上的情景启动效应测试。
目前有两个研究声称训练后的非联觉者有联觉体验(Howells, 1944; Cohen-Kadosh et al., 2009)(见表1)。一个来自于早期Howells (1944)的研究,这个研究对8个非联觉者进行了声音-颜色配对关联训练, 通过联觉Stroop测试发现训练后的非联觉者具有联觉Stroop效应, 据此认为非联觉者训练后具有联觉体验。另一个来自于Cohen-Kadosh等人(2009)的研究, 这个研究使用催眠后指令方式对12名非联觉者进行数字-颜色配对关联训练,除了收集现象的报告之外, 这个研究还要求被试完成一个数字觉察任务。这个研究的结果表明,在现象的报告层面, 在催眠后暗示组的被试都报告了与先天联觉者相匹配的现象, 是一致的、自动的、日常生活中发生的, 而通过数字觉察任务的完成情况分析后认为催眠后暗示组的被试具有一种新的联觉体验。
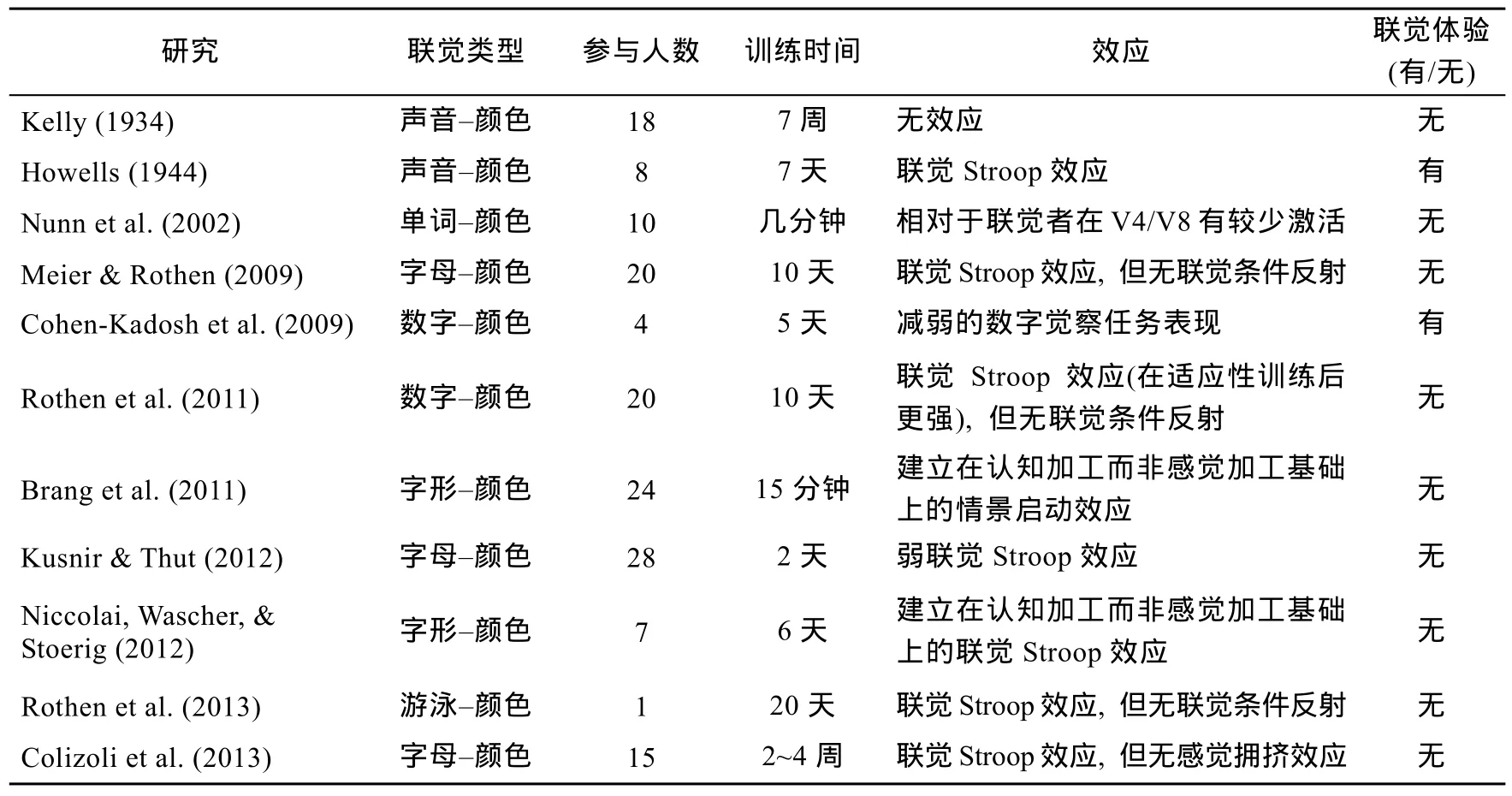
表1 习得训练与效应
这两个研究没有给出直接的证据或强有力的证据来证实非联觉者经过配对关联训练后能成为真正的联觉者。虽然在这两个研究中的非联觉者都报告有颜色体验, 颜色体验都具有类似于真正联觉体验的表现, 比如自动性和一致性, 单向而非双向的, 但问题在于现象报告是主观的、内省性的(Deroy & Spence, 2013), 相似的行为表现并不能充分地说明这一现象就是联觉。配对关联后非联觉者出现类似于联觉者的表现, 极大可能是配对关联训练强化了记忆, 一种感觉的体验总是唤起先前与此种感觉相对应的一种记忆搜索。比如在字体-颜色联觉中, 如果在最初的体验中, 字母Y是与淡黄白色相关联, 那么在经过关联强化训练之后, 字母Y与淡黄白色的关联被编码于记忆中, 随后当单独体验字母Y时, 将会唤起此种关联的记忆, 此种记忆最终将导致对这些颜色体验的提取。如果是这样, 那么训练得到的类似联觉的行为就仅仅是一种关联记忆的加工过程(Prescott, 2012), 不是一种真正的联觉行为。Howells (1944)的研究虽然发现非联觉者在联觉Stroop任务中有类似于联觉者的行为, 然而联觉Stroop效应仅仅是联觉的必要而非充分的条件,联觉Stroop测试对于评定一种语义关联的强度是非常有用的, 但对于评定什么是联觉体验(或什么是联觉)却是失败的(Meier & Rothen, 2009; Deroy& Spence, 2013)。真正的联觉者会同时具有联觉Stroop效应和联觉条件反射, 但从目前的研究发现联觉者具有联觉Stroop效应却不具有联觉条件反射(Meier & Rothen, 2009; Rothen et al., 2011;Rothen et al., 2013)。Cohen-Kadosh等人(2009)的研究采用了催眠后暗示的训练方式进行配对关联训练, 得到催眠后暗示诱发了类似于先天字形-颜色联觉的跨通道体验, 而对于这一结果的解释更多可能是一种心理意象, 而非是一种联觉体验,因为他在实验中给出如此指令“看这个颜色, 这是数字×的颜色, 无论你什么时候看见、思考、或想象到它, 你总是感觉它为这个颜色”, 这一指令更多可能是与催眠后心理意象的诱发相关, 易受暗示影响的个体往往具有更加生动的视觉心理意象(Crawford, 1982; Rader & Tellegen, 1987)。