利用PHYSAT方法反演南海浮游植物优势类群分布的季节变化
2015-01-03李月洋
李月洋,孙 群,王 磊,魏 皓
(1.天津科技大学海洋科学与工程学院,天津 300457;2.天津大学海洋科学与技术学院,天津 300072)
利用PHYSAT方法反演南海浮游植物优势类群分布的季节变化
李月洋1,孙 群1,王 磊1,魏 皓2
(1.天津科技大学海洋科学与工程学院,天津 300457;2.天津大学海洋科学与技术学院,天津 300072)
基于水色遥感数据用PHYSAT算法反演了南海浮游植物优势类群分布的季节变化,结果表明:微型真核生物作为优势类群全年出现几率最大,原绿球藻次之,聚球藻再次之,硅藻最小;硅藻为优势类群的区域主要在近岸和陆架区,外海原绿球藻和聚球藻的优势度增加.在季节差异上,冬季微型真核生物为优势类群的区域在整个南海海区所占面积的百分比最高,春季原绿球藻比例最高,聚球藻呈双峰结构,在春季和秋季出现峰值,硅藻为优势类群区域的季节性差异较小.陆架、陆坡和海盆区浮游植物优势类群比例的季节变化相近,都表现为冬季微型真核生物的绝对优势和春季原绿球藻优势的大大增加.营养盐浓度分布的季节变化是引起浮游植物优势类群改变的主要影响因子,随着海区中营养盐浓度的降低,浮游植物优势类群由粒径较大的微型真核浮游生物向微微型浮游植物聚球藻和原绿球藻转变.
浮游植物优势类群;PHYSAT算法;水色遥感数据;WOA营养盐浓度;南海
浮游植物作为海洋生态系统的初级生产者,对全球碳循环具有重要作用[1],不同种类的浮游植物具有特殊的生态功能,认识和了解浮游植物优势类群组成的时空变化对深刻理解海洋生态系统结构与功能具有重要意义.
遥感反演具有实时、同步和大尺度的优点,在海洋学研究中应用于生物量的估计[2]、探讨引起赤潮的原因[3-4]、海洋浮游植物类群分布研究中[5],对浮游植物类群分布的反演建立了多种遥感反演算法.PHYSAT算法能够反演4种浮游植物类群占优时的分布,在许多海域得到应用[6].本文尝试将该算法应用于南海空间范围为105°E~125°E、0°N~25°N的海域,并以历史调查资料进行验证,然后给出南海浮游植物优势类群的季节变化和空间分布.浮游植物生长受水温、光和营养盐(氮、磷、硅)结构的影响,南海近岸、陆架、陆坡营养盐浓度梯度更加显著.因此,本研究结合WOA13(World Ocean Atlas 2013)营养盐浓度分布数据,初步探讨影响南海浮游植物优势类群时空变化的主要影响因子.
1 资料与方法
PHYSAT算法根据硅藻、微型真核生物、聚球藻和原绿球藻这4种类群的光谱特征对其中的优势类群进行识别,优势类群指色素贡献量超过60%,[7]的种类.为了提高遥感数据质量,需首先利用气溶胶光学厚度和叶绿素a浓度(cChla)剔除不合格数据,并将离水辐亮度(nLw)数据按公式(1)转变为比归一化离水辐亮度(nLw*).
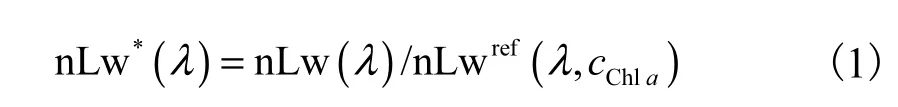
其中nLwref是对应于SeaWiFS卫星标准叶绿素a浓度的nLw平均值,λ是SeaWiFS卫星上波段的中心波长,分别为412、443、490、510和555,nm.依据原文中给出对应的nLw*范围标准,判断是何种浮游植物类群占优.
