干旱胁迫对桔梗种子萌发及幼苗生长的影响
2015-01-07刘自刚
刘自刚,沈 冰 ,张 雁,3
(1 西安理工大学 水利水电学院,陕西 西安 710048;2 甘肃农业大学 农学院,甘肃省干旱生境作物学质量点试验室/甘肃省作物遗传改良与种质创新质量点试验室,甘肃 兰州730070;3 商洛学院,陕西 商洛 726000)
干旱胁迫对桔梗种子萌发及幼苗生长的影响
刘自刚1,2,沈 冰1,张 雁1,3
(1 西安理工大学 水利水电学院,陕西 西安 710048;2 甘肃农业大学 农学院,甘肃省干旱生境作物学质量点试验室/甘肃省作物遗传改良与种质创新质量点试验室,甘肃 兰州730070;3 商洛学院,陕西 商洛 726000)
【目的】 阐明桔梗种子萌发及幼苗生长对干旱胁迫的形态与生理响应机制。【方法】 以桔梗种子为材料,采用质量分数5%,10%,15%,20%,25%的聚乙二醇(PEG-6000)模拟不同程度的干旱胁迫,以蒸馏水处理为对照,测定干旱胁迫下桔梗种子的发芽特征及幼苗叶片的抗氧化酶活性、渗透调节物质和丙二醛含量等指标,探讨干旱胁迫对桔梗种子萌发和幼苗生长的影响。【结果】 (1)干旱胁迫下桔梗种子初始萌发时间延后,发芽率、发芽势、发芽指数、活力指数、胚芽长、胚根长、胚根长/胚芽长和苗鲜质量等指标均较对照明显降低,且随干旱胁迫程度的加剧,以上指标均呈显著下降趋势;干旱胁迫对桔梗幼苗胚根生长的抑制作用较其对胚芽的影响更为明显。(2)随着干旱胁迫程度的增大,桔梗幼苗叶色逐渐变为暗绿色,叶片卷曲,气孔关闭。(3)质量分数10% PEG-6000处理桔梗幼苗的脯氨酸含量略高于对照,其余各处理的游离脯氨酸含量均低于对照;低质量分数(≤5%)PEG-6000处理桔梗幼苗的可溶性糖含量略低于对照,但高质量分数(≥10%)PEG-6000处理的可溶性糖含量均显著高于对照。(4) 与对照相比,不同质量分数PEG-6000处理后桔梗幼苗叶片的SOD活性均显著升高,而POD活性均显著下降;质量分数5%PEG-6000处理桔梗幼苗叶片的CAT活性较对照略有升高,但差异并不显著,而高质量分数(≥10%)PEG-6000处理叶片的CAT活性较对照显著降低;PEG-6000处理幼苗叶片的MDA含量均显著高于对照。【结论】 干旱胁迫对桔梗种子萌发和幼根、幼芽生长均有明显的抑制作用,但对幼根生长的抑制作用更加明显,轻度干旱胁迫即能显著抑制幼根生长可能是造成桔梗种子田间出苗率低的主要原因之一。
桔梗;种子萌发;幼苗生长;干旱胁迫;生理特性
全球气候变化与局部干旱化导致越来越多的地区受到严重的干旱胁迫影响[1],水分短缺成为这些地区作物生产与植被恢复的关键制约因素[2-3]。大多数植物在干旱胁迫下气孔关闭,叶片水势下降,形态特征及生物量分配格局也发生变化[4-5],使植物机体受到胁迫伤害甚至死亡。植物在长期演化过程中,形成了一套通过调节体内一系列生理生化反应以适应干旱胁迫生境的防御机制[6-7]。
桔梗(Platycodongrandiflorum(Jacq.) A.DC)属桔梗科、桔梗属多年生双子叶草本植物,是一种药用、食用、观赏兼用型植物[8]。种子萌发和幼苗生长阶段是一个植物种群能够在特定环境下定植的关键时期。桔梗种子较为细小,千粒质量平均约为1.21 g[9],桔梗土壤种子库通常分布在2 cm左右的土壤表层。由于春季多风,温度回升快,降雨也逐渐增多,很容易导致表层土壤的桔梗种子处于干湿交替的环境之中,使其在种子萌发阶段遭受干旱胁迫的干扰,进而影响其种群更新过程和群体规模维系。