实际上, 在Cohen-Kadosh等人(2009)之前有一个实验报道了相似的现象(Leuba, 1940), 在催眠的条件下, 将一个被试的手臂摩擦与防腐油的味道配对关联, 后来这个被试的手臂只要摩擦时, 他就会有防腐油的嗅觉意象, 而在这个实验中将这种现象描述为是一种条件反射的心理意象。对于这种催眠后暗示到底诱发的是一种心理意象还是一种联觉体验, 还需要我们将来通过使用神经影像来测试大脑特定领域的激活情况, 比如在字母-颜色联觉中, 大脑V4区有相应的激活, 但在一种心理意象中, 大脑V4区通常不会显示一种激活。
目前9个研究得出非联觉者经过配对关联训练后不会产生联觉体验(Kelly, 1934; Nunn et al.,2002; Meier & Rothen, 2009; Rothen et al., 2011;Brang et al., 2011; Kusnir & Thut, 2012; Niccolai et al., 2012; Rothen et al., 2013; Colizoli et al., 2013)(见表1)。Kelly (1934)的研究使用的是参与者的现象报告, 非联觉者经过配对关联训练后被问是否有与声音相关联的颜色体验, 部分非联觉者报告没有任何稳定的颜色体验, 这个研究由此断定联觉体验不能训练习得。Nunn等人(2002)的研究使用fMRI测试发现经过关联配对训练后的非联觉者不像联觉者一样在V4/V8区有激活现象出现。而Brang等人(2011)的研究则使用ERP实验测试发现训练后的非联觉者有建立在认知加工基础上的情景启动效应, 而不像联觉者建立在感觉加工基础上的情景启动效应。其余的6个研究对关联配对训练后的非联觉者进行了联觉Stroop测试,Kusnir和Thut (2012)的研究发现具有弱的联觉Stroop效应, 再加上训练后的非联觉者报告没有颜色体验, 因此这个研究断定训练关联在本质上不同于联觉。Colizoli等人(2013)的研究发现训练后的非联觉者具有联觉Stroop效应, 但无感觉拥挤效应, 而无感觉拥挤效应说明配对训练后的关联不具有联觉的感觉性质。Niccolai等人(2012)的研究也发现训练后的非联觉者具有联觉Stroop效应, 然而他们通过使用ERP测量发现联觉与训练后获得的关联具有不同的神经加工; Meier和Rothen (2009)、Rothen等(2011)以及Rothen等人(2013)既进行了联觉Stroop测试, 还进行了条件反射测试, 他们都发现训练后的非联觉者会显示出联觉Stroop效应, 然而都没有显示出一种条件反射。
从这些研究证据可以看到, 训练关联与联觉存在不少的差异, 虽然训练关联也会显示联觉Stroop效应, 然而不会显示出一种联觉的条件反射, 而联觉既会有联觉Stroop效应, 也会有联觉的条件发射。训练关联与联觉有不同的加工基础,训练关联主要涉及高层次的认知加工, 而联觉还涉及低层次的感觉加工。另一方面, 目前大部分训练研究仅仅是在配对关联训练后就对关联的一致性进行测试, 没有为这种关联的长期一致性提供证据, 比如在半年或更长时间之后, 再对这些非联觉者进行关联一致性测试, 看看他们是否还具有这种关联一致性。