本文研究区域地形图如图1所示,收集105°E~125°E、0°N~25°N范围内,1998—2010年间日均SeaWiFS卫星离水辐亮度数据、气溶胶光学厚度数据、叶绿素a浓度数据(http://oceancolor.gsfc.nasa.gov/),空间分辨率为9,km,每个像素点代表9,km× 9,km的面积.利用日均nLw数据得到每一天浮游植物优势类群分布结果,对于某一位置,统计某一月份所有在此处占优概率最大的浮游植物类群,作为这一位置当月的优势浮游植物类群.

图1 研究区域地形图Fig.1 Topographic map of the studied area
营养盐浓度源于WOA13(http://www.nodc.noaa.gov/OC5/woa13/).数据空间分辨率为1°×1°.
2 反演结果及验证
2.1 南海浮游植物优势类群分布季节变化
图2为13年(1998—2010年)平均南海浮游植物优势类群的分布图,灰度从浅到深依次代表微型真核生物、原绿球藻、聚球藻和硅藻,白色代表此处无浮游植物类群占优.
图3统计了浮游植物优势类群面积与研究海区面积的百分比,以下简称百分比.分别以12月—2月、3月—5月、6月—8月、9月—11月为冬、春、夏、秋的月份,各季节分别统计浮游植物优势类群在南海所占面积百分比都表现为微型真核生物>原绿球藻>聚球藻>硅藻.冬季,微型真核生物所占百分比在全年最高(60%,~88%,),最高可达88%,,原绿球藻(10%,~36%,)和聚球藻(1%,~5%,)所占百分比在全年最低,硅藻在冬季所占百分比为1%,~2%,;春季,微型真核生物所占百分比在全年最低(40%,~49%,),原绿球藻(37%,~50%,)和聚球藻(5%,~13%,)所占百分比在全年最高,春季4月份原绿球藻所占百分比(50%,)高于微型真核生物(40%,),硅藻在春季3个月份所占百分比均约为1%,;夏季,微型真核生物所占百分比(59%,~63%,)较春季增加,原绿球藻(26%,~30%,)和聚球藻(8%,~10%,)较春季减少,硅藻在夏季所占百分比为1%,~2%,;秋季,与夏季分布较为接近,微型真核生物(56%,~74%,)和聚球藻(4%,~14%,)所占百分比(56%,~74%,)较夏季略有增加,原绿球藻(20%,~27%,)较夏季略有减少,硅藻在秋季所占百分比为2%,~3%,.