桔梗人工栽培中常由于干旱造成生产田缺苗断垄,是产量形成的主要限制因素之一[10-13],但目前就桔梗种子萌发和幼苗生长对干旱环境的适应能力及适应机制的研究尚未见报道。为此,本试验以聚乙二醇(PEG-6000)作为水分胁迫剂模拟不同程度的干旱胁迫,分析干旱胁迫下桔梗种子的萌发特性以及幼苗生长、形态特征、生物量分配、体内渗透调节能力、抗氧化能力、伤害程度等的变化特征,以期为了解桔梗幼苗对干旱生境的适应机制提供参考。
1 材料与方法
1.1 材 料
供试桔梗种子于2008-10采自陕西香菊制药有限公司商洛植物药园2年生栽培群体,选取深黑色大小基本一致的饱满种子备用。PEG-6000购自华大生物公司。
1.2 方 法
1.2.1 种子萌发试验 选取桔梗优质种子用质量分数0.4%的KMnO4溶液表面消毒,蒸馏水冲干净,培养皿垫双层滤纸作为发芽床,将供试种子分别放入加有等量质量分数5%,10%,15%,20%,25%聚乙二醇溶液(PEG-6000) 和蒸馏水(对照)的发芽床中,每皿100粒,准确称质量后放入培养箱中于25 ℃下进行光培养,每天观察统计种子萌发情况,并根据实际情况添加蒸馏水至原始质量,每处理重复5次。将胚根与种子等长视为种子发芽标准,8 d统计发芽势,11 d统计发芽率,发芽结束后计算发芽指数和活力指数。相关指标计算公式如下:
种子发芽率(%)=n/N×100%;
种子相对发芽率(%)=(PEG处理组发芽数/对照组发芽数)×100%;
发芽势=规定时间内发芽种子粒数/供试种子粒数×100%;
发芽指数(GI)=∑Gt/Dt;
活力指数(VI)=发芽指数(GI)×S。
式中:n为发芽终期全部正常发芽的种子数,N为供试种子总数;Gt为t日发芽数,Dt为相应的天数;S为胚根平均鲜质量。
在种子萌发试验进行12 d后,每重复各取10株测量其胚根和胚芽长度,计算平均值。同时称取幼苗鲜质量,以发芽率为对照10%、50%时的PEG-6000质量分数分别作为种子抗旱极限质量分数和种子抗旱半致死质量分数。
1.2.2 幼苗干旱处理 将石英砂、蛭石和有机质土按质量比1∶1∶2混合均匀配成基质,称取等量基质分装于盆钵组成培养床,选取发育良好的桔梗种子播种于基质中,每杯播种20粒种子,添加适量的蒸馏水后称质量,以后每天根据蒸发量添加蒸馏水。将播种的盆钵放入人工气候箱中进行培养 (温度28 ℃,光照时间24 h),待子叶离开土面1 cm后,在28 ℃(昼)/18 ℃(夜)条件下进行变温培养,每天光照12 h,光强500 μmol/(m2·s)。待第1对真叶展开后定苗,每杯选取发育良好且大小一致的幼苗10株继续培养。
幼苗展开3对真叶后进行模拟干旱处理,处理前7 d停止浇水,使土壤含水量下降到田间持水量的40%左右,此时盆土的实际质量含水量约为16%。分别添加质量分数为5%,10%,15%,20%,25%的PEG-6000溶液,然后将盆钵放在28 ℃下让土壤水分不断蒸发,每天用称质量法补充PEG-6000溶液至原质量,2 d后水分蒸发量达到盆土质量的16%左右,此时作为干旱处理的起始时间。干旱处理7 d后观察幼苗的形态反应和干旱表现,以蒸馏水处理为对照,测定桔梗叶片的各项生理指标。
1.3 测定指标及方法
