6种草本植物对干旱胁迫和CO2 浓度升高交互作用的生长响应
2015-01-19高凯敏刘锦春梁千慧AndriesTemmeJohannesCornelissen
高凯敏, 刘锦春, 梁千慧, Andries A. Temme, Johannes H.C. Cornelissen
1 三峡库区生态环境教育部重点试验室, 重庆市三峡库区植物生态与资源重点试验室, 西南大学 生命科学学院, 重庆 400715 2 Department of Ecological Science, VU University Amsterdam, De Boelelaan 1085,1081 HV Amsterdam, the Netherlands
6种草本植物对干旱胁迫和CO2浓度升高交互作用的生长响应
高凯敏1, 刘锦春1,*, 梁千慧1, Andries A. Temme2, Johannes H.C. Cornelissen2
1 三峡库区生态环境教育部重点试验室, 重庆市三峡库区植物生态与资源重点试验室, 西南大学 生命科学学院, 重庆 400715 2 Department of Ecological Science, VU University Amsterdam, De Boelelaan 1085,1081 HV Amsterdam, the Netherlands
以CO2浓度升高为主要标志的全球气候变化及由其引起的极端气候变化对陆地生态系统产生了重要的影响。利用步入式CO2生长室模拟研究了CO2浓度变化(400和700μL/L)和干旱胁迫(水分充足CK:100%FC(田间持水量);中度干旱MS:40%FC;重度干旱SS:20%FC)的交互作用对草本植物网果酸模(Rumexchalepensis)、野豌豆(Viciasepium)、泥胡菜(Hemmisteptalyrata)、风轮菜(Clinopodiumchinense)、藜(Chenopodiumalbum)和玉米石(Sedumalbum)生长特性的影响。结果表明:CO2浓度升高总体上刺激了网果酸模、野豌豆、泥胡菜、风轮菜和藜这5种C3植物在任何水分条件下的生长,也刺激了玉米石在水分条件较好下的生长;干旱胁迫总体上抑制了所有6种植物的生长,但中度干旱胁迫有刺激CAM植物玉米石生长的趋势。CO2浓度升高能否缓解干旱的负面影响具有明显的种间差异:CO2浓度升高减缓了干旱胁迫对泥胡菜和风轮菜的负面影响,这种缓解作用在网果酸模和野豌豆中显著降低,对藜没有明显的促进作用,对干旱下的玉米石的生长却起到了抑制作用。CO2浓度升高总体上增加了根质量分数和干物质含量;干旱胁迫明显提高了6种草本植物的根生物量的分配比例,降低了干物质含量;但CO2浓度升高和干旱胁迫的交互作用可导致不同的物种产生不同的响应,说明植物能够通过调节生物量分配和植株本身的水分含量保持能力来适应CO2浓度和干旱胁迫的交互影响,这种调节能力取决于植物在碳的吸收和水分散失之间的平衡“trade-off”。 研究结果有助于增进草本植物对未来气候变化的适应性理解,为评估和预测全球气候和水文变化对植物的生理生态影响提供理论依据。
CO2浓度升高; 干旱胁迫; 生物量分配; 草本植物; 田间持水量 (FC)
工业革命以来,由于人类大量消耗石油化工燃料和对土地的不合理开发利用,大气中CO2浓度以每年0.4%的速度持续升高,预计到21世纪末,CO2浓度将从目前的约400 μmol/mol增加到700 μmol/mol[1-2]。CO2浓度升高导致的温室效应也加速了全球水循环,引起全球降水的格局变化[3],导致一些干旱地区更加干旱,一些湿润地区的极端干旱事件也频繁发生[4-5]。
植物光合作用时,CO2的获取和水分的散失同时发生。因此,大气CO2浓度和土壤水分总是相互作用,共同影响植物的生长、发育和繁殖,从而影响植物生产力、决定群落结构、并最终影响全球植被的分布及陆地生态系统功能[6]。目前的研究普遍认为,CO2浓度升高对植物的生长和生产力的促进作用(即“施肥效应”)受到土壤水分条件的影响[7-8]。Poorter 和 Pérez-Sobás[9]的综述表明,14种草本植物中有13种植物在干旱条件下受到CO2浓度升高的促进作用大于对照(水分充足时)。Atwell 等[10]发现只有在干旱条件下CO2浓度升高才刺激了细叶桉(Eucalyptustereticornis) 的生长,在水分充足条件下反而没有促进作用。同样,在C4植物的研究中,研究者也发现CO2浓度倍增对提高C4植物的净光合速率无明显的作用[11],但在频繁和严酷的干旱条件下,CO2浓度升高可以缓解干旱对C4植物的不利影响[12],并且在干旱胁迫越严重时表现得更加明显。