记忆的生存加工优势是自然选择的结果:来自跨年龄比较和繁衍情境的证据*
2015-01-23唐卫海刘玉霞白学军刘希平
唐卫海 谢 斯 刘玉霞,2 白学军,3 刘希平
(1天津师范大学教育科学学院, 天津 300387) (2山东省潍坊市景芝镇党委, 山东 262119)(3天津师范大学心理与行为研究院, 天津 300074)
1 前言
通过进化, 人类的记忆系统得以使人们更容易记住与适应相关的信息, 尤其是与生存相关的信息。当对学习材料进行“生存加工”(survival processing)时, 人们的记忆成绩明显优于其他加工条件下的记忆成绩, 这种现象被称为生存加工优势效应(survival-processing advantage effect) (Nairne,Thompson, & Pandeirada, 2007)。
生存加工优势效应的研究范式比较简单。第一步, 学习阶段。研究者请被试对一系列学习项目进行评定。生存条件下, 通常让被试想象自己陷于缺乏生存资料的逃生大草原中, 他们需要寻找到稳定的水源和食物来源, 同时还要保护自己免受猛兽的袭击, 目的是为了确保自己生存。被试需要对每个项目与这一生存情境的相关性作出评估(Nairne et al., 2007); 在其他条件下, 要求被试对材料进行其他深加工, 如愉悦度评定等。第二步, 干扰阶段。请被试完成一些无关活动, 如顺序回忆一串数字(Burns, Hart, Griffith, & Burns, 2013)等。第三步, 提取阶段。要求被试对所评定的材料进行提取, 比较生存加工与其他条件下的记忆效果的差别。
已有研究结果(Kang, McDermott, & Cohen,2008; Nairne, Pandeirada, & Thompson, 2008; Otgaar et al., 2011; Röer, Bell, & Buchner, 2013; Weinstein,Bugg, & Roediger; 2008)发现, 生存加工与多种控制条件相对照都表现出记忆上的优势。这些控制条件在可能影响记忆编码的因素上与生存加工条件进行匹配, 如新异性、唤醒度、媒体曝光度(Kang et al., 2008; Röer et al., 2013), 图式加工(Weinstein et al., 2008)和祖先优势(Nairne, Pandeirada, Gregory,& Van Arsdall, 2009)等因素。同时, 研究者也发现当使用图片代替词汇作为评定材料时, 这种效应仍然存在(Otgaar, Smeets, & Van Bergen, 2010), 它似乎不受刺激呈现形式的影响。因此, 生存加工本身被解释为一种独特而又强大的记忆编码程序(于睿,毛伟宾, 贾喆, 2011)。
然而也有一些研究似乎对生存加工做为一种强大的编码程序这一观点提出挑战。如 Otgaar和Smeets (2010)选取 DRM 词表作为研究材料, 词表中的每个单词都与一个不出现在词表中的“关键诱饵”相关联, 这是一种可以有效引发错误记忆的方法。Otgaar等的研究结果发现使用这种词表, 不仅发现错误回忆率有所增长, 并且就净准确性而言并没有出现生存加工的优势效应。其中, 净准确性 =正确回忆率/(正确回忆率+关键诱饵的错误回忆率)。这一结果暗示了生存加工优势效应也存在着一定的边界条件。
另有研究者(Butler, Kang, & Roediger III, 2009)指出生存加工优势效应可能是学习材料与加工情境之间的一致性导致的。他们认为生存加工优势效应的产生可能是由于所选的单词与生存相关程度大, 更容易同生存联系起来, 因此印象更深刻, 所以在生存情境下加工与生存相关单词的回忆成绩会优于其他条件下加工相同单词的回忆成绩。研究者(Butler et al., 2009)让被试评估词表与草原生存情景或银行抢劫情境的关联程度。前人研究结果已证实, 当单词为随机选择并且与所选情境无实质关联时, 生存情境下的单词回忆成绩优于抢劫情境下的回忆成绩, 即出现典型的生存加工优势效应(Kang et al., 2008)。但Butler等经事先评定将词表分为三类, 分别为与生存高相关但与抢劫低相关的词表、与抢劫高相关但与生存低相关的词表以及与两种情境都无关的词表。结果显示出一致性效应,即在生存(抢劫)情境下加工的生存(抢劫)相关单词的回忆成绩优于生存(抢劫)情境下加工的抢劫(生存)相关单词的回忆成绩, 同时对所有词表来说,没有发现生存加工与抢劫加工下回忆成绩的差异。但后来的研究者对这一问题进行验证, 都证实了生存加工优势效应的存在, 加工条件的一致性只在一定程度上影响着记忆的生存加工优势(Nairne &Pandeirada, 2011; 毛伟宾, 于睿, 李春, 2013)。尽管生存加工优势效应存在一定的限制因素, 但仍不可否认记忆的生存优势是生存加工独特编码的结果。