可溶性糖含量采用蒽酮比色法测定,游离脯氨酸含量采用酸性茚三酮比色法测定[14],超氧化物歧化酶(Superoxide dismultase,SOD)活性采用氮蓝四唑(NBT)光化还原法测定,过氧化物酶(Peroxidase,POD)活性采用愈创木酚法测定,过氧化氢酶(Catalase,CAT)活性采用紫外吸收法测定,丙二醛(Malondialdehyde,MDA)含量采用硫代巴比妥酸(TBA)反应法测定[15]。试验设置3次重复。气孔观察采用指甲油印膜法[16],将功能叶片下表皮取下制成临时切片,光学显微镜下观察气孔特征。
干旱处理期间观察叶片形态、气孔状态等变化情况,根据幼苗形态特征按4级标准记载和评级,级数越小,则受干旱胁迫危害越轻。幼苗形态特征的4级标准为:1级.无受害症状,气孔开放;2级.小部分叶片萎缩,并失去应有光泽,有较少的叶片卷曲,气孔半开放,有少数气孔关闭;3级.大部分叶片萎缩卷曲,多数气孔关闭;4级.叶片卷缩严重,颜色显著深于该品种正常颜色,下部叶片开始变黄,气孔全部关闭。
1.4 数据处理
利用SPSS软件对试验数据进行统计与分析。
2 结果与分析
2.1 干旱胁迫对桔梗种子发芽起始时间的影响
根据观察记录,在萌发培养6 d时,对照(5个重复)种子均开始大量萌发,萌发种子数平均为21.6;培养皿中加质量分数5% PEG-6000溶液的种子也开始萌发,但萌发种子数明显少于对照,其发芽种子数平均为15.0;质量分数10% PEG-6000溶液处理仅2个重复有种子萌发;质量分数15% PEG-6000溶液处理培养8 d时仅1个重复有种子萌发;质量分数20% PEG-6000溶液处理仅1个重复在培养11 d时才有种子萌发;质量分数25% PEG-6000溶液处理一直未见种子萌发。
2.2 干旱胁迫对桔梗种子萌发的影响
发芽率反映了种子在适宜条件下的萌发能力,活力指数是种子活力水平的综合体现。不同质量分数PEG-6000对桔梗种子萌发的影响见表1。从表1可以看出,随着PEG处理质量分数的增大(即干旱胁迫程度的加深),桔梗种子发芽率、相对发芽率、发芽势、发芽指数和活力指数均呈下降趋势。当PEG质量分数增加到20%时,发芽势和活力指数分别由对照的26.8%和3.04下降为0;当PEG质量分数升高至25%时,所有发芽指标均下降为0,表明质量分数20% PEG-6000已完全抑制了桔梗种子的萌发。
从表1还可以看出,PEG-6000试验组种子各萌发指标均低于对照,但不同质量分数PEG-6000对桔梗种子萌发的影响效应不同,低质量分数(≤5%)PEG-6000试验组种子发芽势较对照显著降低,而其他各萌发指标较对照略有降低,差异均未达到显著水平;高质量分数(≥10%)PEG-6000处理的各萌发指标均显著低于对照。质量分数10%和15% PEG-6000试验组种子发芽率分别为对照发芽率的56.5%和14.1%,表明桔梗种子抗旱半致死PEG-6000质量分数和抗旱极限PEG-6000质量分数分别在10%和15%以上。
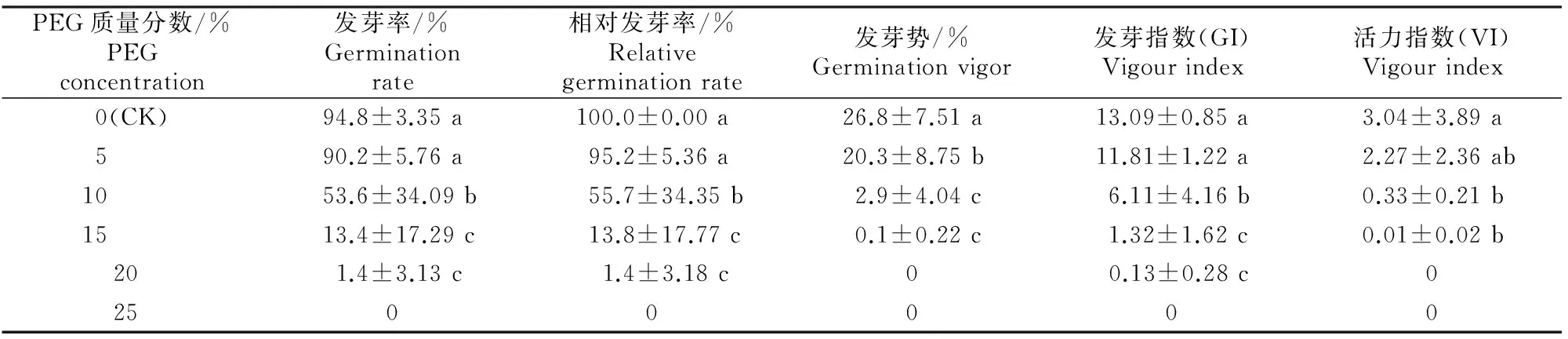
表1 干旱胁迫对桔梗种子萌发的影响Table 1 Effect of drought stress on seed germination of Platycodon grandiflorum
注:同列数据后标不同小写字母表示各处理间在P<0.05水平差异显著。下表同。
Note:Different lowercase letters in each column show significant difference among different PEG concentrations atP<0.05.The same below.