然而,也有研究认为:CO2浓度升高对植物的促进作用在水分条件充足时远大于干旱条件下。如Xu等[13]发现东北半干旱草原的典型灌木中间锦鸡儿(Caraganaintermedia)在对照、轻度干旱和重度干旱下CO2浓度增加对其生物量的促进作用分别是81%,63%和45%,可见,干旱越严重,CO2浓度的施肥效应越低。Erice 等[14]也认为在水分条件较好时,CO2浓度升高增加了植物的生物量,但在干旱条件下CO2浓度升高对植物的生物量没有影响。因此,CO2浓度升高是否可以缓解干旱对植物的影响,或者说植物对CO2浓度升高的响应是否受到土壤水分含量的限制,可能与干旱的程度及物种有关[15-16]。
通常情况下,CO2浓度升高可以通过降低气孔导度来提高水分利用效率和土壤水分含量,从而刺激干旱条件下植物的生长和叶面积的增加[17-18]。然而,由于CO2浓度升高的刺激作用,促进了植物的生长和生物量的累积[19],导致植株增大,水分吸收总量增多,土壤水分消耗增大,从而又有可能降低干旱条件下植物的水分利用效率[20]。因此,在将来可能的CO2升高增加和干旱背景下,植物究竟如何解决CO2吸收和水分散失之间的平衡难题?CO2浓度升高是否能够减缓干旱对植物的影响?不同的物种之间是否对CO2浓度升高和干旱具有一致的响应?这些问题的回答无疑对预测全球气候变化对陆地生态系统的影响具有重要的意义。由于一年生草本植物生长速率较快,对CO2浓度等环境因子变化十分敏感,能够快速而客观表达植物与外部环境相互关系[21],因此本研究以6种常见的草本植物为试验对象,探讨植物对CO2浓度升高和干旱胁迫交互作用的响应,以验证以下假设:
(1) CO2浓度升高可能缓解或者加剧干旱对植物的影响,或者没有影响,即植物对CO2浓度升高和干旱胁迫的交互作用存在明显的种间差异。
(2) 不同植物对CO2浓度升高和干旱胁迫交互作用存在着不同的生物量分配和保水能力差异,这决定了植物在CO2吸收和水分散失之间存在平衡能力的差异。
1 材料与方法
试验在荷兰乌特勒支大学特制的步入式CO2浓度控制生长室内完成。以6种常见的草种网果酸模(Rumexchalepensis)、野豌豆(Viciasepium)、泥胡菜(Hemmisteptalyrata)、风轮菜(Clinopodiumchinense)、藜(Chenopodiumalbum)和玉米石(Sedumalbum)为材料,于2012年3月15日在当前CO2浓度(400μL/L)生长室下进行发芽,出苗后每个物种选取长势基本一致的62株幼苗分成两组移栽于0.4 L内盛等量沙子的花盆中,置于当前CO2浓度(400μL/L)和可预测的将来高CO2浓度(700μL/L)两个不同浓度CO2生长室,同时设置光照条件为400 μmol m-2s-1,温度为25℃ /18℃,相对湿度为70%。同时对每个CO2生长室的6个试验物种进行2种干旱处理:中度干旱胁迫(MS,40%FC(田间持水量))和重度干旱胁迫(SS,20%FC)处理,以土壤水分充足(CK,100%FC)为水分设置的对照。每个物种6个处理,且每个处理8个重复。每2天对幼苗补充一次稀释2倍的Hoagland 营养液,待幼苗第1片真叶完全展开后,对每个物种中的8盆进行初始收获。同时对每个生长室内的每个物种的植株进行水分处理,并使每个处理仍然保持稀释2倍的Hoagland 营养液浓度。水分处理采用称重法控制[22],每周对每个处理收获2株用以校正幼苗生长对土壤水分测定造成的误差。试验处理时间为3周。3周后分成地上部分和地下部分进行收获,分别称量鲜重后装入牛皮纸袋,置入烘箱中105℃杀青15min,然后在80℃下烘干至恒重,再称量各部分干重,进一步计算总生物量:
根质量分数 (RMF)= (根生物量/总生物量)×100%
干物质含量(DMC)=(总干重/总鲜重)×100%
数据用SPSS 20.0进行统计,先以试验幼苗的初始生物量作为协变量进行协因素方差分析(Covariance)以检测不同物种的初始生物量对后期生长的影响(carry-over effects),结果显示无显著性差异,然后采用三因素方差分析(ANOVA)以分析不同CO2浓度和水分处理及其交互作用对6种草本植物的影响及种间差异。
2 结果分析
2.1 存活情况
处理过程中有少数植株可能因处理不当而出现异常情况,故每个处理最终有6—8株不等(表1),在试验结束时6个物种存活情况不同:野豌豆和玉米石全部存活,而其他物种均存在死亡现象,网果酸模在高CO2浓度下的重度干旱(SS)条件下全部死亡,泥胡菜和风轮菜在当前CO2浓度下的重度干旱(SS)条件下死亡率分别为83.3%和87.5% (表1)。