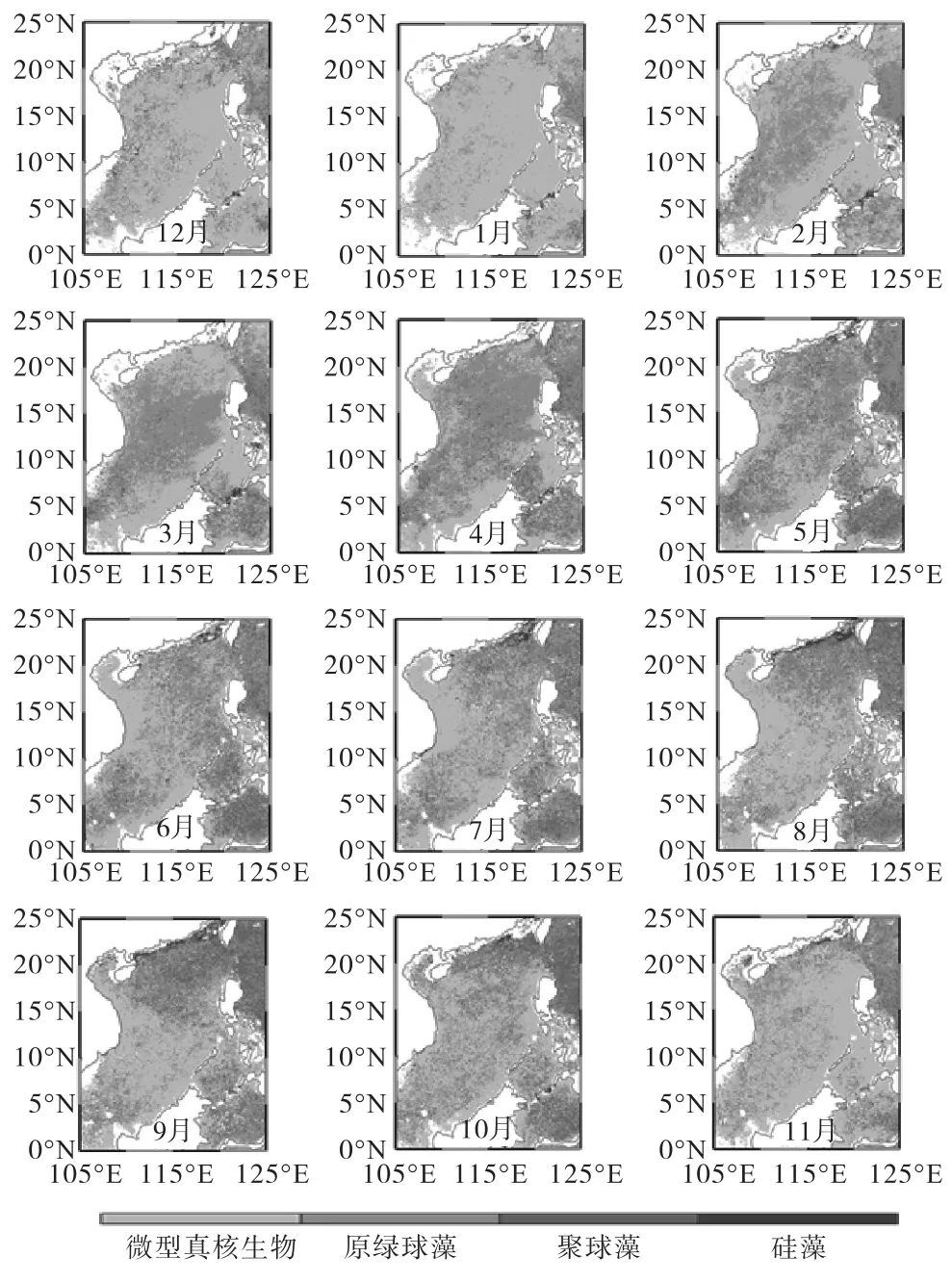
图2 南海13年(1998—2010年)平均各月份浮游植物优势类群分布图Fig.2 Monthly average(1998-2010)of the dominant phytoplankton groups

图3 各浮游植物优势类群出现面积占南海总面积百分比Fig.3 The percentage of each area of dominant phytoplankton groups against the total area of the South China Sea
按照近岸(水深0~50,m)、陆架(水深50~200,m)、陆坡(水深200~1,000,m)和海盆(水深大于1,000,m)来分析浮游植物优势类群的空间变化,图4统计了浮游植物优势类群面积与分区域面积的百分比.近岸的浮游植物优势类群分布最为特殊,大粒径成分高于外海,陆架的浮游植物优势类群分布与陆坡和海盆区接近,陆坡和海盆区的浮游植物优势类群分布非常相似.近岸区,微型真核生物和硅藻所占百分比在4个区域中最高,微型真核生物所占百分比(63%,~82%,)为4个类群中最高,硅藻次之(7%,~19%,),原绿球藻所占百分比(5%,~12%,)略高于聚球藻(3%,~9%,),硅藻表现出了季节差异,在下半年保持较大优势度,在秋季出现峰值(19%,),微型真核浮游植物优势度与之互补;陆架区,微型真核生物(49%,~81%,)和硅藻(1%,~4%,)在陆架所占百分比相较于近岸减少,原绿球藻(15%,~37%,)和聚球藻(2%,~15%,)所占百分比增加;陆坡区,微型真核生物(40%,~86%,)所占百分比相较于陆架在春季减少,其他季节增加,总体上百分比相差不大,原绿球藻(12%,~49%,)和聚球藻(1%,~18%,)较陆架区略有增加,硅藻在陆坡的百分比不足0.5%,,可以忽略不计;海盆区,微型真核生物(28%,~92%,)所占百分比相较于陆坡依然是在春季减少,其他季节增加,总体上百分比相差不大,原绿球藻(8%,~63%,)较陆架区略有增加,聚球藻(0%,~15%,)较陆架区略有减小,硅藻在海盆区所占百分比不足0.1%,.