2.3 干旱胁迫对桔梗胚根和胚芽生长的影响
干旱胁迫对桔梗幼苗生长的影响情况见表2。从表2可以看出,PEG-6000试验组桔梗幼苗鲜质量、胚芽长、胚根长和胚根长/胚芽长均较对照明显降低,但不同质量分数PEG-6000对桔梗幼苗生长指标的影响效果不同,低质量分数(≤5%)PEG-6000处理幼苗的胚芽长、胚根长/胚芽长和幼苗鲜质量3个指标均较对照略有降低,但差异不显著,而此时胚根长度已显著变短,表明轻度干旱胁迫即可显著抑制桔梗幼苗的胚根生长,幼苗胚根长/胚芽长的下降主要是由胚根长度降低所导致。
由表2可知,随着PEG-6000质量分数的增大,桔梗幼苗各生长指标均呈下降趋势;胚芽长、胚根长和胚根长/胚芽长3个指标在不同质量分数PEG-6000处理间均存在显著差异,而PEG-6000质量分数的增大对幼苗鲜质量无显著影响。

表2 干旱胁迫对桔梗幼苗生长的影响Table 2 Effect of drought stress on seedling growth of Platycodon grandiflorum
2.4 干旱胁迫对桔梗幼苗形态特征的影响
水分亏缺常常引起植物形态特征的相应改变,特别是植物叶片特征的改变更为明显。干旱胁迫对桔梗幼苗形态特征的影响见表3。

表3 干旱胁迫对桔梗幼苗形态特征的影响Table 3 Effect of drought dress on morphological characteristics of Platycodon grandiflorum seedling
由表3可以看出,与对照相比,低质量分数(≤5%)PEG-6000处理桔梗幼苗叶色亮绿、气孔开放正常,无明显受害症状;但随着PEG-6000质量分数的增大, 叶片颜色逐渐加深,色泽也逐渐由亮变暗,叶面逐渐变小,卷曲程度逐渐加重,气孔开度逐渐变小直至完全关闭。模拟干旱7 d后,质量分数10% PEG-6000处理幼苗叶片的多数气孔呈半开状态,质量分数15% PEG-6000处理叶片的气孔已有部分完全关闭,而质量分数20% PEG-6000处理叶片的所有气孔基本处于完全关闭状态,以减少植物体内水分的散失。
2.5 干旱胁迫对桔梗幼苗渗透调节物质的影响
游离脯氨酸和可溶性糖在植物体内具有广泛的生理作用,也是2种重要的渗透调节物质,参与调节细胞水势的变化,表4为桔梗幼苗游离脯氨酸和可溶性糖含量在干旱胁迫下的变化情况。
表4 干旱胁迫对桔梗幼苗渗透调节物的影响
Table 4 Effect of drought dress on contents of osmotic regulation substances in Platycodon grandiflorum leaves

PEG质量分数/%PEGconcentration游离脯氨酸/(μg·mL-1)Freeprolineconten可溶性糖/(μg·mL-1)SolublesugarcontentPEG质量分数/%PEGconcentration游离脯氨酸/(μg·mL-1)Freeprolineconten可溶性糖/(μg·mL-1)Solublesugarcontent0(CK)3.56±0.32a186.9±9.23d153.02±0.17b296.5±9.03a53.40±0.41ab175.9±7.96d203.06±0.22b233.4±7.58c103.97±0.28a274.8±8.54b252.99±0.15b222.3±8.21c
由表4可以看出,随着PEG-6000质量分数的增加,桔梗幼苗中的游离脯氨酸和可溶性糖含量均呈“单峰”曲线变化,当PEG-6000质量分数为10%时游离脯氨酸含量达最大值,为3.97 μg/mL,PEG-6000质量分数为15%时可溶性糖含量达到最大值,为296.5 μg/mL,之后随PEG-6000质量分数继续增大其含量开始下降。与对照相比,10% PEG-6000处理幼苗的脯氨酸含量略高于对照,5%处理的脯氨酸含量略低于对照,但差异均不显著;高质量分数(≥15%)PEG-6000处理幼苗的游离脯氨酸含量均显著低于对照。质量分数5% PEG-6000处理桔梗幼苗的可溶性糖含量略低于对照,而用质量分数高于10% 的PEG-6000处理后其含量显著高于对照。
2.6 干旱胁迫对桔梗幼苗抗氧化酶系统的影响
干旱胁迫处理后桔梗幼苗叶片SOD、POD和CAT 3种抗氧化酶活性及脂质过氧化终产物MDA含量的变化情况见表5。