只有一个研究在配对关联训练后6个月再对这些非联觉者进行了关联一致性测试, 然而遗憾的是, 这个研究发现在6个月之后, 这些非联觉者既没有那种训练的配对关联,也没有显示一种联觉Stroop效应(Colizoli et al.,2013)。更为重要的是, 当前唯一一个对单词-颜色关联训练后的非联觉者使用fMRI进行的大脑激活情况测试表明, 相对于真正的联觉者, 这些训练后的非联觉者大脑V4区没有被激活(Nunn et al., 2002)。虽然这个研究还不足以说明训练后的非联觉者就不能变成真正的联觉者, 因为它的训练时间仅仅只有几分钟, 而已有神经影像研究测试了儿童阅读技能的发展(McCandliss, Cohen, &Dehaene, 2003)以及视运动技能——杂技对大脑白质束变化的影响(Scholz, Klein, Behrens, &Johansen-Berg, 2009), 发现对较长期的学习阅读作出反应的腹侧视觉区域有重组现象以及一些较长时间的复杂视运动——杂技训练能够改变健康成年人大脑中的白质束结构, 这说明某种技能经过较长时期的训练可能会改变大脑的结构。但是联觉者与非联觉者的根本性区别就在于两者在联觉体验期间大脑激活情况存在差异, 两者大脑结构存在差异(Rouw et al., 2011), 只要能再显示经过较长时间关联训练后的非联觉者大脑的特定区域是否会出现对应的激活, 大脑结构是否会发生一些改变, 那么就能最终证实关联训练后的非联觉者能否成为真正的联觉者。
因此, 虽然训练后的非联觉者具有一些类似于联觉者的表现, 但现有的研究证据越来越多地显示出训练关联与联觉存在不少差异, 训练后的非联觉者可能无法变成真正的联觉者。如果关联训练后的非联觉者不能变成真正的联觉者, 那么我们又如何解释在一些联觉的发展过程中获得了一些新的刺激物与伴随物的配对关联呢?这是我们需要进一步解释的问题, 一种可能就是联觉者的大脑神经结构中天生就具有负责低层次感觉以及高层次语言和概念加工的大脑神经区域, 在联觉的发展过程中, 由于联觉者不断与外在的文化、语言环境交互, 比如习得新的语言、概念等,而这些新习得的语言、概念就会激活负责高层次语言和概念加工的大脑神经区域而产生新的联觉关联。那么在联觉的大脑神经结构中有没有这样一个进行高层次语言和概念加工的区域呢?这样一个最可能的区域是前颞叶(the anterior temporal lobe), 目前的颜色联觉研究发现, 联觉颜色体验激活了高层次的大脑皮层区域, 而真实颜色则没有(Van Leeuwen, Petersson, & Hagoort, 2010)。前颞叶是进行语言、概念高层次认知加工的核心区域(Patterson, Nestor, & Rogers, 2007), 并且前颞叶对于促成概念对象与它们的标准颜色之间的关联起到重要的作用, 而这种关联与联觉中刺激物与伴随物之间的关联类似(Chiou &Rich, 2014)。