表1 6种草本植物在不同CO2浓度和水分处理3周后的存活情况
CK: Control;MS: 中度干旱Mild drought stress;SS: 重度干旱Severe drought stress;FC:田间持水量 Field capacity
2.2 生物量
除了玉米石, CO2浓度升高均使其他5种植物总生物量呈增加的趋势,干旱程度增加均降低这5种植物总生物量(图1)。在水分充足(CK)条件下,CO2浓度升高刺激了网果酸模和野豌豆生物量的积累,其升高幅度分别为39%和49%。然而,随着干旱程度增加,CO2浓度升高的刺激作用显著降低:在中度干旱(MS)条件下,CO2浓度升高对网果酸模生物量没有显著影响,而野豌豆生物量的增幅降低到21%;在重度干旱(SS)条件下,CO2浓度升高对野豌豆生物量也没有显著影响(图1)。对于泥胡菜和风轮菜,在水分充足(CK)条件下,CO2浓度升高刺激其生物量增加的幅度分别为21%和0%,但在中度干旱(MS)条件下,泥胡菜的生物量增加了391%,而风轮菜的生物量增加了85%,可见,泥胡菜和风轮菜的生物量在CO2浓度升高刺激下的提高幅度随着干旱程度增加而显著增大(图1)。对于藜,CO2浓度升高对其生物量并没有明显的促进或抑制作用(生物量在两种CO2浓度下没有显著差异)(图1)。对于玉米石,在水分充足(CK)条件下,CO2浓度升高对其生物量没有产生显著影响,然而,随着干旱程度增加,CO2浓度升高反而抑制了玉米石的生物量积累,在中度(MS)和重度(SS)干旱条件下其生物量下降幅度分别为75%和78%(图1)。三因素方差分析表明生物量在物种、水分处理、CO2浓度处理下均具有极显著差异,且在物种和水分处理间、物种和CO2浓度处理间具有显著交互作用,但在水分处理和CO2浓度处理间及物种、水分处理和CO2浓度处理三者间没有显著交互作用(表2)。
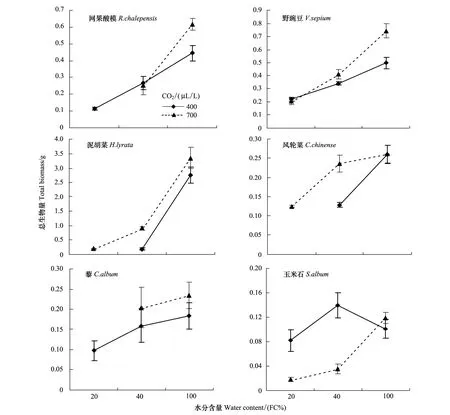
图1 不同CO2浓度和干旱胁迫下网果酸模、野豌豆、玉米石、泥胡菜、风轮菜和藜的总生物量(平均值±标准误)Fig.1 Total biomass of R.chalepensis, V.sepium, S.album, H.lyrata, C.chinense, C.album under two CO2 concentrations and three water regimes (M±SE)图中无数据则表示该物种在此条件下死亡; FC:田间持水量 Field capacity
2.3 根质量分数
除了玉米石,在两种CO2处理下,干旱程度增加总体上均使植物的根质量分数增加。在任何水分条件下,CO2浓度升高均没有刺激网果酸模的根质量分数显著增加(尽管在中度干旱(MS)条件下,CO2浓度升高刺激了根质量分数少量增加,但与当前CO2浓度处理无显著差异)。在水分充足(CK)条件下,CO2浓度升高使野豌豆的根质量分数增加11%,但随着干旱程度增加,CO2浓度升高反倒使根质量分数下降,在中度(MS)和重度(SS)干旱条件下,其下降幅度分别为4%和11%(图2)。在水分充足(CK)条件下,CO2浓度升高使泥胡菜的根质量分数升高53%,但在中度干旱(MS)条件下,根质量分数下降了13%;而风轮菜的根质量分数在任何水分条件下均没有受到CO2升高的显著影响(图2)。对于藜,在水分充足(CK)条件下,CO2浓度升高使其根质量分数增加11%,但在中度干旱(MS)条件下,CO2浓度升高使其根质量分数下降6%,但没有显著影响(图2)。对于玉米石,在水分充足(CK)条件下,CO2浓度升高对其根质量分数没有显著影响,但随着干旱程度的增加,CO2浓度升高促进了根质量分数的增加,在中度(MS)和重度(SS)干旱胁迫下,其根质量分数分别增加了96%和58%(图2)。三因素方差分析表明根质量分数在物种、水分处理、CO2浓度处理下均具有极显著差异,且在物种和水分处理间、物种和CO2浓度处理间及物种、水分处理和CO2浓度处理三者间均有显著交互作用,只有水分处理和CO2浓度处理间没有表现出显著的交互作用(表2)。
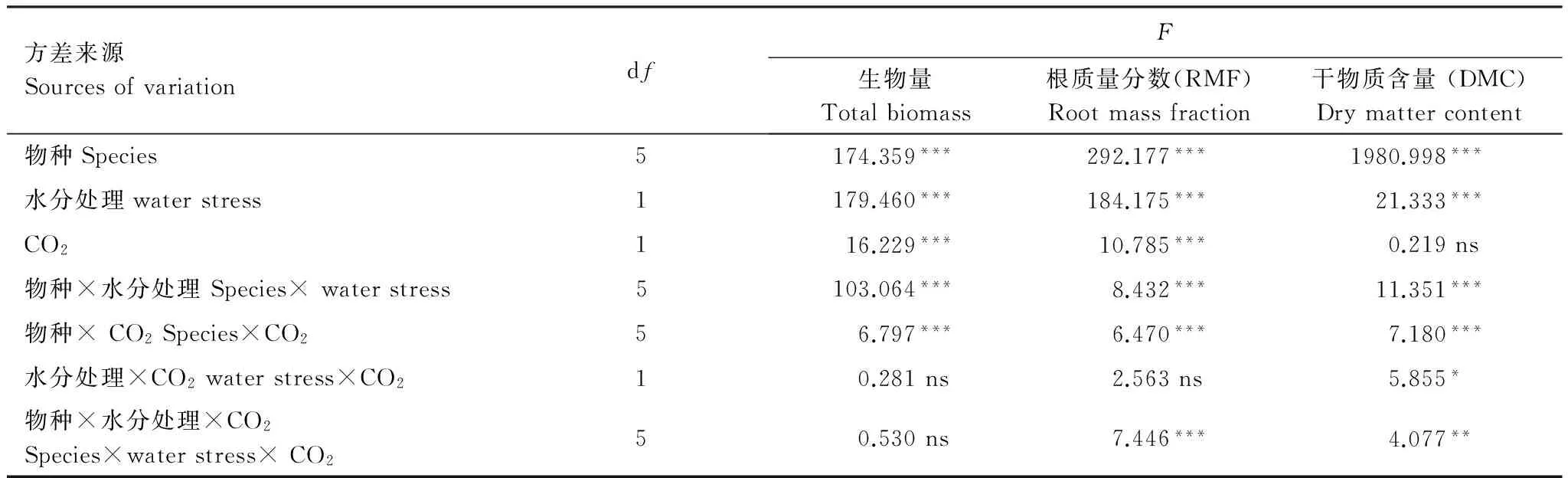
表2 6个草本物种对不同CO2浓度和干旱胁迫响应的三因素方差分析

图2 不同CO2浓度和干旱胁迫下网果酸模、野豌豆、玉米石、泥胡菜、风轮菜和藜的根质量分数(平均值±标准误)Fig.2 Root mass fraction (RMF) of R.chalepensis, V.sepium, S.album, H.lyrata, C.chinense, C.album under two CO2 concentrations and three water regimes (M±SE) (Plants that had died were excluded from these analyses)