表5 干旱胁迫对桔梗幼苗抗氧化酶活性和丙二醛含量的影响Table 5 Effect of drought stress on activities of antioxidant enzymes and content of MDA in Platycodon grandiflorum leaves
从表5可以看出,与对照相比,不同质量分数PEG-6000处理后,桔梗幼苗叶片的SOD活性均显著升高,而POD活性均显著下降,低质量分数(≤5%)PEG-6000处理后叶片CAT活性较对照略有升高,但差异不显著,较高质量分数(≥10%)PEG-6000处理叶片的CAT活性较对照显著降低;高质量分数(≥10%)PEG-6000处理幼苗叶片的MDA含量均显著高于对照。
随干旱胁迫程度的增加,CAT活性呈明显下降趋势;SOD、POD活性及MDA含量均呈“单峰”曲线变化,即随PEG-6000处理质量分数的增加,这3个指标均先呈上升趋势,达到最大值后又开始下降。
3 讨 论
干旱胁迫是植物生长的逆境之一,植物在干旱胁迫条件下,活性氧等自由基的增加引起膜脂过氧化,并可使细胞膜结构及其生理完整性遭到破坏[17-18],抗氧化酶系统可有效清除活性氧自由基,缓解干旱胁迫对植物造成的伤害。抗氧化酶活力的变化是影响植物体内自由基生成与清除平衡的重要因素,植物对干旱胁迫抗性的形成是几种抗氧化酶协同作用的结果。干旱胁迫下,五爪金龙[19]、飞机草[20]、枣树[21]幼苗SOD活性上升,CAT活性下降;空心莲子草[22]的POD活性在轻、中度干旱胁迫时上升,在重度胁迫时下降。桔梗幼苗SOD活性变化与上述研究结果基本相同,即在干旱胁迫下桔梗幼苗的SOD活性显著上升;而POD活性显著下降,CAT活性在轻度胁迫时上升,中度、重度胁迫时下降,这与前人研究结果[19-22]略有不同,其原因可能是不同植物幼苗对干旱胁迫应答的生理特征和机制有所差异。一般认为,MDA在植物体内的积累是活性氧毒害的表现,其含量是判断膜脂过氧化程度的一个重要指标[23]。许多研究表明,干旱胁迫下植物体内会积累MDA[19-22,24-25],桔梗幼苗受到干旱胁迫后体内同样也会积累MDA,表明在干旱胁迫下桔梗幼苗总体朝生理劣变方向发展。
游离脯氨酸和可溶性糖在植物体内具有广泛的生理作用,不仅可为植物的生命活动提供能量,为其他物质的合成提供中间产物,同时还是细胞的重要渗透调节物质,其含量的高低可以影响细胞水势,从而控制植物水分的进出。焦树英等[24]、付秋实等[25]研究认为,干旱胁迫下狼尾草、辣椒细胞内游离脯氨酸含量明显升高,抗旱性能增加。本研究结果显示,桔梗幼苗在轻度、中度干旱胁迫下游离脯氨酸含量变化不明显,在重度干旱胁迫下游离脯氨酸含量较对照显著降低,而可溶性糖含量显著升高,以相对提高幼苗对干旱胁迫的适应能力。
研究表明,轻度、中度干旱胁迫对一些植物种子的胚根生长影响不明显[19,26],而桔梗种子遇到较低程度干旱胁迫时即表现为发芽率降低,胚根生长显著被抑制而伸长缓慢,幼根不能迅速深入深层土壤吸收水分,造成桔梗种子萌发及幼苗阶段对水分亏缺十分敏感,较低程度的干旱胁迫即可导致桔梗幼苗生长困难,为了弥补这种生存缺陷,桔梗种子采取了“避害”的生存策略。桔梗种子千粒质量仅为1.21 g左右,能供给萌发和胚芽、胚根伸长的种子贮藏物质十分有限,在正常水分条件下,桔梗种子萌发结束后其幼苗根茎比为5.72,远高于其他植物[19,24-25,27-28],这表明当表层土壤较为湿润时,萌发后的桔梗种子能迅速调动其贮藏物质优先供给胚根生长,使胚根在较短的时间内迅速伸长并伸入深层土壤吸取土壤深层水分,从而降低桔梗种子萌发阶段土壤表层干旱胁迫的危害。
4 结 论
随着干旱胁迫程度的加深,桔梗种子萌发起始时间延迟,幼苗叶色逐渐变为灰绿色、叶片发生卷曲,气孔逐渐关闭,受害程度加深,叶片生理状态明显劣变;幼芽和幼根生长均被明显抑制,但幼根生长对水分亏缺更为敏感,这可能是干旱生境下桔梗种子田间出苗率低的主要原因。
[1] Intergovernmental Panel on Climate Change.Climate change:The scientific basis [M]//Hougton J T,Dung Y,Griggs D J,et al.Contribution of working group Ⅰ:Third assessment report of intergovernmental panel on climate change.Cambridge:Cambridge University Press,2007:1.