当然, 我们有一种将训练关联纳入到联觉中的简单办法, 那就是改变我们以往对于联觉的界定。心理学对联觉的界定基本上以跨通道的感觉特征为标准, 即一种感觉诱发另一种感觉的现象,以这种标准对联觉进行界定, 那么语言与文学中的隐喻、美学中的通感以及这里的训练关联都可纳入联觉的范畴。虽然心理学将联觉定义为一种感觉诱发另一种感觉的现象, 但是心理学的联觉研究视角却是将联觉严格限定为少数人拥有且具有遗传性的, 这样又会将语言与文学中的隐喻、美学中的通感以及这里的训练关联排除在外。一些学者为了消解这样的矛盾, 提出将联觉分为强联觉和弱联觉(Martino & Marks, 2001; Sakamoto& Utsumi, 2009; Nakamura, Sakamoto, & Utsumi,2010; 刘思耘, 2012), 强联觉就是少数人才有且具有遗传性, 弱联觉则是指出现在语言中的一种隐喻、文学与美学中的通感以及训练关联现象。如果按照这样的做法, 训练关联就能被纳入到联觉中, 然而这是一种过于草率的做法, 如果训练关联是一种联觉, 那么它们就应该具有相同的本质, 特别是要具有相同的大脑神经机制, 可我们现在的证据越来越多地表明它们两者可能存在本质的不同, 它们可能就是两种不同的心理现象。
4 小结与展望
联觉不是完全被基因遗传所决定的, 能够在后期发展变化, 这就意味着联觉的形成会受到个体发展过程中多种环境因素的影响, 比如语言、文化等。认识到联觉是发展的, 将会使我们对于联觉的研究视角被进一步的拓展, 一个直接相关、令人感兴趣的问题就是:什么样的因素能够形成联觉体验?虽然目前的研究已经显示了基因遗传会发展出与联觉相关的行为和大脑结构, 然而一些研究也表明通过训练或者其它的环境影响可能会导致行为和大脑结构的变化(Draganski et al., 2006; Colcombe et al., 2006)。这样就有一种形成联觉的可能:无需基因遗传, 而直接可通过训练习得。
目前一些研究通过配对关联训练后, 发现非联觉者具有类似于联觉者的表现, 也发现训练后的非联觉者与真正的联觉者具有一定的差异。我们通过对这些研究证据的对比分析表明, 虽然训练后的非联觉者具有类似于联觉者的行为, 但是现有的证据越来越多地显示联觉可能无法直接通过训练习得, 训练后的非联觉者可能无法成为真正的联觉者。由于现有的训练研究还存在不少有待改进的地方, 比如训练方式还比较单一、训练时间比较短、脑区结构研究少等, 因此我们还不能够完全确定联觉就无法训练习得。
而要最终证实联觉能否通过训练习得, 将来的训练研究还需要关注如下几个方面:(1)配对关联训练后的非联觉者与联觉者的大脑机制有何相同或不同?联觉者具有特定的大脑结构, 如果非联觉者经过关联训练后能变成联觉者, 那么也应该具有相似的大脑结构, 因此将来的研究需要使用神经成像技术确定两者的大脑机制的异同。(2)强的记忆(Radvansky, Gibson, & McNerney, 2011;Meier & Rothen, 2013a)、强的创造力(Ward, Thompson-Lake, Ely, & Kaminski, 2008)以及强的意象(Barnett & Newell, 2008; Meier & Rothen, 2013b)会伴随有联觉, 它们可能单独或一起引起某种联觉, 因此将来的训练研究对象可以不局限于普通的非联觉者, 而可以考虑具有这些特征的非联觉者,看看他们经过关联训练后能否成为联觉者。(3)目前训练研究的对象都集中在成年人, 很少涉及儿童群体, 而儿童的生理可塑性更强(Rothen & Meier,2014), 儿童时期的训练相对而言可能更有希望导致联觉, 因此将来的训练研究对象还需要考虑儿童非联觉者。