研究者用自然选择(natural selection)理论来解释生存加工优势效应。自然选择理论来源于进化心理学。它认为自然界会发生很多变化, 处于狩猎—采集环境中的祖先的大脑为了适应这种变化也会做出相应的改变, 从而更容易解决环境中出现的问题, 而这种改变会遗传到现代人身上, 并使人们拥有一套处理狩猎—采集环境中问题的认知适应工具(toolkit of cognitive adaptations) (Tooby & Cosmides,1992)。这说明生存加工优势效应的产生是自然选择的作用。依据自然选择理论的假设, 那些人类祖先所经受的生存威胁比那些在进化历史过程中出现相对较晚的威胁更容易引起人们的注意(Öhman& Mineka, 2001), 即生存加工应该对祖先优势更为敏感。Weinstein等人(2008, 实验 2)的研究设置了生存加工的两种实验条件, 即草原生存情境和城市生存情境, 分别对应了原始生存背景和现代生存背景。其中草原生存情境沿用了Nairne等(2007)所用材料, 而城市生存情境将草原生存情境中的草原替换为城市、猛兽替换为袭击者。研究者假设, 由于草原求生更接近于进化适应环境中所面临的问题,所以草原生存情境下所评估单词的回忆成绩应该优于城市生存情境下所评估单词的回忆成绩, 结果也恰恰验证了这一假设。同时这一结论也得到其他研究的验证(Nairne & Pandeirada, 2010)。这支持了生存加工优势效应来源于自然选择。
总体而言, 自然选择理论认为祖先因自然选择压力而形成的心理机制具有继承性, 对生存相关的信息记忆成绩好也是心理机制的一个方面; 人类存在很多由祖先遗传下来的适应器, 这些适应器用来解决特定性的适应性问题。适应性问题包括生存、繁衍等, 并且生存与繁衍息息相关。然而已有研究很少对生存加工优势效应的自然选择理论解释直接进行证明, 如果自然选择理论可以解释生存加工优势效应, 则生存加工优势效应应该与个体的经验没有关系。同时如果自然选择理论成立, 则自然选择的, 除了生存加工还有其他与种族延续、人类发展有关的特性, 也应该表现出加工优势。研究设计了两个实验, 实验1考察了不同年龄个体间的生存加工优势效应的差异; 实验2试图探讨在人类发展中与生存处于类似地位的繁衍等是否也存在加工优势。
2 实验1 年龄与生存加工优势效应
2.1 研究目的
考查不同年龄条件下生存加工优势效应的变化。
2.2 研究假设
如果自然选择理论的解释成立, 则生存加工优势效应在不同年龄被试间的差异量应该没有明显差别; 如果随着被试年龄的增长, 其生存加工优势效应有所变化, 则说明后天的成长也是制约生存加工优势效应的因素。
2.3 方法
2.3.1 被试
随机选取8岁、10岁、12岁、21岁和64岁年龄组被试共300名(男107名, 女193名), 每个年龄组60名。各组年龄平均数和标准差分别为8.73 ± 0.37、10.32 ± 0.52、12.60 ± 0.56、21.43 ± 1.77、64.07 ± 4.63。所有被试裸眼或矫正视力正常。每个被试在实验结束后均获得一份小礼品。
2.3.2 材料
从Battig and Montague norms (2004)分类词表中挑选100个词, 这些词在相应类别中出现的比例≥0.5, 随后将这些词翻译成汉语。随机选择未参加正式实验的15名大学生对上述100个词汇进行与远古狩猎和狩猎比赛情境关联度的评定。评定采取5点量表法, 其中 1表示完全不相关, 5表示完全相关。对评定结果整理后, 从中选出与两种情境的评定值大致相等的40个词, 每个等级8个。从5个等级中分别选取一个词用于练习, 其余 35个词作为正式实验的材料。
2.3.3 实验设计
采用2(情境:远古狩猎和狩猎比赛)×5(年龄:8岁、10岁、12岁、21岁和64岁)被试间设计, 每个年龄组的一半被试被随机分配到远古狩猎情境,另一半被试被随机分配到狩猎比赛情境。因变量为正确回忆率、生存效应量、错误回忆率、词语的评定值和评定时间。学习材料随机呈现。
2.3.4 实验程序
实验程序采用 E-prime编制, 由电脑呈现, 被试距电脑屏幕约50 cm。正式实验前有练习, 让被试熟悉实验程序。整个实验大约持续30 min。正式实验共包括以下三部分:
(1)学习阶段。对词语和情境进行关联度评定阶段, 指导语改编自Nairne等(2009)实验2中所用指导语, 如下:
远古狩猎:请想象你生活在远古的大草原上,作为部落成员, 你需要为部落捕获猎物, 你可以捕获猛兽、小动物或者附近河流湖泊里的鱼类。你作为捕猎者需要经常穿越很远的路程去追踪猎物并成功返回部落。不管条件如何, 你都要保证捕猎成功, 从而为部落的生存提供保障。下面将呈现一些词语, 你的任务是对这些词语所指代的事物与你成功狩猎并返回部落进行相关性评定。
狩猎比赛:请想象你参加一项狩猎比赛, 作为小组成员, 你的主要任务是和同伴们一起去捕获更多的猎物, 使小组取得胜利。你可以捕获猛兽和小动物, 或者附近河流或湖泊里的鱼类。你作为捕猎者需要经常穿越很远的路程去追踪猎物并成功返回比赛地点。不管条件如何, 你都要保证捕猎成功,从而为小组的胜利提供保障。下面将呈现一些词语,你的任务是对这些词语所指代的事物与你在狩猎比赛中取得胜利进行相关性评定。
每个词语呈现5 s, 评定等级为1~5, 1表示完全不相关, 5表示完全相关, 待评定等级出现在每个词语的下方, 被试需要在5 s内按键盘上的数字键做反应。如果在5 s内未作出反应, 则会呈现下一个词语。
(2)干扰阶段。让被试欣赏 30张风景图片, 每张图片呈现4 s, 整个干扰阶段持续2 min。
(3)测验阶段。在没有提前告知有测验任务的情况下, 要求被试尽可能的回忆出刚才评定过的 35个词语, 并将词语写在答题纸上, 该阶段持续10 min。