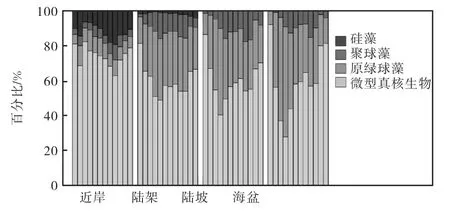
图4 按空间划分各浮游植物优势类群出现面积占各区域面积百分比Fig.4 The percentage of each dominant phytoplankton group against each area according to the spatial division
2.2 结果验证
本文研究结果与历史调查相比有较好的一致性.对南海北部网采浮游植物群落结构有很多的实测调查[8-13],结果表明硅藻为优势种,本文结果是基于色素表征的浮游植物类群,实测结果表明硅藻的丰度从沿岸到外海逐渐降低,这一点与本文反演结果相符.Ning等[14]调查表明,夏季原绿球藻的丰度最高;Liu等[15]分析了SEATS站Pico-级浮游植物,认为原绿球藻是南海丰度最大的Pico-级浮游植物,而聚球藻的丰度比原绿球藻小1个量级,这一点与本文的研究结果一致.但Liu等[15]的观测表明,原绿球藻的最大值出现在夏季,聚球藻的最大值出现在冬季到春季之间;Cai等[16]的观测表明,聚球藻和原绿球藻的丰度都是夏季高于冬季,聚球藻的丰度近岸和陆架区高于陆坡和海盆,原绿球藻分布模式与聚球藻相反,浮游植物群落变化与环境变化关系密切,实测调查结果存在年际差异;本文是多年平均的结果,与某次现场观测存在一定差异.Zhai等[17]得到了硅藻在近岸占优、而原绿球藻和微型真核生物在开阔大洋占优的结论,与本文反演结果一致.
3 营养盐对浮游植物优势类群的影响
南海浮游植物优势类群的时空分布受到“季风–环流–营养盐”变化的驱动,是物理、化学和生物耦合作用的结果[18].除近岸区外,南海呈寡营养盐状态,表层硝酸盐和磷酸盐的浓度经常在检测限附近[19-20],营养盐是限制陆架、陆坡和海盆区浮游植物生长的主要因子[21–22].
本文以1、4、7、10月为南海的冬、春、夏、秋季代表月份,图5给出了WOA13数据集气候态平均各季节南海表层营养盐浓度.按陆架、陆坡和海盆区海区平均,得到营养盐浓度在各海区的季节变化(图6),受WOA13数据集分辨率的限制,近岸区缺少营养盐数据.
在南海北部近岸区域,受沿岸上升流或入海径流(珠江等河流)的影响[23],硝酸盐和磷酸盐的浓度呈近岸高外海低,充足的氮磷营养盐适合大粒级浮游植物的生长,硅酸盐对于硅藻的生长非常重要,在近岸区域硅酸盐有较高的浓度分布[21,24],因此硅藻在南海北部沿岸能够保持占优.
陆架区,冬季,南海盛行东北季风,季风的剧烈搅动使混合层深度加深,带来了丰富的营养盐[25],表层硝酸盐浓度为0.83,µmol/L,明显高于其他季节,磷酸盐浓度为0.17,µmol/L,虽处于全年最低,但浓度并不低,在硝酸盐和磷酸盐都较为丰富的条件下,微型真核生物占绝对优势,高于其他季节,原绿球藻占一定的比重,硅藻和聚球藻的比重非常小,约占2%,.春季,硝酸盐浓度为0.17,µmol/L,为全年最低水平,磷酸盐平均浓度为0.28,µmol/L,为全年最高水平,在硝酸盐较低而磷酸盐浓度偏高的情况下,原绿球藻覆盖面积大幅增加,仅次于微型真核生物且相差不大,聚球藻也占有一定的比重,硅藻比重为全年最低,约1%,.夏季与冬季相比,磷酸盐浓度相同,硝酸盐浓度仅次于冬季,但较冬季大幅减小,为0.23,µmol/L,微型真核生物的比重减少,原绿球藻和聚球藻的比重增加,硅藻比重非常接近.秋季与春季相比,硝酸盐浓度相同,磷酸盐浓度低于春季,为0.19,µmol/L,微型真核生物和聚球藻的比重增加,原绿球藻的比重减小,硅藻比重增加,与冬、夏季基本一致.