黄希庭. (2007).心理学导论(p.293). 北京: 人民教育出版社.
刘思耘. (2012). 强联觉的认知加工模型及其脑机制.心理科学进展, 20(4), 514-522.
Asher, J. E., Lamb, J. A., Brocklebank, D., Cazier, J. B.,Maestrini, E., Addis, L., … Monaco, A. P. (2009). A whole-genome scan and fine-mapping linkage study of auditory-visual synesthesia reveals evidence of linkage to chromosomes 2q24, 5q33, 6p12, and 12p12.American Journal of Human Genetics, 84, 279-285.
Banissy, M. J., Stewart, L., Muggleton, N. G., Griffiths, T.D., Walsh, V. Y., Ward, J., & Kanai, R. (2012). Grapheme-color and tone-color synesthesia is associated with structural brain changes in visual regions implicated in color,form, and motion.Cognitive Neuroscience, 3(1), 29-35.
Bargary, G., & Mitchell, K. J. (2008). Synaesthesia and cortical connectivity.Trends in Neurosciences, 31(7), 335-342.
Barnett, K. J., Finucane, C., Asher, J. E., Bargary, G., Corvin,A. P., Newell, F. N., & Mitchel, K. J. (2008). Familial patterns and the origins of individual differences in synaesthesia.Cognition, 106(2), 871-893.
Barnett, K. J., & Newell, F. N. (2008). Synaesthesia is associated with enhanced, self-rated visual imagery.Consciousness and Cognition, 17, 1032-1039.
Bechara, A., Tranel, D., Damasio, H., Adolphs, R., Rockland,C., & Damsio, A. R. (1995). Double dissociation of conditioning and declarative knowledge relative to the amygdale and hippocampus in humans.Science, 269, 1115-1118.