2.4 干物质含量
CO2浓度升高增加了网果酸模和野豌豆在中度干旱(MS)条件下的干物质含量,而在水分充足(CK)和重度干旱(SS)条件下,没有显著影响(图3)。CO2浓度升高总体上降低了泥胡菜和风轮菜的干物质含量,但随着干旱程度的增加,泥胡菜干物质的降低幅度逐渐下降,从水分充足(CK)条件下的13%下降到中度干旱(MS)条件下的0.3%,而风轮菜干物质含量的降低幅度逐渐升高,从水分充足(CK)条件下的1%上升到中度干旱(MS)下的15%(图3)。对于藜,CO2浓度升高增加了其干物质含量,在水分充足(CK)条件下,CO2浓度升高使其干物质含量增加21%,而在中度干旱(MS)条件下,其增加幅度下降到6%(图3)。CO2浓度升高在水分充足(CK)条件下并没有增加玉米石的干物质含量,但在干旱胁迫下显著增加,其增加幅度随着干旱程度的增加而增大,从中度干旱(MS)下的68%升高到重度干旱(SS)下的98%(图3)。三因素方差分析表明干物质含量在CO2浓度处理下没有显著差异,而在其它处理、处理的两两间和处理的三者间均具有显著或极显著的交互作用(表2)。

图3 不同CO2浓度和干旱胁迫下网果酸模、野豌豆、玉米石、泥胡菜、风轮菜和藜的干物质含量(平均值±标准误)Fig.3 Dry matter content (DMC) of R.chalepensis, V.sepium, S.album, H.lyrata, C.chinense, C.album under two CO2 concentrations and three water regimes (M±SE) (Plants that had died were excluded from these analyses)
3 讨论
植物对其光合底物CO2浓度的响应,往往受到其它环境因子如土壤水分等的综合影响。一般情况下,一定范围内CO2浓度升高对植物具有重要的施肥作用,同时能够减缓干旱胁迫对植物的影响[23-24]。本研究发现,CO2浓度升高促进了网果酸模和野豌豆在水分充足条件下的生物量积累,而随着干旱程度的增加,其生物量积累显著减低,说明在水分充足条件下,CO2浓度升高确实对网果酸模和野豌豆具有重要的“施肥效应”,但发生土壤干旱胁迫时,CO2施肥效应受到了一定程度的抑制[25-26],同时也表明了CO2浓度升高没有减缓干旱胁迫对这两种草本植物的负面影响。通常情况下,CO2浓度升高导致气孔关闭,气孔导度下降,从而提高了水分利用效率,然而,由于光合底物CO2充足,光合速率增加,促进了植物的生长和生物量的累积[19],导致植株增大,水分吸收总量增多,土壤水分消耗增大,从而又降低了水分利用效率[20],最终导致CO2浓度升高对干旱的减缓作用被消弱。但对于泥胡菜和风轮菜,本研究发现在水分充足条件下,CO2升高对两者的生物量没有显著的影响,但随着干旱程度的增加,却有显著的施肥效应。这与王云霞等[27]发现高CO2浓度下玉米和高粱的产量增幅在干旱条件明显大于湿润条件的研究结果一致。这表明了CO2浓度升高导致植物叶片气孔导度降低, 降低了蒸腾速率, 使单位叶面积土壤水分耗损率降低,提高了植物水分利用率, 从而增加了植物避免干旱的能力[17-18]。然而,不管在哪种水分条件下,CO2升高对藜的生物量没有显著影响,而干旱胁迫导致其生物量显著降低,可能是CO2升高对藜的正面影响和干旱胁迫的负面影响相当,相互抵消了所致。在水分充足条件下,CO2浓度升高对玉米石生物量无显著影响,与泥胡菜和风轮菜结果一样,然而在干旱条件下,CO2浓度升高显著降低了玉米石的生物量。这可能是由于玉米石为CAM植物,具有特殊的光合机制所致:在当前CO2浓度下,为了减缓干旱的影响,气孔部分关闭,PEP羧化酶的活性已经饱和或者接近饱和, CO2浓度升高进一步引起气孔变小或关闭,CO2的吸收大量减少,CO2的同化速率降低[28], CO2浓度升高失去了其补偿作用,导致植物的生长在高CO2浓度下受限。