2.4 结果
被试都能顺利完成实验任务, 对实验中99%以上的目标词都进行了评估, 并且未对目标词做出评估(按键反应出现在 5 s之后)的反应数目没有组间差异, 因此在数据分析中不对未作反应的数据进行删除。
(1)远古狩猎与狩猎比赛条件下记忆效果的比较
为考查不同条件下产生的记忆效果, 对不同年龄组被试在两种情境下的正确回忆率进行了统计,具体结果见表1。
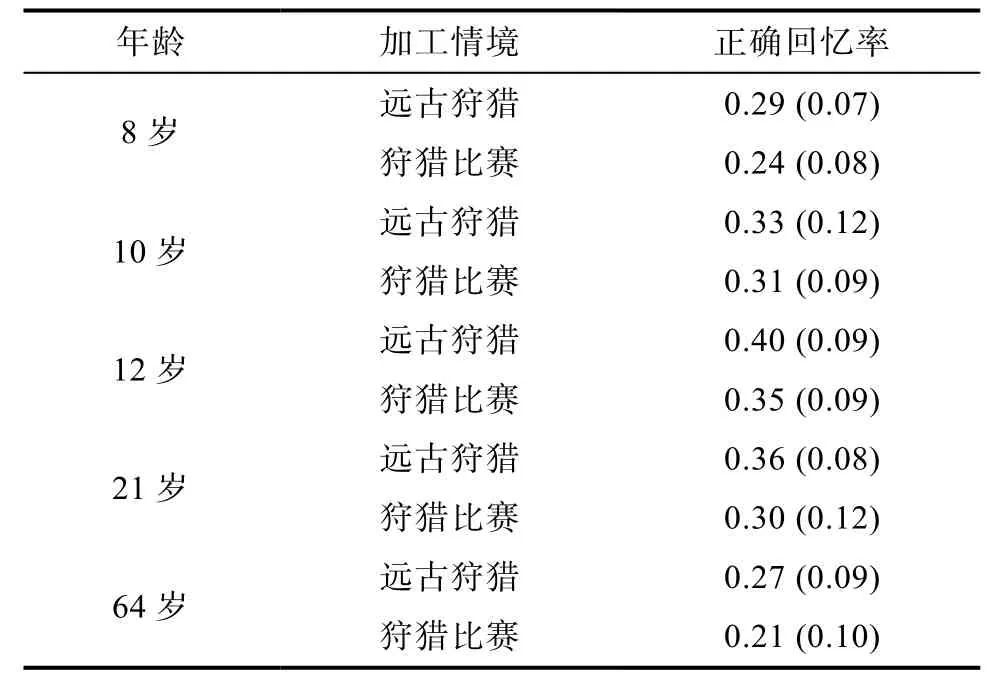
表1 各年龄组在两种情境下的正确回忆率M(SD)
Levene方差检验结果显示, 各实验处理内总方差为齐性,F(9, 290) = 1.34,p> 0.05。对不同条件的正确回忆率进行2×5的方差分析, 结果表明:情境主效应非常显著,F(1, 290) = 17.75,p< 0.01,=0.058, 说明远古狩猎情境下被试的正确回忆成绩高于狩猎比赛情境, 出现生存加工优势效应; 年龄主效应非常显著,F(4, 290) = 19.57,p< 0.01,η2p=0.213, 说明不同年龄组被试的正确回忆成绩差异显著; 情境和年龄的交互作用不显著,F(4, 290) =0.34,p= 0.851>0.05,η2p= 0.005, 说明不同年龄组被试在两种情境下正确回忆成绩的变化趋势一致。
(2)不同年龄组远古狩猎与狩猎比赛条件下回忆成绩差异的变化
各年龄组被试在总的回忆成绩上差异显著, 但这并不是本研究的关注点。研究需要关注不同年龄的被试是否都产生了等量的加工优势。在不同年龄组上, 对远古狩猎和狩猎比赛两种情境中被试回忆成绩差异的差异进行检验, 也就是检验不同的年龄组间生存优势效应量(生存优势效应量 = 生存条件下的正确回忆率-控制条件下的正确回忆率, 另见Nouchi, 2012; Olds, Lanska, & Westerman, 2014)是否存在差异。以往的研究没有比较过这种差异的差异, 为达到这种统计目的, 我们使用如下检验方法。采用SAS软件进行编程, 将不同年龄组的远古狩猎情境和狩猎比赛情境的成绩分别作为一个向量, 因本实验中每个处理都包括30个数据, 所以每种情境均为30×1的向量, 新的向量Z = (第一个向量-第二个向量) - (第三个向量-第四个向量) = (8岁组远古狩猎情境-8岁组狩猎比赛情境) - (10岁组远古狩猎情境-10岁组狩猎比赛情境), 然后对“Z的诸分量之均值是否等于 0”这一问题进行检验,也就是转化成常规的t检验, 其他9组的检验方法同上。结果显示:每两个年龄组间的生存优势效应量均没有显著差异,p> 0.05。
(3)远古狩猎与狩猎比赛条件下错误回忆率的比较
在被试的回忆词单中, 出现了“闯入词”, 即被试回忆出学习时未出现的词语。实验中计算了闯入词的回忆成绩, 即错误回忆率, 错误回忆率 = 闯入词个数⁄(闯入词个数+正确回忆个数), 对各条件下的错误回忆率进行统计, 结果如表2所示。
Levene方差检验结果显示, 各实验处理内总方差为不齐,F(9, 290) = 3.02,p< 0.05。故对被试错误回忆率Y的原始数据进行平方根转换, 得到一个新的因变量, 即新错误回忆率对其进行 Levene方差检验, 结果显示各实验处理内总方差齐性,F(9, 290) = 1.83,p> 0.05。
对被试在各实验条件下的新错误回忆率进行方差分析, 结果表明:年龄主效应极其显著,F(4,290) = 12.84,p< 0.001,= 0.150, 说明各年龄组被试的新错误回忆率差异显著。进一步多重比较结果表明, 64岁年龄组的新错误显著高于其他各组,p<0.001, 21岁年龄组的新错误率显著高于8岁组和12岁组,p< 0.05。情境主效应不显著,F(l, 290) = 0.087,p> 0.05,ηp2= 0.000, 说明被试在远古狩猎情境下的新错误回忆率与狩猎比赛情境下的新错误回忆率没有差异。年龄和情境的交互作用边缘显著,F(4,290) = 0.88,p= 0.475,ηp2= 0.012, 说明不同年龄组被试在两种情境下的新错误回忆率变化趋势不一致。简单效应分析显示, 12岁组、21岁组和64岁组的新错误回忆率表现为远古狩猎情境低于狩猎比赛情境, 而8岁组和10岁组的新错误回忆率表现为远古狩猎情境高于狩猎比赛情境。
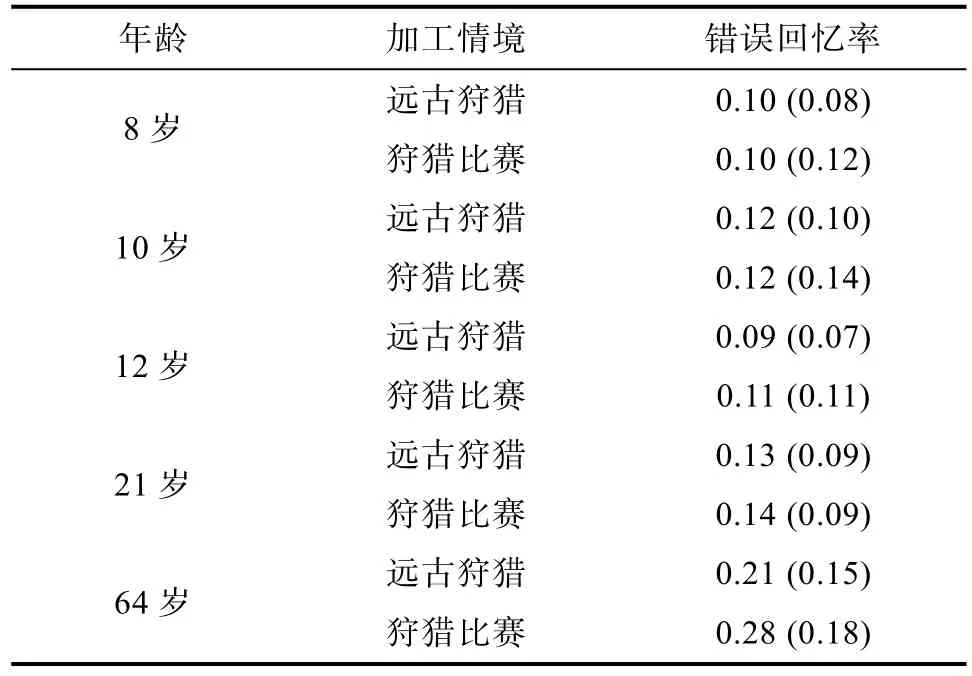
表2 各年龄组在两种情境下的错误回忆率M(SD)
(4)远古狩猎与狩猎比赛条件下词语评定值和反应时间的比较
因为词语与情境的评定值和评定时间会影响被试的记忆成绩, 所以对不同条件下词语的评定值和评定时间进行统计, 结果见表3。
由于不同年龄被试在反应能力上存在着较大的差异, 生存加工优势效应考查的是同一年龄组被试在两种情境下的表现情况, 为排除评定值和评定时间的影响, 分别对同一年龄组被试在两种情境下的评定值和评定时间进行独立样本t检验。在评定值上,t8岁组(58) = 0.47,p= 0.642,d= 0.12;t10岁组(58) =-0.12,p= 0.908,d= 0.03;t12岁组(58) = 1.10,p=0.278,d= 0.28;t21岁组(58) = 0.91,p= 0.367,d= 0.24;t64岁组(58) = 1.58,p= 0.120,d= 0.41; 在评定时间上,t8岁组(58) = -0.95,p= 0.347,d= 0.25;t10岁组(58) =-1.41,p= 0.165,d= 0.36;t12岁组(58) = 0.43,p=0.668,d= 0.11;t21岁组(58) = -0.61,p= 0.544,d=0.16;t64岁组(58) = -0.77,p= 0.445,d= 0.20。t检验结果表明, 同一年龄组内, 被试在两种情境中的评定值和评定时间没有差异。这说明前述生存加工优势效应没有受到评定等级和加工时间的制约, 具有较强的说服力。