Brang, D., Kanai, S., Ramachandran, V. S., & Coulson, S.(2011). Contextual priming in grapheme-color synesthetes and yoked controls: 400 msec in the life of a synesthete.Journal of Cognitive Neuroscience, 23, 1681-1696.
Chiou, R., & Rich, A. N. (2014). The role of conceptual knowledge in understanding synesthesia: Evaluating contemporary findings from a “hub-and-spokes”perspective.Frontiers in Psychology, 5, 105.
Cohen-Kadosh, R., Henik, A., & Walsh, V. (2009). Synaesthesia: Learned or lost?Developmental Science, 12(3),484-491.
Colcombe, S. J., Erickson, K. I., Scalf, P. E., Kim, J. S.,Prakash, R., …. Kramer, A. F. (2006). Aerobic exercise training increases brain volume in aging humans.The Journals of Gerontology Series A: Biological Sciences and Medical Sciences, 61, 1166-1170.
Colizoli, O., Murre, J. M. J., & Rouw, R. (2013). Pseudosynaesthesia through reading books with colored letters.PLoS One, 7, e39799.
Crawford, H. J. (1982). Hypnotizability, daydreaming styles,imagery vividness, and absorption: A multidimensional study.Journal of Personality and Social Psychology, 42,915-926.
Dehaene-Lambertz, G., Hertz-Pannier, L., Dubois, J., Mériaux, S., Roche, A., Sigman, M., & Dehaene, S. (2006).Functional organization of perisylvian activation during presentation of sentences in preverbal infants.Proceedings of National Academy of Science of the United States of America, 103, 14240-14245.
Deroy, O., & Spence, C. (2013). Training, hypnosis, and drugs: Artificial synaesthesia, or artificial paradises?Frontiers in Psychology, 4, 660.
Dixon, M. J., Smilek, D., & Merikle, P. M. (2004). Not all synaesthetes are created equal: Projector versus associator synaesthetes.Cognitive Affective & Behavioral Neuroscience,4, 335-343.
Draganski, B., Gaser, C., Kempermann, G., Kuhn, H. G.,Winkler, J., Büchel, C., & May, A. (2006). Temporal and spatial dynamics of brain structure changes during extensive learning.The Journal of Neuroscience, 26, 6314-6317.
Hänggi, J., Beeli, G., Oechslin, M. S., & Jäncke, L. (2008).The multiple synaesthete ES-Neuroanatomical basis of interval-taste and tone-colour synaesthesia.Neuroimage,43(2), 192-203.
Hochel, M., & Milán, E. G. (2008). Synaesthesia: The existing state of affairs.Cognitive Neuropsychology, 25(1), 93-117.
Howells, T. H. (1944). The experimental development of color-tone synesthesia.Journal of Experimental Psychology,34, 87-103.
Jäncke, L., Beeli, G., Eulig, C., & Hänggi, J. (2009). The neuroanatomy of grapheme-color synesthesia.European Journal of Neuroscience, 29(6), 1287-1293.
Jewanski, J., Day, S. A., & Ward, J. (2009). A colorful albino: The first documented case of synaesthesia by Georg Tobias Ludwig Sachs in 1812.Journal of the History of the Neurosciences, 18, 293-303.
Kelly, E. L. (1934). An experimental attempt to produce artificial chromaesthesia by the technique of the conditioned response.Journal of Experimental Psychology,17, 315-341.
Kusnir, F., & Thut, G. (2012). Formation of automatic letter-colour associations in non-synaesthetes through likelihood manipulation of letter-colour pairings.Neuropsychologia, 50, 3641-3652.
Leuba, C. (1940). Images as conditioned sensations.Journal of Experimental Psychology, 26, 345-351.
Martino, G., & Marks, L. E. (2001). Synesthesia: Strong and weak.Current Directions in Psychological Science, 10(2),61-65.