植物资源分配是指植物将同化的资源分配于不同功能器官的比例。 植物能在环境资源限制条件下将光合作用所固定的能量进行优化分配,从而提高自身在多变环境中的适合度[29],根质量分数反映的就是植物对根系的资源分配比例。在干旱胁迫环境中,植物通常通过增大根质量分数,向深层土壤延伸以利用深层土壤水来适应干旱环境,以维持生长[30]。本研究中的6种草本植物,不管在何种CO2浓度处理下,根质量分数均随着干旱程度的增加而增大, 说明这6种草本植物确实能够通过提高根系生物量分配而提高对干旱环境的适应。CO2浓度升高通常也促进植物根系生物量分配[31],因为CO2浓度升高增加了植物叶片的净同化率[32],这样就使得整个冠层C 获取增加,地下总非结构性碳水化合物迁移增加导致根系生物量增多[33]。本研究中发现,CO2浓度升高能够提高CK条件下的野豌豆、泥胡菜和藜根系生物量的分配比例,可能原因是这3种植物在水分较好和CO2浓度升高条件下,根、冠生长速度基本一致,地上、地下资源均不形成限制造成的[34]。然而,随着干旱程度的增加,CO2浓度升高对这3种植物根系生物量分配的刺激作用显著降低,在中度干旱条件下刺激作用消失,在重度干旱时反而有抑制作用,可能原因是在中度干旱时植物适度关闭了气孔,一定程度限制了叶片对碳的吸收,导致植株对水的需求降低,植物对根和冠的生物量分配恰好能够达到平衡;但干旱程度继续增加时,为了提高水分利用效率,气孔导度继续降低,理论上此时植株应该依照植物提高逆境竞争能力的方式来满足能量和物质的平衡,即相对减少光合产物向营养器官的投入,而投资更多的生物量分配于根系以吸收更多的水分,但干旱已严重抑制了植株的生长,且对根的抑制程度大于冠[35-36],故导致最后的根质量分数显著降低。对于玉米石,尽管在水分充足条件下,CO2升高对其生物量没有影响,但在干旱条件下,CO2升高却促进了其根质量分数的增加,可能原因是CAM植物玉米石在干旱下气孔关闭较多,以提高水分利用效率,但以消耗光合作用的合成为代价,光合作用降低,也导致了地下总非结构性碳水化合物迁移减少,从而导致根系生物量分配显著下降,这也进一步说明了CO2浓度升高反而加剧干旱胁迫对玉米石的生长影响。
干物质含量DMC是反映植物生态行为差异的指标,它可以表示为植物干物质重量和植物鲜重的比值,用于表示植物在自然状态下的含水率状况,可以反映植物获取资源的能力[37]。本研究中6种草本植物的干物质含量不管在何种CO2浓度下,总体上均随着干旱程度的增加而升高,这是由于植株在干旱胁迫中逐渐失水所致,但玉米石在当前CO2浓度下,即使土壤水分降低,干物质含量也基本上保持不变,原因是玉米石为CAM植物,肉质、多汁,具有很好的贮水系统,即使是在重度干旱下仍能保持较高的水分含量,而在高CO2浓度下,由于高CO2浓度并不能缓解干旱对其的负面影响,在干旱程度加剧情况下,植株失水过多,干物质含量也随之升高。不管土壤水分含量如何变化,CO2浓度升高均增加了网果酸模和野豌豆的干物质含量,是因为与CO2浓度升高刺激了植物的光合物质的积累[38]。对于藜和玉米石,CO2浓度升高虽然也增加了两者的干物质含量,却与光合产物的积累无关,而与其保水能力较强有关。由于CO2对光合的刺激作用与植物对水分的保持作用不同,最终也导致了不同的植物干物质含量在CO2浓度处理和水分处理交互作用下的差异。
4 结论
CO2浓度升高总体上刺激网果酸模、野豌豆、泥胡菜、风轮菜和藜这5种C3植物在任何水分条件下的生长,也刺激了玉米石在水分条件较好下的生长;干旱胁迫总体上抑制了所有6种植物的生长,但中度干旱胁迫有刺激CAM植物玉米石生长的趋势。CO2浓度升高减缓了干旱胁迫对泥胡菜和风轮菜的负面影响,但对于网果酸模和野豌豆,CO2施肥效应受到了干旱胁迫的抑制,也导致CO2浓度升高对干旱胁迫的减缓作用显著降低;而对于藜,干旱对其生长影响并不受CO2浓度变化的影响;但是CO2浓度升高却对干旱下的CAM植物玉米石的生长起到抑制作用。可见,CO2浓度升高能否缓解干旱的负面影响具有明显的种间差异。
植物能够通过调节生物量分配和植株本身的水分含量保持能力来适应CO2浓度和干旱胁迫的交互影响,这种调节能力取决于植物在碳的吸收和水分散失之间的平衡“trade-off”。
[1] 杨连新, 王云霞, 朱建国, Hasegawa T, 王余龙.开放式空气中CO2浓度增高(FACE)对水稻生长和发育的影响. 生态学报, 2010, 30(6): 1573-1585.
[2] Stocker T F, Qin D, Plattner G K, Tignor M, Allen S K, Boschung J, Nauels A, Yu X, Vincent B, Midgley P M. Climate change 2013: The physical science basis // Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge: Cambridge University Press, 2013.
[3] 王英, 曹明奎, 陶波, 李克让. 全球气候变化背景下中国降水量空间格局的变化特征. 地理研究, 2006, 25(6): 1031-1040.