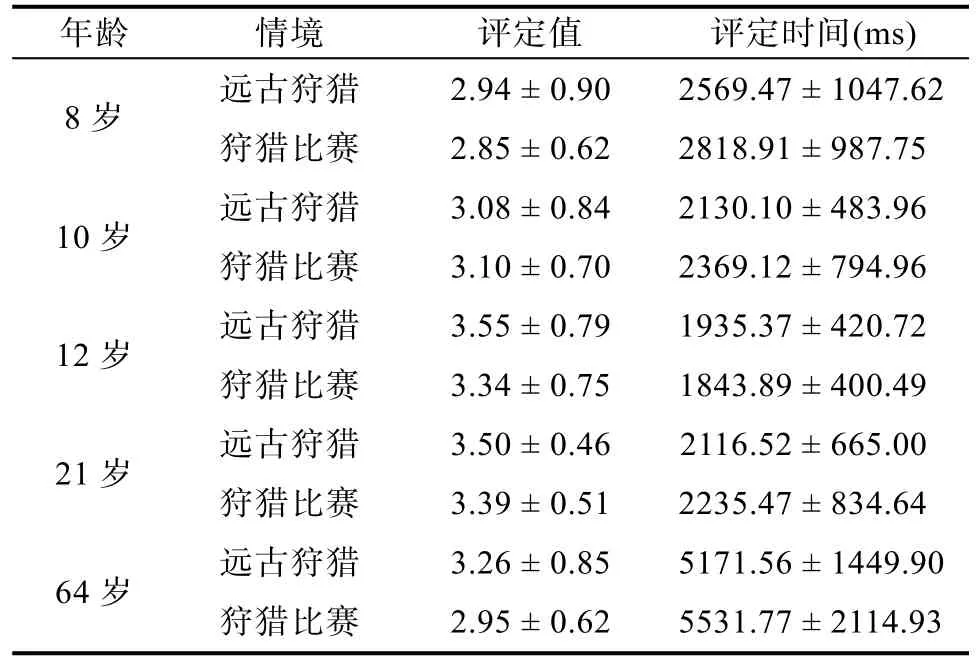
表3 两种情境中的评定值和评定时间(M ± SD)
2.5 讨论
实验1主要目的是通过考查不同年龄组被试的生存加工优势效应, 为自然选择解释提供证据。
实验结果表明, 8岁组、10岁组、12岁组、21岁组和 64岁组在远古狩猎上的回忆成绩高于狩猎比赛, 表现出生存加工优势效应。年龄与情景的交互作用不显著, 表明各年龄组间的生存优势效应量没有差异, 生存加工优势效应是自然选择的结果。但对该交互作用的power分析表明(G-power, Faul,Erdfelder, Lang, & Buchner, 2007), 现有的样本量能够探测出生存优势量的年龄差异的效力仅为0.14。所以虽然在统计结果上可以推断各年龄组间的生存优势量相等, 但是这一结论还有待于进一步验证。
有研究者认为生存加工优势其实是一种主题优势, 实验采用两种指标认为生存优势不是主题优势:(1)正确回忆率。主题优势表现为对学习材料进行某一主题的加工会提高最终的记忆成绩。本实验采用远古狩猎和狩猎比赛两种情境, 它们都围绕“狩猎”这一主题进行加工, 区别在于前者有生存因素, 结果发现远古狩猎情境下被试的回忆成绩高于狩猎比赛。(2)错误回忆率。主题优势认为, 生存优势之所以会出现是因为被试利用“生存”主题对信息进行了有效地编码或提取。而错误率可以作为主题或分类加工的指标。如果在回忆时, 被试依赖建立的结构(如, 某个主题或分类)对学习项目进行提取, 那么在回忆结果中可能会出现与主题或分类相关的干扰或错误记忆。实验中对两种情境的错误回忆率进行分析, 结果发现远古狩猎情境下被试的错误回忆率与狩猎比赛情境没有显著差异。但是一些研究(Otgaar & Smeets, 2010; Howe & Derbish, 2010)在考查错误回忆率时, 通常发现生存条件下的错误回忆率高于控制条件, 与本研究结果不一致。可能的原因在于上述研究所用实验材料均为DRM词表或类别词表, 而这些材料通常产生更多的关联加工,从而更容易诱发错误记忆(Howe & Derbish, 2013)。而本实验所用材料既非 DRM 词表也非类别词表,所以生存条件下的错误回忆率并不一定比其他条件高(Kroneisen, Rummel, & Erdfelder, 2014)。总结以上观点, 本研究正确回忆率和错误回忆率的结果均与主题优势的预期相悖, 说明主题优势不是生存加工优势效应产生的原因。
还有研究者认为生存加工优势效应可理解为一致性效应。如果一致性效应解释成立, 那么生存条件与材料的关联性要高于非生存条件。通过比较两条件的评定值, 发现生存条件和非生存条件的评定值没有差异, 说明在该实验条件下, 生存加工优势效应不能等同于一致性效应。
已知个体与学习材料接触时间长短也会影响最终的记忆成绩。因此, 本实验对词语与两情境的评定时间进行了分析, 结果显示, 两组的评定时间差异不显著, 所以评定时间不足以解释本实验出现的生存加工优势效应。
为了避免因主题不同而带来的潜在无关变量,本实验将情境的主题统一为“狩猎”。但有研究者(Silverman & Eals, 1992)认为在狩猎—采集的社会环境中, 男性与女性的劳动分工尤为典型, 男性负责狩猎而女性负责采集。男性也通常在与狩猎有关的技能上比女性表现得好(Voyer, Postma, Brake, &Imperato-McGinley, 2007)。所以, 本实验中的狩猎主题可能对女性被试来说是不太适合的。但以正确回忆率为因变量, 性别与情境为自变量进行方差分析的结果却表明, 性别对回忆成绩没有影响,F(1,296) = 1.31,p> 0.05= 0.004, 且交互作用不显著,F(1, 296) = 0.41,p> 0.05,η2p= 0.001, 表明男女被试在两种情境下的回忆成绩趋势一致。说明在本实验条件下以狩猎为主题的情境材料对于不同性别的被试来说是公平的, 这一论证与Nairne等(2009)得到的结果一致, 即在生存加工范式下, 狩猎与采集情境中的记忆成绩并没有表现出性别差异。
本实验发现, 同狩猎比赛情境相比, 远古狩猎条件下产生了更好的记忆效果, 也就是说人们对与生存有关的信息记得更好, 对材料进行生存加工产生了很好的记忆效果, 这是记忆适应性的表现。
3 实验2 繁衍加工的优势效应
3.1 研究目的
选取生存和自我联想情境同繁衍情境进行比较, 考查繁衍情境的优势效应。参照生存加工范式,对学习材料进行繁衍加工是否也会出现繁衍优势效应?本实验将自我联想和生存情境作为基线水平, 与择偶、怀孕、抚育情境比较来考查繁衍加工是否也具有优势效应。繁衍加工如果具有优势效应, 那么它与生存加工的优势效应之间又是否存在差异?