[2] Bouwan A F,Van Vuuren D P,Derwent R G,et al.A global analysis of acidification and eutrophication of terrestrial ecosystems [J].Water,Air and Soil Pollution,2002,141:349-382.
[3] Bosabalidis A M,Kofidis G.Comparative effects of drought str-ess on leaf anatomy of two olive cultivars [J].Plant Science,2002,163:375-379.
[4] Patnaitk D,Khurana P.Wheat biotechnology:Amini-review [J].Electronic Journal of Biotechnology,2002,4:74-102.
[5] 初晓辉,刀丽琼,谢堂民,等.马蹄金草坪草对干旱胁迫的生理响应研究 [J].草地学报,2014,22(1):134-138.
Chu X H,Dao L Q,Xie T M,et al.Physiological response of creeping dichondra herb to drought stress [J].Acta Agrestia Sinica,2014,22(1):134-138.(in Chinese)
[6] 邬佳宝,马明科,张 刚,等.文冠果对干旱胁迫的光合生理响应 [J].干旱地区农业研究,2014,32(5):55-60.
Wu J B,Ma M K,Zhang G,et al.Photosynthetic physiological response ofXanthocerassorbifoliaunder drought stress [J].Agricultural Research in the Arid Areas,2014,32(5):55-60.(in Chinese)
[7] 张金政,张起源,孙国峰,等.干旱胁迫及复水对玉簪生长和光合作用的影响 [J].草业学报,2014,23(1):167-176.
Zhang J Z,Zhang Q Y,Sun G F,et al.Effects of drought stress and re-watering on growth and photosynthesis of Hosta [J].Acta Prataculturae Sinica,2014,23(1):167-176.(in Chinese)
[8] 刘自刚,张 雁,王新军,等.桔梗育种研究进展 [J].中草药,2006,37(6):962-964.
Liu Z G,Zhang Y,Wang X J,et al.Research progress ofPlatycodongrandiflorum(Jacq.)A.DC breeding [J].Chinese Traditional and Herbal Drugs,2006,37(6): 962-964.(in Chinese)
[9] 刘自刚,呼天明,杨亚丽.桔梗种子的成熟生理动态研究 [J].中草药,2009,40(2):300-303.
Liu Z G,Hu T M,Yang Y L.Ripening physiological characteristics on seeds ofPlatycodongrandiflorum[J].Chinese Traditional and Herbal Drugs,2009,40(2):300-303.(in Chinese)
[10] 严一字, 吴基日. 桔梗种子质量对发芽、出苗及苗素质的影响 [J].中草药,2007,38(11):1726-1729.
Yan Y Z,Wu J R.Effect ofPlatycodongrandiflorumseed quality on germination,seedling,and seedling quality [J].Chinese Traditional and Herbal Drugs,2007,38(11):1726-1729.(in Chinese)
[11] 谭玲玲,胡正海.桔梗根的发育解剖学研究 [J].西北植物学报, 2014,34(5):938-942.
Tan L L,Hu Z H.Developmental and anatomical studies on the root ofPlatycodongrandiflorumA.DC [J].Acta Bot Bore-Occident Sin,2014,34(5):938-942.(in Chinese)
[12] 刘自刚.桔梗种子休眠解除方法研究 [J].种子,2009,28(1):72-74.
Liu Z G.Studies on seed dormancy breaking ofPlatycodon[J].Seed,2009,28(1):72-74.(in Chinese)
[13] 郭巧生,赵荣梅,刘 丽,等.桔梗种子发芽特性的研究 [J].中国中药杂志,2006,31(1):870-881.