[4] Bowes G. Facing the inevitable: plants and increasing atmospheric CO2. Annual Review of Plant Physiology and Plant Molecular Biology, 1993, 44(1): 309-332.
[5] 白莉萍, 林而达. CO2浓度升高与气候变化对农业的影响研究进展. 中国生态农业学报, 2003, 11(2): 132-134.
[6] Tissue D T, Lewis J D. Learning from the past: how low [CO2] studies inform plant and ecosystem response to future climate change. New Phytologist, 2012, 194(1): 4-6.
[7] Xu C G, Gertner G Z, Scheller R M. Potential effects of interaction between CO2and temperature on forest landscape response to global warming. Global Change Biology, 2007, 13(7): 1469-1483.
[8] Leakey A D B, Bishop K A, Ainsworth E A. A multi-biome gap in understanding of crop and ecosystem responses to elevated CO2. Current Opinion in Plant Biology, 2012, 15(3): 228-236.
[9] Poorter H, Pérez-Soba M. The growth response of plants to elevated CO2under non-optimal environmental conditions. Oecologia, 2001, 129(1): 1-20.
[10] Atwell B J, Henery M L, Rogers G S, Seneweera S P, Treadwell M, Conroy J P. Canopy development and hydraulic function inEucalyptustereticornisgrown in drought in CO2-enriched atmospheres. Functional Plant Biology, 2007, 34(12): 1137-1149.
[11] Tissue D T, Griffin K L, Thomas R B, Strain B R. Effects of low and elevated CO2on C3and C4annuals. Oecologia, 1995, 101(1): 21-28.
[12] Ward J K, Tissue D T, Thomas R B, Strain B R. Comparative responses of model C3and C4plants to drought in low and elevated CO2. Global Change Biology, 1999, 5(8): 857-867.
[13] Xu Z Z, Zhou G S, Wang Y H. Combined effects of elevated CO2and soil drought on carbon and nitrogen allocation of the desert shrubCaraganaintermedia. Plant and Soil, 2007, 301(1/2): 87-97.
[14] Erice G, Irigoyen J J, Sánchez-Díaz M, Avice J C, Ourry A. Effect of drought, elevated CO2and temperature on accumulation of N and vegetative storage proteins(VSP) in taproot of nodulated alfalfa before and after cutting. Plant Science, 2007, 172(5): 903-912.