3.2 研究假设
如果自然选择理论的解释成立, 那么择偶、怀孕和抚育情境下被试的回忆成绩要高于自我联想情境下被试的回忆成绩, 表现出繁衍加工优势效应;且繁衍加工优势效应与生存加工优势效应不存在明显差异。
3.3 方法
3.3.1 被试
随机选取大学生 150名, 男生 70名, 女生 80名, 平均年龄20.72岁(SD = 1.74)。所有被试裸眼或矫正视力正常。每个被试在实验结束后均获得一份小礼品。
3.3.2 材料
从Battig and Montague norms (2004)分类词表中挑选出100个词并将这些词翻译成汉语, 随机选取未参加正式实验的 16名大学生分别对这些词语同生存情境、择偶情境、怀孕情境、抚育情境和自我联想情境进行关联度评定。评定采取5点量表法,其中1表示完全不相关, 5表示完全相关。对评定结果进行整理后, 选出37个词, 这些词与5个情境的关联度大致相等, 差值小于1.0。其中5个用于练习,32个词用于正式实验。
3.3.3 实验设计
采用单因素实验设计。自变量为加工条件。自变量的5个水平分别是生存加工、择偶加工、怀孕加工、抚育加工和自我联想。被试被随机分配到5种情境, 每种情境30人。因变量为正确回忆率、错误回忆率、词语的评定值和评定时间。
3.3.4 实验程序
实验程序采用 E-prime编制, 由电脑呈现, 安排被试坐在距计算机屏幕约50 cm处。正式实验前有练习, 使被试了解实验程序。整个实验持续约20 min。正式实验共包括以下三部分:
(1)学习阶段, 即对词语和情境进行关联度评定阶段。5种情境的指导语如下:
生存加工:请想象自己困在陌生的大草原上,没有任何生存物资。在随后的6个月, 你需要找到稳定的食物和水源, 并且保护自己免受肉食动物的攻击。下面将呈现一些词语, 你的任务是对这些词语所指代的事物与你所处的生存情境进行相关性评定。
择偶加工:请想象你即将参加一项异性联谊活动, 你的任务是采取相应的措施保证在此活动中找到终身伴侣。下面将呈现一些词语, 你的任务是对这些词语所指代的事物与你在联谊活动中找到终身伴侣进行相关性评定。
怀孕加工:请想象几年之后, 你成了家, 并准备有个小宝宝。你的任务是采取相应的措施来保证自己(妻子)顺利怀孕。下面将呈现一些词语, 你的任务是对这些词语所指代的事物与你成功孕育小宝宝进行相关性评定。
抚育加工:请想象几年之后你结婚生子, 在接下来的若干年, 你的任务是抚养和照顾自己的孩子,保证他(她)能健康的成长。下面将呈现一些词语,你的任务是对这些词语所指代的事物与你成功地抚养和照顾孩子进行相关性评定。
自我联想:请回想一下自己的人生经历, 下面将会呈现一些词语, 你的任务是对这些词语所指代的事物与帮助你回想起重要经历的容易程度进行评定。
每个词语呈现5 s, 评定等级为1~5, 1表示完全不相关, 5表示完全相关, 待评定等级出现在每个词语的下方, 被试需要在5 s内按键盘上的数字键做反应。如果在5 s内未作出反应, 则会呈现下一个词语。
(2)干扰阶段。让被试完成 24道计算题, 每道题呈现5 s, 干扰阶段持续2 min。
(3)测验阶段。在没有提前告知有测验任务的情况下, 要求被试尽可能多的回忆出刚才评定过的 32个词语, 并将词语写到答题纸上, 该阶段持续10 min。
3.4 结果
被试都能顺利完成实验任务, 对实验中99%以上的目标词都进行了评估, 并且未对目标词做出评估(按键反应出现在 5 s之后)的反应数目没有组间差异, 因此在数据分析中不对未作反应的数据进行删除。
(1)各加工情境下记忆效果的比较
为考查不同情境加工的记忆效果, 对被试在 5种情境中的正确回忆率进行统计, 结果如表4所示。
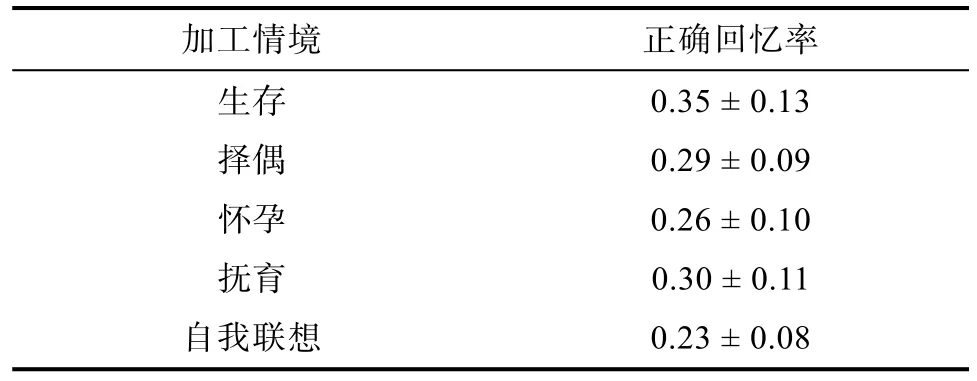
表4 5种情境下被试的正确回忆率(M ± SD)
Levene方差检验结果显示, 各实验处理内总方差为齐性,F(4, 145) = 1.85,p> 0.05。对5种情境下被试的正确回忆率进行单因素方差分析, 结果表明, 情境主效应显著,F(4, 149) = 5.09,p< 0.01,ηp2=0.123。进一步多重比较结果表明:生存情境下被试的回忆成绩高于自我联想情境(p< 0.01), 出现生存加工优势效应; 择偶情境下被试的回忆成绩高于自我联想情境(p= 0.051), 达到边缘显著, 抚育情境下被试的回忆成绩高于自我联想情境(p< 0.05), 怀孕情境下被试的回忆成绩与自我联想情境相等(p= 0.29),说明在择偶和抚育情境上出现了繁衍加工优势。
(2)各加工情境下错误回忆率的比较
对词语进行情境加工能提高个体的记忆成绩,但可能会引起更多的错误回忆, 为考查不同加工情境出现的错误回忆, 对5种情境下的错误回忆成绩进行统计, 结果如表5所示。
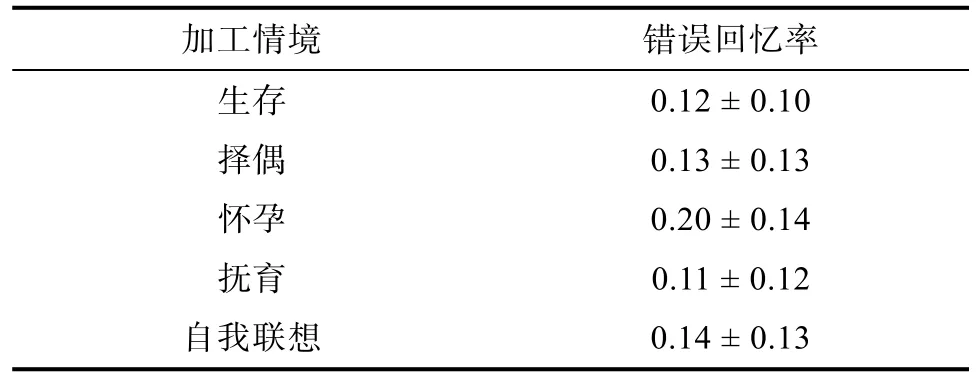
表5 5种情境下被试的错误回忆率(M ± SD)
Levene方差检验结果显示, 各实验处理内总方差为齐性,F(4, 145) = 0.91,p> 0.05。对不同情境下的错误回忆成绩进行单因素方差分析, 结果表明:情境主效应显著,F(4, 149) = 2.46,p= 0.048 <0.05,ηp2= 0.064, 说明不同情境下被试的错误回忆率差异显著。进一步的多重比较显示, 生存、择偶和抚育情境下被试的错误回忆率与自我联想情境相等, 怀孕情境下被试的错误回忆率与自我联想情境达到边缘显著,p= 0.071。说明与自我联想情境相比, 生存、择偶和抚育情境在回忆时没有产生更多的错误干扰, 而怀孕情境产生了更多的错误干扰。