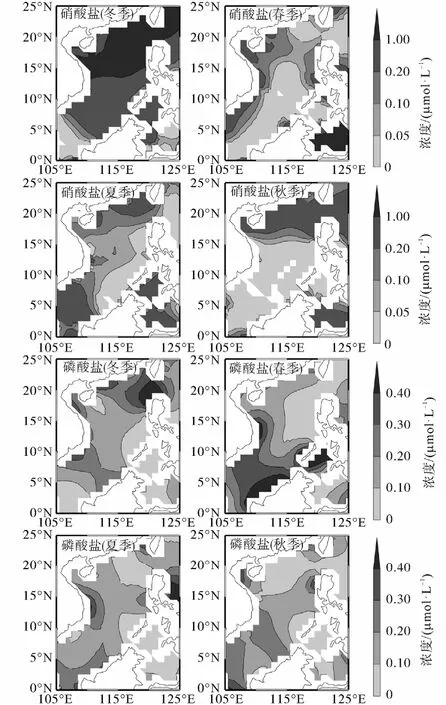
图5 南海表层气候态平均季节代表月份营养盐浓度分布图Fig.5 Climatological monthly average distribution of nutrient concentration in the surface layer of the South China Sea
陆坡区,冬季,硝酸盐和磷酸盐都比较丰富,表层硝酸盐浓度为1.39,µmol/L,全年最高,磷酸盐浓度为0.19,µmol/L,微型真核生物占绝对优势,在四季中比重最大,原绿球藻比重很小,聚球藻仅占约2%,,硅藻在陆坡区比重过小,可忽略不计.春季,硝酸盐浓度非常低,为0.09,µmol/L,磷酸盐浓度为0.21 µmol/L,在全年最高,微型真核生物、原绿球藻和聚球藻占优的面积比约为4∶5∶1.夏季,硝酸盐浓度较春季增加,为0.13,µmol/L,磷酸盐浓度较春季减小,为0.15,µmol/L,但变化幅度都较小,微型真核生物和聚球藻的比重增加,原绿球藻减小.秋季,硝酸盐浓度较春、夏季进一步增加,为0.15,µmol/L,磷酸盐浓度进一步减小,为0.10,µmol/L,聚球藻和原绿球藻的比重略有增加,微型真核生物的比重减小.
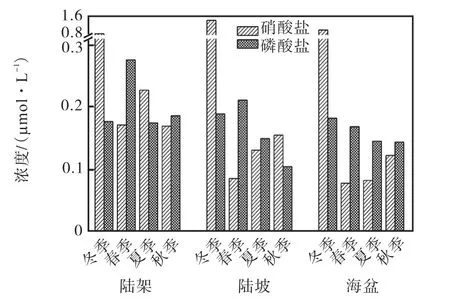
图6 气候态营养盐浓度空间平均季节变化图Fig.6 Seasonal variation of climatological spatial average nutrient concentration
海盆区,冬季,表层营养盐比较丰富,表层硝酸盐浓度为0.99,µmol/L,全年最高,磷酸盐浓度为0.18,µmol/L,微型真核生物占绝对优势,原绿球藻比重很小,几乎没有聚球藻占优分布.春季,与冬季相比,硝酸盐浓度大幅减小到全年最低水平,为0.08,µmol/L,磷酸盐浓度略有减小,为0.17,µmol/L,原绿球藻此时占绝对优势,作为优势类群出现的面积超过了海盆区面积的60%,,聚球藻比重有所增加,微型真核生物的比重减小.夏季,与春季相比,硝酸盐浓度相同,磷酸盐浓度减小,为0.15,µmol/L,但优势群落结构组成相差较大,微型真核生物比重增加,原绿球藻和聚球藻减小.秋季,磷酸盐浓度与夏季相同,硝酸盐浓度较春季增加,为0.12,µmol/L,优势类群组成比例与夏季相似,聚球藻比重增加,微型真核生物和原绿球藻的比重略有减小.