Maurer, D., & Mondlach, C. J. (2005). Neonatal synesthesia:A reevaluation. In L. C. Robertson & S. Sagiv (Eds.),Synesthesia: Perspectives from cognitive neuroscience(pp.193-213). New York: Oxford University Press.
McCandliss, B. D., Cohen, L., & Dehaene, S. (2003). The visual word form area: Expertise for reading in the fusiform gyrus.Trends Cognition Science, 7, 293-299.
Meier, B., & Rothen, N. (2009). Training grapheme-colour associations produces a synaesthetic Stroop effect, but not a conditioned synaesthetic response.Neuropsychologia,47, 1208-1211.
Meier, B., & Rothen, N. (2013a). Synaesthesia and memory.In J. Simner & E. M. Hubbard (Eds.),Oxford handbook of synesthesia(pp. 692-706). Oxford: Oxford University Press.
Meier, B., & Rothen, N. (2013b). Grapheme-color synaesthesia is associated with a distinct cognitive style.Frontiers in Cognitive Science, 4, 632.
Meier, B., Rothen, N., & Walter, S. (2014). Developmental aspects of synaesthesia across the adult lifespan.Frontiers in Human Neuroscience, 8, 129.
Nader, K., & Hardt, O. (2009). A single standard for memory:The case for reconsolidation.Nature Review Neuroscience,10, 224-234.
Nakamura, T., Sakamoto, M., & Utsumi, A. (2010). The role of event knowledge in comprehending synesthetic metaphors. InProceeding of the 32st Annual Meeting of the Cognitive Science Society,1898-1903.
Niccolai, V., Wascher, E., & Stoerig, P. (2012). Distinct neural processes in grapheme-colour synaesthetes and semantic controls.European Journal of Neuroscience, 36,3593-3601.
Novich, S., Cheng, S., & Eagleman, D. M. (2011). Is synaesthesia one condition or many? A largescale analysis reveals subgroups.Journal of Neuropsychology, 5, 353-371.
Nunn, J. A., Gregory, L. J., Brammer, M., Williams, S. C. R.,Parslow, D. M., Morgan, M. J., … Gray, J. A. (2002).Functional magnetic resonance imaging of synesthesia:Activation of V4/V8 by spoken words.Nature Neuroscience, 5(4), 371-375.
Patterson, K., Nestor, P. J., & Rogers, T. T. (2007). Where do you know what you know? The representation of semantic knowledge in the human brain.Nature Reviews Neuroscience, 8, 976-987.
Prescott, J. (2012). Chemosensory learning and flavour:Perception, preference and intake.Physiology & Behavior,107(4), 553-559.
Rader, C. M., & Tellegen, A. (1987). An investigation of synesthesia.Journal of Personality and Social Psychology,52, 981-987.
Radvansky, G. A., Gibson, B. S., & McNerney, M. W. (2011).Synesthesia and memory: Color congruency, von Restorff,and false memory effects.Journal of Experimental Psychology, Learning, Memory, and Cognition, 37, 219-229
Ramachandran, V. S., Hubbard, E. M., & Butcher, P. A.(2004). Synesthesia, cross-activation, and the foundations of neuroepistemology. In G. Calvert, C. Spence, & B. E.Stein (Eds.),The handbook of multisensory processes(pp.867-883). Cambridge, MA: MIT Press.
Ronga, I., Bazzanella, C., Rossi, F., & Iannetti, G. (2012).Linguistic synaesthesia, perceptual synaesthesia, and the interaction between multiple sensory modalities.Pragmatics and Cognition, 20(1), 135-167.
Rothen, N., Wantz, A. L., & Meier, B. (2011). Training synaesthesia.Perception, 40, 1248-1250.
Rothen, N., Nikolić, D., Jürgens, U. M., Mroczko-Wąsowicz,A., Cock, J., & Meier, B. (2013). Psychophysiological evidence for the genuineness of swimming-style colour synaesthesia.Consciousness and Cognition, 22, 35-46.