(3)各加工情境下词语评定值和评定时间的比较
词语与情境的关联度和评定时间会对个体的记忆成绩产生影响, 因此, 对 5种情境下的词语和情境的关联度评定值和评定时间进行统计, 结果如表6所示。
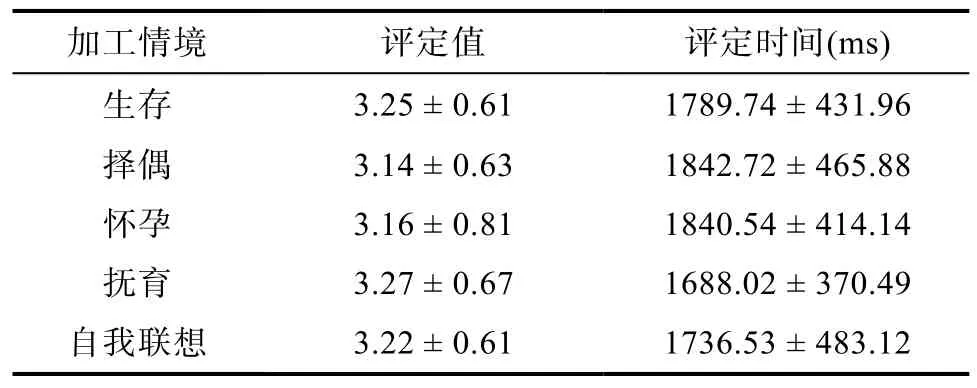
表6 5种情境下的评定值和评定时间(M ± SD)
Levene方差检验结果显示, 在评定时间与评定值上, 各实验处理内总方差为齐性,F(4, 145) =0.89;F(4, 145) = 1.89,p> 0.05。对不同情境下的评定时间和评定值进行单因素方差分析, 结果表明,评定时间主效应不显著,F(4, 149) = 0.72,p> 0.05,= 0.019, 说明5种情境下被试的评定时间没有差异; 评定值主效应也不显著,F(4, 149) = 0.22,p> 0.05,= 0.006, 说明5种情境下被试的评定值相等。
3.5 讨论
实验 2旨在通过考查繁衍的三种情境——择偶、怀孕和抚育是否会出现记忆优势, 探讨生存加工优势效应的自然选择解释是否成立。
实验结果显示:择偶和抚育情境下的正确回忆成绩高于自我联想情境, 错误回忆成绩与自我联想情境相等, 表现出繁衍加工优势, 符合预期假设,支持自然选择理论。
但是实验中没有出现怀孕情境的优势效应, 可能是因为情境对适应性意义的贡献大小不同造成的。现代进化理论认为, 所有的适应性问题都必须和繁衍活动有关, 并能够直接或间接地增加有机体的繁殖成功率。它认为适应性问题主要包括:(1)生存和成长的问题, 主要是让有机体存活到能够繁衍的年龄; (2)择偶问题, 主要指选择、吸引并留住配偶, 进行繁衍所必需的行为; (3)抚育问题, 主要指帮助后代生存和生长, 让后代存活到能够繁衍的年龄; (4)帮助遗传亲属的问题, 主要指帮助拥有个体基因的遗传亲属获得繁衍成功(Buss, 2007)。在上述适应中, 择偶和抚育都是为保证后代的质量, 而怀孕似乎是择偶之后顺其自然的事情。在实际生活中, 也确实能够看到人们在择偶和抚育行为中会投入更多的精力并做出更加慎重的选择, 而仅有部分个体因为婚龄过大或身体状况才会更多的做出保证怀孕的工作, 这可能是怀孕情境没有产生记忆优势的原因之一。另一个可能的原因是实验所用被试均为大学生, 几乎没有怀孕的经历, 而择偶观正逐步形成, 抚育行为也可通过父母长辈的耳濡目染而有所认识, 唯独怀孕情境是相对陌生的。所以被试在进行繁衍加工的时候, 由于没有过怀孕的经历,对怀孕的加工就不大可能有身临其境的感觉。这样就可能导致本来怀孕加工优势是存在的, 但却因为被试不知道何为怀孕, 出现了优势削弱。未来可通过使用有过怀孕经历的个体作为被试来验证上述可能, 如果在有怀孕经历的被试身上表现出了怀孕加工的记忆优势, 那么则说明对情境的理解与投入程度是造成本实验没有得到怀孕记忆优势的原因。如果在这些被试身上仍没有表现出记忆优势, 则说明怀孕的适应性意义相对较小。
同实验1类似, 实验2对词语的评定值和评定时间进行了分析, 结果发现各情境间没有差异, 进一步确定自然选择理论的解释在该实验条件下是成立的。
实验2并未将性别作为一个自变量进行考察。显然, 在人类的繁衍行为中, 男性和女性各自的角色和职责可能有所不同, 所以对于某种繁衍情境来说, 男性或者女性可能表现得更好。为了避免被试的性别差异而造成不同加工条件下的表现差异, 研究者在分配被试的时候已注意将各加工条件下的被试性别比例进行匹配。所以即便是男女在繁衍加工上表现不同, 也不会影响本实验的考查重点, 即人类在繁衍与生存加工上的记忆表现是否好于非适应性加工, 也就是本实验条件中的自我联想加工。
实际上, 在本实验中, 虽然在总的回忆成绩上女性要高于男性(p < 0.05), 但是就各个繁衍加工条件而言(择偶加工、怀孕加工、抚育加工), 男性与女性被试在正确回忆率上的差异不具有统计学上的意义(p > 0.05)。上述统计检验的数据, 是否表明在相关研究领域中不存在性别差异, 尚需进一步证实。如果确实不存在差异, 则说明繁衍加工优势效应与性别角色无关; 如果确实存在差异, 则说明,加工者个人在生活中扮演的角色可能对加工效果具有一定的影响。
自然选择理论认为生存和繁衍密切相关, 生存是繁衍的必要条件。但是对于生存和繁衍谁更重要的问题没有给出确定的答案, 有时保证生存是为了使自己具有繁衍的可能性, 但是有时为了能够繁衍,却使自己置身于危险之中。本实验同时采用了生存和繁衍情境, 结果显示, 生存情境下被试的回忆成绩高于择偶(p < 0.05)和怀孕(p < 0.05), 同抚育情境的回忆成绩达到边缘显著(p = 0.079), 参照生存加工范式, 可以说在本实验条件下, 生存比繁衍重要。
4 总讨论
生存加工优势效应是从记忆的功能性角度出发得出的, 自2007年Nairne等人发现生存加工优势效应以来, 研究者们对其展开大量研究, 证实了生存加工优势效应是普遍存在的。然而研究者们多通过对主题加工、一致性效应等解释的排除, 而间接地证明生存加工优势效应是自然选择的结果, 直接针对自然选择理论进行的研究则很少。自然选择理论认为:祖先在进化过程中因自然选择压力而形成的心理机制具有继承性, 记忆也是心理机制的一个方面; 人们存在很多由祖先遗传下来的适应器,这些适应器用来解决特定的适应性问题, 包括生存、繁衍等。根据自然选择理论的解释, 排除经验之后的个体在生存加工优势效应上的表现应不存在差异, 并且对繁衍进行加工时也应表现出繁衍的记忆优势(刘希平, 刘玉霞, 唐卫海, 2013)。本研究分别从自然选择的继承性和适应性问题的角度, 以记忆成绩为指标, 对生存加工优势效应的自然选择理论解释展开直接的证明。