Guo Q S,Zhao R M,Liu L,et al.Studies on gerrmination characteristics ofPlatycodongrandiflorumseed [J].China Journal of Chinese Materia Medica,2006,31(1):870-881.(in Chinese)
[14] 陈俊辉,陶 力,朱婉华,等.生物化学试验 [M].3版.北京:科学出版社,2003.
Chen J H,Tao L,Zhu W H,et al.Biochemistry experiment [M].3rd ed.Beijing:Science Press,2003.(in Chinese)
[15] 张志良,瞿伟菁.植物生理学试验指导 [M].3版.北京:高等教育出版社,2004.
Zhang Z L,Zhai W Q.Plant physiology labora [M].3rd ed.Beijing:Academic Press,2004.(in Chinese)
[16] Dami I,Hughes H.Leaf anatomy and water loss ofinvitroPEG-treated ‘Valiant’ grape [J].Plant Cell,Tissue and Organ Culture,1995,42:179-184.
[17] 邓明华,文锦芬,邹学校,等.辣椒细胞质雄性不育系的物质代谢和过氧化物酶分析 [J].云南农业大学学报,2007,22(6):791-794.
Deng M H,Wen J F,Zou X X,et al.A study on material metabolism and peroxidase in cytoplasmic male-sterile lines and their maintainer lines of hot pepper [J].Journal of Yunnan Agricultural University,2007,22(6):791-794.(in Chinese)
[18] 王荣华,石 雷,汤庚国,等.渗透胁迫对蒙古冰草幼苗保护酶系统的影响 [J].植物学通报,2003,20(3): 330-335.
Wang R H,Shi L,Tang G G,et al.Effect of osmotic stress on activities of protective enzymes system inAgropyronmongolicumseedling [J].Chinese Bulletin of Botany,2003,20(3):330-335.(in Chinese)
[19] 朱 慧,马瑞君,吴双桃,等.干旱胁迫对五爪金龙种子萌发与幼苗生长的影响 [J].西北植物学报,2009,29(2):344-349.
Zhu H,Ma R J,Wu S T,et al.Seed germination and seedling growth of invasive weedIpomoeacairicaunder drought stress [J].Acta Bot Boreal-Occident Sin,2009,29(2):344-349.(in Chinese)
[20] 鲁 萍,桑卫国,马克平.外来入侵种飞机草在不同环境胁迫下抗氧化酶系统的变化 [J].生态学报,2006,26(11):3578-3585.
Lu P,Sang W G,Ma K P.Activity of antioxidant enzymes in the invasive plantEupatoriumodoratumunder various environmental stresses [J].Acta Ecologica Sinica,2006,26(11):3578-3582.(in Chinese)
[21] 刘世鹏,刘济明,陈宗礼,等.模拟干旱胁迫对枣树幼苗的抗氧化系统和渗透调节的影响 [J].西北植物学报,2006,26(9):1781-1787.
Liu S P,Liu J M,Chen Z L,et al.Effects of simulated drought on antioxidative enzymes and osmotic regulation inZizyphusjujubaseedlings [J].Acta Bot Borea-Occident Sin,2006,26(9):1781-1787.(in Chinese)
[22] 肖 强,高建明,罗立廷,等.干旱胁迫对空心莲子草抗氧化酶活性和组织学的影响 [J].生物技术通讯,2006,17(4):556-559.
Xiao Q,Gao J M,Luo L Y,etal.Effects of drought stress on the activities of antioxidant enzymes and histology inAlternantheraphiloxeroides[J].Letters in Biotechnology,2006,17(4):556-559.(in Chinese)
[23] 郝树荣,郭相平,王文娟.不同时期水分胁迫对玉米生长的后效性影响 [J].农业工程学报,2010,26(7):71-75.
Hao S R,Guo X P,Wang W J.Afte reffects of water stress on corn growth at different stages [J].Transactions of the CSAE,2010,26(7):71-75.(in Chinese)
[24] 焦树英,李永强,沙依拉,等.干旱胁迫对3种狼尾草种子萌发和幼苗生长的影响 [J].西北植物学报,2009,29(2):308-313.
Jiao S Y,Li Y Q,Sha Y L,et al.Seeds germination and seedling growth about 3Pennisetumornamentalgrasses under drought stress [J].Acta Bot Boreal-Occident Sin,2009,29(2):308-313.(in Chinese)
[25] 付秋实,李红岭,崔 健,等.水分胁迫对辣椒光合作用及相关生理特性的影响 [J].中国农业科学,2009,42(5):1859-1866.