南海存在多个上升流区,在冬季吕宋上升流区和夏季越南沿岸上升流区[26],从营养盐的平面分布图中可知,磷酸盐的浓度明显高于附近海域,硝酸盐浓度也非常高,上升流带来了丰富的营养盐,微型真核生物在上升流区为优势类群.而在其他季节,上升流减弱,由于上升流的消失,导致海区的营养盐供应减少,高营养盐浓度环境会选择粒径较大的浮游植物种群[27],进而导致大粒级浮游植物受到营养盐的胁迫,最终导致浮游植物的优势类群由微型真核生物演替为微微型浮游植物聚球藻和原绿球藻.营养盐加富实验表明[21],营养盐添加能改变浮游植物的粒级结构和群落结构.
从上面的分析中可知,南海浮游植物优势类群占海区面积比例变化主要是微型真核生物和原绿球藻的季节更替.冬季各海区硝酸盐浓度都较高,磷酸盐也比较丰富,微型真核生物在高营养盐状态下占绝对优势.春季各海区的营养盐结构呈硝酸盐浓度全年最低,磷酸盐浓度全年最高,原绿球藻占绝对优势,从陆架、陆坡到海盆区,硝酸盐浓度快速降低,原绿球藻在海区占优面积的比例呈增加趋势,因为原绿球藻丰度和生长速率与磷酸盐和硝酸盐浓度都呈负相关[28],原绿球藻在低硝酸盐高磷酸盐状态下占优的几率更大.在陆坡区,从春季到秋季,硝酸盐浓度呈增加趋势,磷酸盐浓度呈减小趋势,聚球藻的比重在增加,因为聚球藻对硝酸盐反应敏感[29],相比较而言聚球藻适应高硝酸盐浓度环境.
4 结 论
本文基于水色遥感数据用PHYSAT算法反演了南海浮游植物优势类群分布的季节变化,结果与南海历史观测研究结果相对比,有较好的一致性.结果表明,微型真核生物作为优势类群在南海出现几率最大,原绿球藻次之,聚球藻再次之,硅藻最小;在季节差异上,冬季微型真核生物在南海海区所占面积的比重最高,春季原绿球藻最高,聚球藻呈双峰结构,在春季和秋季出现峰值,硅藻季节性差异较小.硅藻占优的区域主要在近岸、陆架区,外海原绿球藻和聚球藻的优势度增加,在深海盆区这两个类群与微型真核生物成为优势种的几率势均力敌.陆架、陆坡和海盆区浮游植物优势类群比例的季节变化相近,都表现为冬季微型真核生物的绝对优势和春季原绿球藻优势的大大增加,深海盆区春季原绿球藻优势甚至超过60%,,夏秋季微型真核生物在3个区域的比例恢复到50%,以上.
除河口近岸海区外,南海是典型的寡营养盐海区,硝酸盐浓度甚至比东中国海低1个量级,营养盐浓度分布的季节变化是引起浮游植物优势类群改变的主要的影响因子.硅藻在硝酸盐、磷酸盐和硅酸盐都比较充足的条件下占优;微型真核生物在硝酸盐和磷酸盐浓度都相对较高的条件下占优;原绿球藻在高磷酸盐、低硝酸盐条件下占优;聚球藻在硝酸盐浓度较高条件下优势度有所增加.随着海区中营养盐浓度的降低,浮游植物优势类群由粒径较大微型真核浮游生物向微微型浮游植物聚球藻和原绿球藻转变.