实验1考查了不同年龄个体间生存加工优势效应的变化。我们推测生存加工优势效应应该会出现在人的生命全程中。已有研究多采用大学生为被试的进行研究, 有两篇分别以5岁、8岁、10岁年龄组(Aslan & Bäuml, 2012)和以8、11岁儿童(Otgaar &Smeets, 2010)为被试, 发现了生存加工优势效应,但是同时选取多个年龄段被试的研究比较少。为了考查低年龄段个体生存加工优势效应的具体表现,实验选取了5个年龄段的被试, 分别是8岁、10岁、12岁、21岁和64岁。结果显示, 5个年龄段都出现了生存加工优势效应, 并且各年龄段的生存优势效应量相等。然而, 关于老年人是否与其他年龄段个体表现出同样的生存加工优势效应, Stillman,Coane, Profaci, Howard和Howard (2014)的研究结果与本研究结果并不一致。Stillman等(2014)进行了三个实验, 既包括被试间设计也包括被试内设计,他们的研究中所使用的情境材料和词语与 Nairne等(2007)实验2所用材料相同。结果都表明老年被试没有产生生存加工优势效应, 即便是在生存条件下的评估值显著高于控制条件的情况下, 也就是可能由一致性效应而得到记忆优势的情况下, 也未见生存加工优势效应的产生。Stillman等提出这种表现可能是由于“老年人把他们已经减少的认知资源的优先权从获得新信息转移到更令人满意的、更具社会价值的目标上。”我们与Stillman 等(2014)的结果不一致, 我们猜测, 也许正是 Stillman等与我们的情境不同导致的。而果真如此, 则可能是老年人被试对Stillman等使用的材料并不敏感, 即使存在生存加工优势效应, 也并没有表现出来。在我们的研究中, 老年人的生存加工优势效应得到了充分的展示。为了考察此处分析是否有道理, 我们对青年人的生存加工优势大小与老年人的生存加工优势大小进行了比较, 发现青年人与老年人生存加工优势大小没有差异(p > 0.05)。Stillman等的研究中,虽然同样的材料在年青被试上获得了生存加工优势效应, 但存在一种可能是他们使用的材料并不是对所有年龄段被试都是同等适用的。从他们的研究中老年人没有表现出生存加工优势效应看, 可能搬家情境对老人而言加工效果比较强, 弱化了生存加工带来的效应。相比较而言, 我们的研究对生存加工组和控制组其情境控制更加公平。我们的研究应该是对Stillman研究的推进。如果这种分析不够到位, 则跨情境控制组的生存优势效应则应该成为今后研究继续探索的选题。另有Nouchi (2012)的研究使用了Nairne等(2007)所用生存情景, 对照组进行自我联想任务, 被试同样是老年人, 结果发现老年被试表现出生存加工优势效应, 但其生存优势量小于青年被试。Nouchi的研究中老年人的优势量小于青年人的原因可能在于其使用了被试内设计。以往仅对青年被试的研究发现被试内设计相对于被试间设计获得的生存优势量更大(Nairne et al., 2007),所以在Nouchi的研究中, 青年被试的优势增大; 老年人虽然也用的是被试内设计, 但是老年人可能更容易受任务转换的影响而更少地受益于被试内设计, 所以在优势量上表现出了年龄差异。虽然青年和老年被试在反应时间的数据上没有显著的差异,但是仍不能排除老年被试更易受任务转换影响的可能。另一种可能的原因在于对照条件下加工方式的不同。本研究中对照条件与实验条件同为情境加工, 两种情境加工相对照, 受老化影响的因素对两种加工条件的作用是相同的。而 Nouchi的研究中的对照条件为自我参照加工, 不同于实验条件下的情境加工, 那么有可能是两种加工方式受老化影响的不同, 造成了这种优势量上的年龄差异。
实验2考查了繁衍加工的记忆优势。实验采用生存、择偶、怀孕、抚育和自我联想5种情境探讨了繁衍加工的记忆优势, 比较五种情境的记忆成绩发现:生存情境下的正确回忆率高于自我联想情境,出现生存加工优势; 择偶和抚育情境下的正确回忆率均高于自我联想情境, 出现繁衍加工优势; 而怀孕情境下的正确回忆率同自我联想情境没有差异,这可能与怀孕情境有较少的适应性或大学生被试缺少怀孕经历有关。
研究结果符合预期假设, 说明在本实验条件下自然选择理论的解释是成立的。这一结果为自然选择解释提供了实验证据, 也因此奠定了更稳固的基础, 并且为生存加工优势效应作用机制的研究提供了具体的研究框架, 未来的研究可从这个框架展开,将节省大量时间。
综上所述, 人们的记忆系统确实具有功能性,对涉及生存和繁衍的信息记得更好, 自然选择理论可以解释生存加工优势效应。
5 结论
在本研究条件下, 得到以下结果:(1)不同年龄组个体均表现出生存加工优势效应, 但生存优势效应量是否存在年龄差异还需进一步验证; (2)择偶和抚育情境表现出繁衍加工优势。这一结果支持了生存加工优势效应的自然选择理论。
Aslan, A., & Bäuml, K. H. T. (2012). Adaptive memory:Young children show enhanced retention of fitness-related information. Cognition, 122, 118-122.