[15] Morgan J A, Lecain D R, Mosier A R, Milchunas D G. Elevated CO2enhances water relations and productivity and affects gas exchange in C3and C4grasses of the Colorado shortgrass steppe. Global Change Biology, 2001, 7(4): 451-466.
[16] Luo Y Q, Hui D F, Zhang D Q. Elevated CO2stimulates net accumulations of carbon and nitrogen in land ecosystems: a meta-analysis. Ecology, 2006, 87(1): 53-63.
[17] 王润佳, 高世铭, 张绪成. 高大气CO2浓度下C3植物叶片水分利用效率升高的研究进展. 干旱地区农业研究, 2010, 28(6): 190-195.
[18] 郝兴宇, 李萍, 杨宏斌, 韩雪, 林而达. 大气CO2浓度升高对绿豆生长及C、 N吸收的影响. 中国生态农业学报, 2011, 19(4): 794-798.
[19] 李秀华, 苏文华, 周鸿, 张光飞. 大气二氧化碳倍增对短葶飞蓬生长和有效成分积累的影响. 应用生态学报, 2009, 20(8): 1852-1856.
[20] Duursma R A, Barton C V M, Eamus D, Medlyn B E, Ellsworth D S, Forster M A, Tissue D T, Linder S, McMurtrie R E. Rooting depth explains [CO2]× drought interaction inEucalyptussaligna. Tree Physiology, 2011, 31(9): 922-931.
[21] Smith S D, Charlet T N, Zitzer S F, Abella S R, Vanier C H, Huxman T E. Long-term response of a Mojave Desert winter annual plant community to a whole-ecosystem atmospheric CO2manipulation (FACE). Global Change Biology, 2014, 20(3): 879-892.
[22] 刘锦春, 钟章成, 何跃军. 干旱胁迫及复水对喀斯特地区柏木幼苗活性氧清除系统的影响. 应用生态学报, 2011, 22(11): 2836-2840.
[23] 张昌胜, 刘国彬, 薛萐, 冀智清, 张超. 干旱胁迫和CO2浓度升高条件下白羊草的光合特征. 应用生态学报, 2012, 23(11): 3009-3015.
[24] 许振柱, 周广胜, 肖春旺, 王玉辉. CO2浓度倍增条件下土壤干旱对两种沙生灌木碳氮含量及其适应性的影响. 生态学报, 2004, 24(10): 2186-2191.
[25] 王美玉, 赵天宏, 张巍巍, 史奕. CO2浓度升高与温度、干旱相互作用对植物生理生态过程的影响. 干旱地区农业研究, 2007, 25(2): 99-103.
[26] 高素华, 郭建平, 周广胜. 高CO2浓度下羊草对土壤干旱胁迫的响应. 中国生态农业学报, 2002, 10(4): 31-33.
[27] 王云霞, 杨连新, Manderscheid R, 王余龙. C4作物FACE (free-air CO2enrichment) 研究进展. 生态学报, 2011, 31(5): 1450-1459.
[28] Herrera A, Fernández M D, Taisma M A. Effects of drought on CAM and water relations in plants ofPeperomiacarnevalii. Annals of Botany, 2000, 86(3): 511-517.
[29] 赵彬彬, 牛克昌, 杜国祯. 放牧对青藏高原东缘高寒草甸群落27种植物地上生物量分配的影响. 生态学报, 2009, 29(3): 1596-1606.
[30] 郭建平, 高素华. 沙地植物生长对CO2增加和土壤干旱的响应. 水土保持学报, 2004, 18(4): 174-176.
[31] Mortensen L M. Review: CO2enrichment in greenhouses. Crop response. Scientia Horticulturae, 1987, 33(1/2): 1-25.
[32] Wall G W, Adam N R, Brooks T J, Kimball B A, Pinter P J Jr, Lamorte R L, Adamsen F J, Hunsaker D J, Wechsung G, Wechsung F, Grossman-Clarke S, Leavitt S W, Matthias A D, Webber A N. Acclimation response of spring wheat in a free-air CO2enrichment (FACE) atmosphere with variable soil nitrogen regimes. Net assimilation and stomatal conductance of leaves. 2. Net assimilation and stomatal conductance of leaves. Photosynthesis Research, 2000, 66(1/2): 79-95.
[33] Wechsung G, Wechsung F, Wall G W, Adamsen F J, Kimball B A, Pinter JR P J, Lamorte R L, Garcia R L, Kartschall T H. The effects of free-air CO2enrichment and soil water availability on special and seasonal patterns of wheat root growth. Global Change Biology, 1999, 5(5): 519-529.
[34] 乔匀周, 王开运, 张远彬, 徐燕, 李青超. 红桦幼苗干物质分配与营养对CO2升高的响应. 福建林学院学报, 2007, 27(2): 161-164.
[35] 王海珍, 粱宗锁, 韩蕊莲, 韩路. 土壤干旱对黄土高原4个乡土树种生长及干物质分配的影响. 植物资源与环境学报, 2005, 14(1): 10-15.