Fu Q S,Li H L,Cui J,et al.Effects of water stress on photosynthesis and associated physiological characters ofCapsicumannuumL. [J].Scientia Agricultura Sinica,2009,42(5):1859-1866.(in Chinese)
[26] 徐洪伟, 陆静梅,周晓馥.水分胁迫条件下玉米毛状根再生植株耐旱性研究 [J].农业工程学报,2007,23(7):19-23.
Xu H W,Lu J M,Zhou X F.Drought resistance of regenerative plants from hairy root cultures in maize under water stress [J].Transactions of the CSAE,2007,23(7):19-23.(in Chinese)
[27] 张寄阳,段爱旺,孟兆江,等.不同水分状况下棉花茎直径变化规律研究 [J].农业工程学报,2005,21(5):7-11.
Zhang J Y,Duan A W,Meng Z J,et al.Stem diameter variations of cotton under different water conditions [J].Transactions of the CSAE,2005,21(5):7-11.(in Chinese)
[28] 李文娆,张岁岐,山 仑.水分胁迫下紫花苜蓿和高梁种子萌发特性及幼苗耐旱性 [J].生态学报,2009,29(6):3066-3074.
Li W Y,Zhang S Q,Shan L.Seeds germination characteristics and drought-tolerance of alfalfa and sorghum seedling under water stress [J].Acta Ecologica Sinica,2009,29(6):3067-3074.(in Chinese)
Effects of drought stress on seed germination and seedling growth ofPlatycodongrandiflorum(Jacq.)A.DC
LIU Zi-gang1,2,SHEN Bing1,ZHANG Yan1,3
(1InstituteofWaterResourcesandHydro-electricEngineering,Xi’anUniversityofTechnology,Xi’an,Shaanxi710048,China;2GansuKeyLaboratoryofCropImprovementandGermplasmEnhancement/KeyLaboratoryofAridLandCropScienceinGansuProvince,CollegeofAgronomy,GansuAgriculturalUniversity,Lanzhou,Gansu730070,China;3ShangluoUniversity,Shangluo,Shaanxi726000,China)
【Objective】 The research investigated the effects of drought stress on seed germination and seedling growth ofPlatycodongrandiflorum(Jacq.)A.DC.【Method】 Indexes such as germination ratio,germination vigor,germination index,and vigor index ofPlatycodongrandiflorumseed and free proline content,soluble sugar content,activities of antioxidant enzymes,and mass fraction of MDA in Platycodon leaves were determined and compared by treating the seeds with PEG-6000 at different concentrations of 5%,10%,15%,20%,and 25%.The effects of drought stress on seed germination and seedling growth were also studied.【Result】 With drought stress,initial germination time of seeds was delayed.Germination ratio,germination vigor,germination index and vigor index of seed as well as radicle length,embryonic bud length,root/shoot ratio,and fresh weight of seedling were significantly lower than the control.With the increase of drought stress,these index reduced fatherly.Drought stress could more clearly inhibit the growth of radicle than the growth of embryonic bud.With the increase of drought stress level,thePlatycodongrandiflorumseedling leaves gradually turned into dark green,leaf curled,and stomatal closed.The mass fraction of free proline with 10% PEG-6000 stress was slightly higher than the control while the fractions of other PEG-6000 stress levels were lower than the control.The content of soluble sugar was decreased by low mass fraction PEG-6000(≤5%) stress while it was improved by high mass fraction PEG-6000(>5%)stress.Compared with the control,activity of SOD in seedling leaves was significantly increased with different drought stress degrees,while activity of POD was significantly decreased.Activity of CAT was slightly enhanced by low mass fractions,but it was reduced by high mass fractions (≥10%).Content of MDA was higher than that of the control.【Conclusion】 Drought stress significantly inhibited seed germination and growth of radicle and bud.The inhibition of radical growth by drought stress could be one of the reasons why the germination ratio ofPlatycodongrandiflorumseeds in field was low.
Platycodongrandiflorum;seeds germination;seedling stage;drought stress;physiological characteristics
时间:2015-09-09 15:41
10.13207/j.cnki.jnwafu.2015.10.022
2014-12-10
陕西教育厅专项科研计划项目(2013JK0718)
刘自刚(1975-),男,甘肃天水人,副教授,主要从事植物种质资源及育种研究。 E-mail:lzgworking@163.com
S567.23+9;Q945.6
A
1671-9387(2015)10-0162-07
网络出版地址:http://www.cnki.net/kcms/detail/61.1390.S.20150909.1541.044.html