致谢:感谢NASA提供的SeaWiFS遥感数据和NOAA提供的WOA数据.
[1] Field C B,Behrenfeld M J,Randerson J T,et al.Primary production of the biosphere:Integrating terrestrial and oceanic components[J].Science,1998,281(5374):237-240.
[2] Platt T,Sathyendranath S.Oceanic primary production:Estimation by remote sensing at local and regional scales[J].Science,1988,241(4873):1613-1620.
[3] Tang D L,Kawamura H,Doan-Nhu H,et al.Remote sensing oceanography of a harmful algal bloom off the coast of southeastern Vietnam[J].Journal of Geophysical Research,2004,109:C03014-1-C03014-7.
[4] Tang D L,Kawamura H,Van Dien T,et al.Offshore phytoplankton biomass increase and its oceanographic causes in the South China Sea[J].Marine Ecology Progress Series,2004,268:31-41.
[5] International Ocean-Colour Coordinating Group.Remote Sensing of Inherent Optical Properties:Fundamentals,Tests of Algorithms,and Applications[R].Lee Z P.Dartmouth,Canada:IOCCG,2006.
[6] Alvain S,Moulin C,Dandonneau Y,et al.Seasonal distribution and succession of dominant phytoplankton groups in the global ocean:A satellite view[J].Global Biogeochemical Cycles,2008,22(3):GB3001-1-GB3001-15.
[7] Alvain S,Moulin C,Dandonneau Y,et al.Remote sensing of phytoplankton groups in case 1 waters from global SeaWiFS imagery[J].Deep Sea Research Part I:Oceanographic Research Papers,2005,52(11):1989-2004.
[8] 孙军,宋书群,乐凤凤,等.2004年冬季南海北部浮游植物[J].海洋学报,2007,29(5):132-145.
[9] 乐凤凤,孙军,宁修仁,等.2004年夏季中国南海北部的浮游植物[J].海洋与湖沼,2006,37(3):238-248.
[10] 马威,孙军,田伟.2009年冬季南海北部网采浮游植物群落结构[J].海洋科学,2011,35(9):8-13.
[11] 宫相忠,马威,田伟,等.2009年夏季南海北部的网采浮游植物群落[J].中国海洋大学学报:自然科学版,2012,42(4):48-54.
[12] 柯志新,黄良民,谭烨辉,等.2007年夏季南海北部浮游植物的物种组成及丰度分布[J].热带海洋学报,2011,30(1):131-143.
[13] 朱根海,宁修仁,蔡昱明,等.南海浮游植物种类组成和丰度分布的研究[J].海洋学报:中文版,2003(S2):8-23.
[14] Ning X,Chai F,Xue H,et al.Physical-biological oceanographic coupling influencing phytoplankton and primary production in the South China Sea[J].Journal of Geophysical Research,2004,109:C10005-1-C10005-20.
[15] Liu H,Chang J,Tseng C M,et al.Seasonal variability of picoplankton in the Northern South China Sea at the SEATS station[J].Deep Sea Research Part II:Topical Studies in Oceanography,2007,54(14/15):1602-1616.
[16] Cai Y M,Ning X R,Liu C G,et al.Distribution pattern of photosynthetic picoplankton and heterotrophic bacteria in the northern South China Sea[J].Journal of Integrative Plant Biology,2007,49(3):282-298.
[17] Zhai H,Ning X,Tang X,et al.Phytoplankton pigment patterns and community composition in the northern South China Sea during winter[J].Chinese Journal of Oceanology and Limnology,2011,29(2):233-245.
[18] Ning X,Cai Y,Li G,et al.Photosynthetic picoplaakton in the northern South China Sea[J].Acta Oceanologica Sinica,2002,25(3):82-97.
[19] 蔡昱明,宁修仁,刘诚刚.1999年夏季南海北部和北部湾海域粒度分级叶绿素a和初级生产力的分布特征[J].海洋科学集刊,2002(44):11-21.
[20] Chen Y L,Chen H Y,Karl D M,et al.Nitrogen modulates phytoplankton growth in spring in the South China Sea[J].Continental Shelf Research,2004,24(4):527-541.