[36] 郝树荣, 郭相平, 王为木, 张烈君, 王琴, 王青梅, 刘展鹏. 水稻分蘖期水分胁迫及复水对根系生长的影响. 干旱地区农业研究, 2007, 25(1): 149-152.
[37] 李玉霖, 崔建垣, 苏永中. 不同沙丘生境主要植物比叶面积和叶干物质含量的比较. 生态学报, 2005, 25(2): 304-311.
[38] 于显枫, 张绪成, 王红丽. 高浓度CO2下氮素对小麦叶片干物质积累及碳氮关系的影响. 核农学报, 2012, 26(7): 1058-1063.
Growth responses to the interaction of elevated CO2and drought stress in six annual species
GAO Kaimin1, LIU Jinchun1,*, LIANG Qianhui1, Andries A. TEMME2, Johannes H.C. CORNELISSEN2
1KeyLaboratoryofEco-environmentinThreeGorgesReservoirRegion,ChongqingKeyLaboratoryofPlantEcologyandResourceResearchinThreeGorgesReservoirRegion,SchoolofLifeScience,SouthwestChinaUniversity,Chongqing400715,China2DepartmentofSystemsEcology,InstituteofEcologicalScience,VUUniversityAmsterdam,DeBoelelaan1085, 1081HVAmsterdam,theNetherlands
Carbon dioxide (CO2) is the most important anthropogenic greenhouse gas. Global atmospheric CO2may reach 700 μL/L by 2100 and increase at a high rate of 0.4% per year according to the Intergovernmental Panel on Climate Change. Elevation of atmospheric CO2can have an indirect influence on global climate change through “the greenhouse effect” and a direct influence on plant growth, survival, and reproduction. Therefore, elevated CO2will have a great effect upon structure and function of terrestrial ecosystems and on the distribution and productivity of global vegetation. Global warming has recently resulted in uneven precipitation and more frequent extreme droughts and shortages of available soil water in many areas of the world. In addition, plants have different sensitivities to elevated CO2and drought stress, even when growing in the same environment. Thus it is essential to consider both elevated CO2and different soil moisture conditions in order to assess the possible effect of global climate change on different species. In this paper, the six annual species,Rumexchalepensis,Viciasepium,Sedumalbum,Hemmisteptalyrata,ClinopodiumchinenseandChenopodiumalbum, were treated with two levels of CO2concentrations (400 μL/L and 700 μL/L) and three levels of drought stress (CK: 100% FC (field capacity), MS: 40% FC and SS: 20% FC) in walk-in CO2chambers to determine the responses of growth and biomass allocation to the interaction of elevated CO2and drought stress and to test whether elevated atmospheric CO2ameliorates negative effects of drought by increasing water use efficiency. Results showed thatV.sepiumandS.albumsurvived under all experimental conditions. However, there was some mortality in other species in drought treatments. Elevated CO2stimulated the growth of the five C3plantsR.chalepensis,V.sepium,H.lyrata,C.chinenseandC.albumunder three levels of drought stress, while it inhibited the growth ofS.albumunder drought stress. Drought stress inhibited the growth of all six species, but the growth of the CAM plantS.albumwas fastest under drought stress. The interaction of elevated CO2and drought stress showed significant interspecific variation: elevated CO2concentration alleviated the negative impact of drought onH.lyrataandC.chinensebut less so inR.chalepensisandV.sepium; elevated CO2had no effect on the impact of drought onC.album; forS.album, elevated CO2even inhibited its growth under drought stress. Overall, elevated CO2increased the root mass fraction (RMF) and dry matter content (DMC) for most of the species; drought stress obviously increased RMF and decreased DMC in all six species. However, different species had different responses to the interaction of elevated CO2and drought stress. This indicated that plants could adapt to the interactive effect of elevated CO2and drought stress by regulating biomass allocation and moisture retention capacity, as dependent on the trade-off between carbon absorption and water loss. The results could help us to understand the adaptation of annual herbs to future climatic change and provide a basis for assessing and evaluating global climate change and its hydrological effect on plant physiological ecology.
elevated CO2; drought stress; biomass accumulation and allocation; herbs; field capacity (FC)
中央高校基本科研业务费专项(XDJK2014C158); 重庆市自然科学基金(cstc2014jcyjA80016)
2014-01-21;
日期:2014-11-19
10.5846/stxb201401210159
*通讯作者Corresponding author.E-mail: jinchun@swu.edu.cn
高凯敏, 刘锦春, 梁千慧, Andries A. Temme, Johannes H.C. Cornelissen.6种草本植物对干旱胁迫和CO2浓度升高交互作用的生长响应.生态学报,2015,35(18):6110-6119.
Gao K M, Liu J C, Liang Q H, Andries A. TEMME, Johannes H.C. CORNELISSEN.Growth responses to the interaction of elevated CO2and drought stress in six annual species.Acta Ecologica Sinica,2015,35(18):6110-6119.