Rothen, N., & Meier, B. (2014). Acquiring synaesthesia:Insights from training studies.Frontiers in Human Neuroscience, 8, 109-122.
Rouw, R., & Scholte, H. S. (2010). Neural basis of individual differences in synesthetic experiences.Journal of Neuroscience, 30(18), 6205-6213.
Rouw, R., Scholte, H. S., & Colizoli, O. (2011). Brain areas involved in synaesthesia: A review.Journal of Neuropsychology, 5, 214-242.
Sakamoto, M., & Utsumi, A. (2009). Cognitive effects of synesthetic metaphors evoked by the semantic interaction.InProceeding of the 31st Annual Meeting of the Cognitive Science Society,1593-1598.
Scholz, J., Klein, M. C., Behrens, T. E. J., & Johansen-Berg,H. (2009). Training induces changes in white-matter architecture.Nature Neuroscience, 12(11), 1370-1371.
Shing, Y. L., & Lindenberger, U. (2011). The development of episodic memory: Lifespan lessons.Child Development Perspectives, 5, 148-155.
Simner, J., & Holenstein, E. (2007). Ordinal linguistic personification as a variant of synesthesia.Journal of Cognitive Neuroscience, 19, 694-703.
Simner, J., Harrold, J., Creed, H., Monro, L., & Foulkes, L.(2009). Early detection of markers for synaesthesia in childhood populations.Brain, 132, 57-64.
Simner, J., & Haywood, S. L. (2009). Tasty non-words and neighbours: The cognitive roots of lexical-gustatory synaesthesias.Cognition, 110, 171-181.
Simner, J. (2012). Defining synaesthesia.British Journal of Psychology, 103, 1-15.
Simner, J., & Bain, A. E. (2013). A longitudinal study of grapheme-color synesthesia in childhood: 6/7 years to 10/11 years.Frontiers in Human Neuroscience, 7, 603.
Smilek, D., Moffatt, B. A., Pasternak, J., White, B. N.,Dixon, M. J., & Merikle, P. M. (2002). Synaesthesia: A case study of discordant monozygotic twins.Neurocase, 8,338-342.
Spector, F., & Maurer, D. (2009). Synesthesia: A new approach to understanding the development of perception.Developmental Psychology, 45(1), 175-189.
Tomson, S. N., Avidan, N., Lee, K., Sarma, A. K., Tushe, R.,Milewicz, D. M., … Eagleman, D. M. (2011). The genetics of colored sequence synesthesia: Suggestive evidence of linkage to 16q and genetic heterogeneity for the condition.Behavioural Brain Research, 223, 48-52.
Van Leeuwen, T. M., Petersson, K. M., & Hagoort, P. (2010).Synaesthetic colour in the brain: Beyond colour areas. A functional magnetic resonance imaging study of synaesthetes and matched controls.Plos One, 5(8), 2074.
Wagner, K., & Dobkins, K. R. (2011). Synaesthetic associations decrease during infancy.Psychological Science, 22,1067-1072.
Ward, J., Li, R., Salih, S., & Sagiv, N. (2007). Varieties of grapheme-colour synaesthesia: A new theory of phenomenological and behavioural differences.Consciousness and Cognition, 16, 913-931.
Ward, J., Thompson-Lake, D., Ely, R., & Kaminski, F.(2008). Synaesthesia, creativity and art: What is the link?British Journal of Psychology, 99, 127-141.
Watson, M. R., Akins, K. A., Spiker, C., Crawford, L., &Enns, J. T. (2014). Synesthesia and learning: A critical review and novel theory.Frontiers in Human Neuroscience, 8, 98-113.
Weiermann, B., & Meier, B. (2012). Incidental sequence learning across the lifespan.Cognition, 123, 380-391.
Weiss, P. H., & Fink, G. R. (2009). Grapheme-colour synaesthetes show increased grey matter volumes of parietal and fusiform cortex.Brain, 132(Pt 1), 65-70.