[21] 彭欣,宁修仁,孙军,等.南海北部浮游植物生长对营养盐的响应[J].生态学报,2006,26(12):3959-3968.
[22] 郝锵,宁修仁,刘诚刚,等.南海北部初级生产力遥感反演及其环境调控机制[J].海洋学报,2007,29(3):58-68.
[23] 薛惠洁,柴扉,徐丹亚,等.南海海流数值计算[J].中国海洋学文集:南海海流数值计算及中尺度特征研究,2001(13):1-14.
[24] 黄邦钦,洪华生,柯林,等.珠江口分粒级叶绿素a和初级生产力研究[J].海洋学报,2006,27(6):180-186.
[25] 赵辉,齐义泉,王东晓,等.南海叶绿素浓度季节变化及空间分布特征研究[J].海洋学报,2005,27(4):45-52.
[26] 柴扉,薛惠洁,侍茂崇.南海升降流区域分布及形成机制分析[J].中国海洋学文集:南海海流数值计算及中尺度特征研究,2001(13):117-128.
[27] Vaillancourt R D,Marra J,Seki M P,et al.Impact of a cyclonic eddy on phytoplankton community structure and photosynthetic competency in the subtropical North Pacific Ocean[J].Deep Sea Research Part I:Oceanographic Research Papers,2003,50(7):829-847.
[28] Partensky F,Blanchot J,Vaulot D.Differential distribution and ecology of Prochlorococcus and Synechococcus in oceanic waters:A review[J].Bulletin de l'institut d'océanographie,1999,19:457-476.
[29] Glover H E,Prézelin B B,Campbell L,et al.A nitratedependent Synechococcus bloom in surface Sargasso Sea water[J].Nature,1988,331(6152):161-163.
责任编辑:郎婧
Seasonal Distribution of Dominant Phytoplankton Groups in the South China Sea Based on PHYSAT
LI Yueyang1,SUN Qun1,WANG Lei1,WEI Hao2
(1.College of Marine Science and Engineering,Tianjin University of Science & Technology,Tianjin 300457,China;2.School of Marine Science and Technology,Tianjin University,Tianjin 300072,China)
PHYSAT algorithm was used to reproduce the average seasonal distribution of the dominant phytoplankton groups in the South China Sea based on water color satellite data.The results show that nanoeukaryote is the most dominant group in the South China Sea in the whole year,with Prochlorococcus,Synechococcus and diatoms following in turn.Diatoms is mostly observed in the coastal and continental shelf areas,while Prochlorococcus and Synechococcus dominance increased in the offshore.Nanoeukaryotes dominance is very high in winter but Prochlorococcus in spring.Synechococcus seasonal distribution is bimodal with peaks occurring in spring and autumn.The seasonal variation of diatoms dominance is very small.Seasonal variation of the proportion of phytoplankton dominant groups in continental shelf,continental slope and basin is very similar.Nanoeukaryotes are predominating in winter.The dominance of Prochlorococcus greatly increases in spring compared with other seasons.Seasonal variation of nutrient concentration is the main factor influencing the distribution of the dominant phytoplankton groups.With the nutrient concentration decreasing,phytoplankton dominant group changes from nanoplanktonic nanoeukaryotes species to picoplanktonic Prochlorococcus and Synechococcus species.
dominant phytoplankton groups;PHYSAT algorithm;water color satellite data;WOA nutrient concentration;South China Sea
P735
A
1672-6510(2015)05-0042-06
10.13364/j.issn.1672-6510.20140165
2014-12-26;
2015-01-29
国家重点基础研究发展计划(973计划)资助项目(2011CB403606,2010CB428904);国家自然科学基金资助项目(41376112,41276016,41406158)
李月洋(1991—),女(蒙古),内蒙古通辽人,硕士研究生;通信作者:孙 群,副教授,sunqun@tust.edu.cn.
数字出版日期:2015-05-08;数字出版网址:http://www.cnki.net/kcms/detail/12.1355.N.20150508.1519.006.html.