Burns, D. J., Hart, J., Griffith, S. E., & Burns, A. D. (2013).Adaptive memory: The survival scenario enhances itemspecific processing relative to a moving scenario. Memory,21, 695-706.
Buss, D. M. (Ed). (2007). Evolutionary psychology: The new science of the mind. Boston: Allyn and Bacon.
Butler, A. C., Kang, S. H., & Roediger III, H. L. (2009).Congruity effects between materials and processing tasks in the survival processing paradigm. Journal of Experimental Psychology: Learning, Memory, and Cognition, 35, 1477-1486.
Faul, F., Erdfelder, E., Lang, A.-G., & Buchner, A. (2007).G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior Research Methods, 39, 175-191.
Howe, M. L., & Derbish, M. H. (2010). On the susceptibility of adaptive memory to false memory illusions. Cognition,115, 252-267.
Howe, M. L., & Derbish, M. H. (2013). Adaptive memory:Survival processing, ancestral relevance, and the role of elaboration. In What is adaptive about adaptive memory.New York: Oxford University Press.
Kang, S. H. K., McDermott, K. B., & Cohen, S. M. (2008).The mnemonic advantage of fitness-relevant information.Memory & Cognition, 36, 1151-1156.
Kroneisen, M., Rummel, J., & Erdfelder, E. (2014). Working memory load eliminates the survival processing effect.Memory, 22, 92-102.
Liu, X. P., Liu, Y. X., & Tang, W. H. (2013). The interpretation of survival processing advantage effect. Journal of Psychological Science, 36, 240-244.
[刘希平, 刘玉霞, 唐卫海. (2013). 生存优势效应的理论解释. 心理科学, 36, 240-244.]
Mao, W. B., Yu, R., & Li, C. (2013). Effects of relevance and congruity on survival advantage of memory. Acta Psychologica Sinica, 45, 253-262.
[毛伟宾, 于睿, 李春. (2013). 一致性、相关性对记忆生存优势效应的影响. 心理学报, 45, 253-262.]
Nairne, J. S., & Pandeirada, J. N. (2010). Adaptive memory:Ancestral priorities and the mnemonic value of survival processing. Cognitive Psychology, 61, 1-22.
Nairne, J. S., & Pandeirada, J. N. (2011). Congruity effects in the survival processing paradigm. Journal of Experimental Psychology: Learning, Memory, and Cognition, 37, 539-549.
Nairne, J. S., Pandeirada, J. N., Gregory, K. J., & Van Arsdall,J. E. (2009). Adaptive memory fitness relevance and the hunter-gatherer mind. Psychological Science, 20, 740-746.
Nairne, J. S., Pandeirada, J. N., & Thompson, S. R. (2008).Adaptive memory the comparative value of survival processing. Psychological Science, 19, 176-180.
Nairne, J. S., Thompson, S. R., & Pandeirada, J. N. (2007).Adaptive memory: Survival processing enhances retention.Journal of Experimental Psychology: Learning, Memory,and Cognition, 33, 263-273.
Nouchi, R. U. I. (2012). The effect of aging on the memory enhancement of the survival judgment task. Japanese Psychological Research, 54, 210-217.
Öhman, A., & Mineka, S. (2001). Fears, phobias, and preparedness: toward an evolved module of fear and fear learning. Psychological Review, 108, 483-522.
Olds, J. M., Lanska, M., & Westerman, D. L. (2014). The role of perceived threat in the survival processing memory advantage. Memory, 22, 26-35.
Otgaar, H., Smeets, T., Merckelbach, H., Jelicic, M., Verschuere,B., Galliot, A. M., & Van Riel, L. (2011). Adaptive memory:Stereotype activation is not enough. Memory & Cognition,39, 1033-1041.
Otgaar, H., & Smeets, T. (2010). Adaptive memory: Survival processing increases both true and false memory in adults and children. Journal of Experimental Psychology: Learning,Memory, and Cognition, 36, 1010-1016.
Otgaar, H., Smeets, T., & Van Bergen, S. (2010). Picturing survival memories: Enhanced memory after fitness-relevant processing occurs for verbal and visual stimuli. Memory &Cognition, 38, 23-28.
Röer, J. P., Bell, R., & Buchner, A. (2013). Is the survivalprocessing memory advantage due to richness of encoding?Journal of Experimental Psychology: Learning, Memory,and Cognition, 39, 1294-1302.
Silverman, I., & Eals, M. (1992). Sex differences in spatial abilities: Evolutionary theory and data. In J. H. Barkow, L.Cosmides, & J. Tooby (Eds.), The adapted mind: Evolutionary psychology and the generation of culture (pp. 533-549).Oxford: Oxford University Press.
Stillman, C. M., Coane, J. H., Profaci, C. P., Howard, J., &Howard, D. V. (2014). The effects of healthy aging on the mnemonic benefit of survival processing. Memory &Cognition,42, 175-185.
Tooby, J., & Cosmides, L. (1995). The psychological foundations of culture. In J. H. Barkow, L. Cosmides, & J. Tooby (Eds.),The adapted mind: Evolutionary psychology and the generation of culture (pp. 19-136). Oxford: Oxford University Press.
Voyer, D., Postma, A., Brake, B., & Imperato-McGinley, J.(2007). Gender differences in object location memory: A meta-analysis. Psychonomic Bulletin & Review, 14, 23-38.
Weinstein, Y., Bugg, J. M., & Roediger, H. L. (2008). Can the survival recall advantage be explained by basic memory processes? Memory & Cognition, 36, 913-919.
Yu, R., Mao, W. B., & Jia, Z. (2011). Survival processing: A special and powerful memory encoding procedure.Advances in Psychological Science, 19, 825-831.
[于睿, 毛伟宾, 贾喆. (2011). 生存加工: 一种独特而强大的记忆编码程序. 心理科学进展, 19, 825-831.]
